
恩施盛家坝红椿天然种群动态与预测
2016-09-15 15:52闵水发江雄波操英南付翠林
福建林业科技 2016年2期
汪 洋,闵水发,江雄波,操英南,张 敏,付翠林,白 琳
(湖北生态工程职业技术学院,湖北 武汉 430200)
恩施盛家坝红椿天然种群动态与预测
汪 洋,闵水发,江雄波,操英南,张 敏,付翠林,白 琳
(湖北生态工程职业技术学院,湖北 武汉 430200)
通过对典型代表意义的红椿样地调查和数据统计分析,绘制红椿自然种群的龄级结构图,量化分析种群各龄级间个体数量、变化动态。编制种群的特定时间生命表,分析存活曲线、死亡率曲线和寿命期望等重要参数,并运用时间序列模型预测红椿种群数量动态。结果表明:①红椿种群中,1龄级和7龄级存活数分别占总数的28.57%和16.07%。27.5 cm以上径级的立木有10.71%。动态指数Vpi=27.84%>0。②生命表表明,种群死亡趋势前期高于后期,死亡高峰在1龄级和4龄级。存活曲线不符合Deevey型,说明环境筛和种群竞争加强造成波动。③时间动态对第3、5、7龄级预测表明,种群呈现老龄个体增加的趋势。④红椿种群死亡率曲线与消失率曲线基本一致,即在1~7龄级波动很大,其中3~4龄级时死亡率和损失度最大。研究认为:红椿种群的生长发育主要受限于其自身的生物学特性、种内竞争和环境干扰。对现存种群和生境的保护,以及适当的人工辅助措施是红椿种群恢复的关键。
红椿;生命表;种群动态;种群结构;预测
研究种群动态变化及其群落的演替趋势,对阐明生态系统发展、群落结构特征和发展趋势有重要的研究意义[1-2]。种群统计的核心是建立反映种群全部生活史的各年龄组或生活级出生率、死亡率,甚至包括迁移率在内的信息综合表,即生命表[3-4]。静态生命表、种群结构动态量化分析法和时间序列预测法是研究种群数量动态变化和进行种群统计的重要工具[5]。
红椿(ToonaciliataRoem.)又名红楝子,为楝科(Meliaceae)香椿属(Toona)落叶或近常绿高大乔木。是国家Ⅱ级植物,有“中国桃花心木”之称[6-7]。作为南方地区珍贵的乡土用材树种,因其速生性好、适应性强,是很好的造林树种,在山区造林有很大潜力。红椿分布在东经100°16′—119°40′、北纬24°21′—32°42′[8-9]。在我国分布不广,且呈天然零星分布[10]。过度开发以及天然更新较慢,其数量不断减少[3]。因而,对红椿进行全面系统的种群动态分析,发现其濒危机制并采取科学有效的保护措施十分重要。
1 研究地概况
研究样地位于湖北恩施市盛家坝的马鹿河流域,在星斗山国家级自然保护区东部边界,东经109°13′7.03″、北纬30°00′14.43″,属于中亚热带与北亚热带的过渡地带,亚热带大陆性季风气候。该区年均气温14.9 ℃,年均降水量1400 mm以上,土壤为泥质页岩发育成的以扁砂土为主的质地疏松的山地黄壤[11-12],pH 6.5,土壤自然含水率较高,有机质、全氮、有效氮、全磷、速效钾含量相对较高。植被生长繁茂,红椿为群落优势种。
该红椿群落为天然林,乔灌草丰富度较高。虽受一定人为干扰,但群落恢复较好。乔木主要有红椿、绒毛钓樟(Linderafloribunda)、黑壳楠(Linderamegaphylla)、竹叶楠(Arundinagraminifolia)、鳞木稠李(Padusbuergeriana)、红果黄肉楠(Actinodaphnecupularis)、紫弹树(Celtisbiondii)、小叶青冈(Cyclobalanopsismyrsinifolia)等。灌木层主要有臭牡丹(Clerodendrumbungei)、棠叶悬钩子(RubusmalifoliusFocke)、砚壳花椒(Zanthoxylumdissitum)、中国旌节花(StachyuruschinensisFranch)、枇杷叶荚蒾(Viburnumrhytidophyllum)等27种。草本主要有楼梯草(Elatostemainvolucratum)、透茎冷水花(Pileapumila)、团叶鳞始蕨(Lindsaeaorbiculata)、野棉花(Anemonevitifolia)、赤爮(Thladianthadubia)、青牛胆(金果榄)(Tinosporasagittata)、打破碗花花(Anemonehupehensis)等38种。群落内部藤本植物丰富,群落结构完整。
2 研究方法
2.1 野外调查
红椿在沿河、山沟等水湿环境较好的地段多有分布,但以红椿为优势种的天然群落比较少见。天然群落一般种群规模较小,地势环境复杂。经过2 a的实地调查发现,大于400 m2的天然群落极少。根据现存红椿种群很小的特点和盛家坝红椿群落的现有规模,设置样地完全覆盖所有红椿种群,即:20 m×20 m。样地内设16个5 m×5 m的样方。记录样方内生境、群落学特征。每木调查并记录样方中物种名、树高、胸径、冠幅及生长情况等(表1)。每木调查时,以基株为单位进行统计,如果1个树桩上有多个无性系分株,则记录径级最大的1株。记录样方内胸径≥2.5 cm植株的胸径、基径、树高、冠幅、枝下高;胸径<2.5 cm的植株,记录株数和高度。该种群共有红椿大小植株56株,种群密度为1400 株·hm-2。该种群作为湖北恩施地区较为典型且相对完整的种群,对其种群生态学研究具有代表性。

表1 盛家坝红椿群落样地的环境特征
2.2 龄级划分及特征分析
红椿为国家二级重点保护植物,不能测定每个个体的确切年龄。根据戴其生等[13]对红椿用材林解析数据,以及龙汉利等[9]提出的红椿胸径平均、连年生长过程表,采用“空间替代时间法”,即将林木依胸径大小分级,以立木级结构代替种群年龄结构分析种群动态[14-18]。结合样地内最大立木径级为36 cm,将该种群划分为8个龄级。幼苗级即1龄级:(DBH≤2.5 cm);从第2龄级开始,以胸径5 cm为步长增加1级,幼树级即2龄级:(2.5 cm≤DBH<7.5 cm)、3龄级:(7.5 cm≤DBH<12.5 cm)、4龄级:(12.5 cm≤DBH<17.5 cm),中树级即5龄级:(17.5 cm≤DBH<22.5 cm)、6龄级:(22.5 cm≤DBH<27.5 cm),大树级即7龄级(27.5 cm≤DBH<32.5 cm)、8龄级:(DBH≥32.5 cm)。将树木径级从小到大的顺序看作是时间顺序关系。统计各龄级的植株数,编制红椿种群静态生命表,分析其动态变化规律。
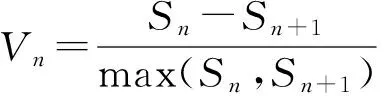
2.3 生命表编制

2.4 种群数量动态时间序列预测
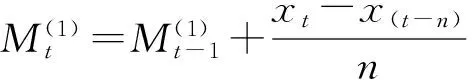
2.5 数据处理与分析
文中所有数据通过IBM的SPSS 22和Excel 2007软件处理。
3 结果与分析
3.1 红椿种群龄级结构
以8个年龄级为横坐标,以各龄级个体数为纵坐标,绘制红椿种群龄级结构分布图。由图1可以看出,400 m2样地,种群径级结构不规则,8个立木级均有分布,以1龄级、7龄级较多,分别占总数的28.57%、16.07%。最大径级(27.5 cm)以上的立木占10.71%;2~7龄级出现较大波动,2,4,6,7等龄级分别低于后一龄级;而幼苗数量占比最大,这充分说明环境筛对红椿幼苗和幼树的作用。红椿属于强阳性树种,对林窗的依赖较强,因此,较高的郁闭度限制幼苗向幼树发展。另外,红椿个体竞争在4~7龄级加强,死亡率较高,该现象表明红椿幼树个体向大树阶段的发育是不连续的。
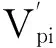

表2 红椿种群径级结构动态变化指数
3.2 生命表与存活曲线
静态生命表是根据一定时期样地内一个种群所有个体的年龄数据编制而成的[14,25-26],通过静态生命表可分析群体的出生率、死亡率等重要参数,提供更多关于种群年龄结构和数量统计方面的信息[27]。由表3可知,随着年龄的增加,死亡数dx呈现剧烈下降,再缓慢增加的趋势,1龄级~2龄级死亡数最大。平均存活数Lx随年龄增加出现2次波动,1龄级最大,6龄级达到最大值。说明达到一定龄级后,红椿种群趋于稳态。总存活数Tx线性递减。生命期望值Ex表现的是个体的平均生存能力大小[27],8龄级最小,4龄级出现峰值。损失度Kx与生命期望值Ex均在第2龄级和第4龄级大幅上升,说明红椿种群衰减的生理与环境筛选相关。
以种群的龄级作为横坐标,以生命表中存活量lx的自然对数lnlx为纵坐标,绘制得到红椿种群的存活曲线(图2)。可以发现,1~2龄级,3~4龄级死亡率出现2次突变,可以分别预测环境筛的压力、种类种间的生存竞争加剧以及来自种群外的干扰。
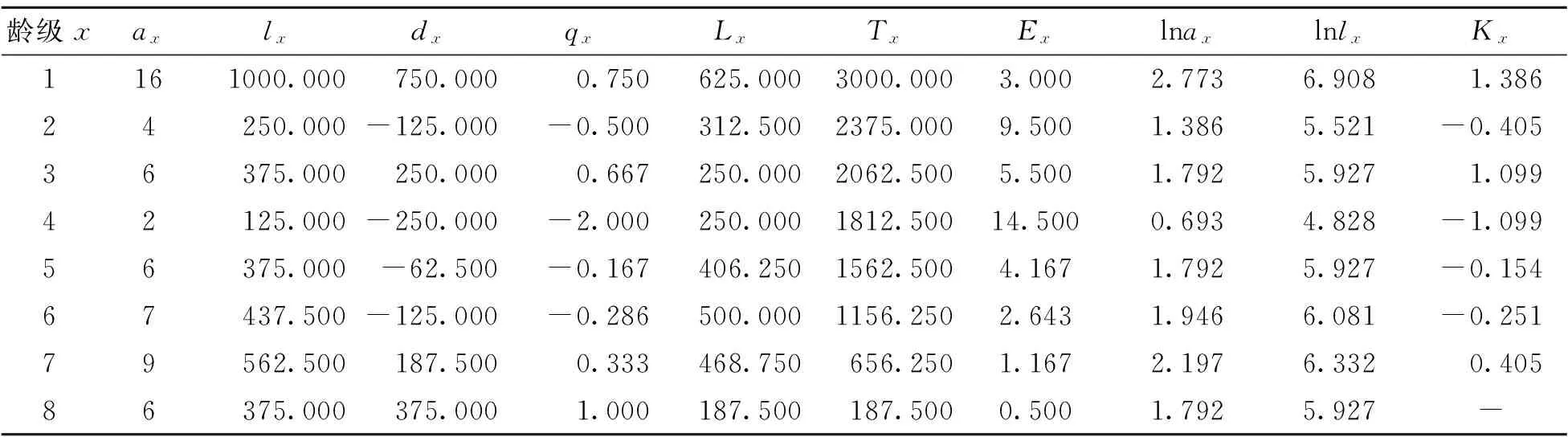
表3 红椿种群静态生命表
鉴于图2曲线的波动特征,为了检验种群存活曲线是否符合Deevey 3种类型存活曲线[4,27-29],分别用指数函数方程式Nx=N0e-bx和幂函数方程式Nx=N0x-b进行回归,得到如下方程:N=5.937e-0.00x,R2=0.000;N=6.175x-0.03,R2=0.053。
Deevey的3种类型曲线均不能解释盛家坝红椿种群存活类型。通过SPSS软件用不同的回归方程对图2的数据进行存活曲线拟合,方程为:y=-0.038x3+0.592x2-2.641x+8.972,R2=0.689,F=2.956。方程比较近似地反应了该种群的存活曲线特征。通过方程可以预计各龄级个体数量。
3.3 种群数量动态序列预测
采用一次移动平均法对红椿的种群存活曲线结构进行模拟和预测[15,24,30]。其结果(图3)可以看出,预测到第3龄级时,除第4龄级植株数量下降外,种群各径级存活数峰值在预测序列中依次向后推移。依据现有种群存活值,在3龄级以后,幼树的比例下降较快,成树存活数量少量下降,老树比例上升。预测到5龄级时,种群存活数在6龄级数量微降,7、8龄级种群保持稳态,成树比例较大。预测到第7龄级时,成树比例占最大,种群呈老龄型稳态。可以推断,如不采取适当的保护措施,为幼苗更新创造条件,红椿种群将趋向老龄化。
3.4 死亡率和损失度曲线
种群死亡率曲线反映种群死亡的动态变化过程[31]。分别以死亡率、损失度为纵坐标,以龄级为横坐标,绘制红椿种群的死亡率曲线(qx)、消失率(损失度)曲线(Kx)。从图4可知,红椿种群死亡率曲线与消失率曲线基本一致,即在1~7龄级波动很大,其中3~4龄级时死亡率和损失度最大,表明此时环境筛选强度很高,经过第4龄级后的幼树,逐步进入稳定期。7龄级时呈现一个损失度峰值。红椿幼苗期和进入成树期2个阶段,首先受到环境筛的强烈选择而淘汰大部分幼苗,其后的高损失度来自种间竞争加剧和非自然干扰。
4 讨论与结论
红椿种群的龄级结构、生命表、存活曲线和死亡率曲线是红椿种群生物学特性与环境因素共同作用的结果[25]。作为种群动态的重要特征之一,存活曲线反映了生存率和死亡率随年龄的变化状况[32]。红椿龄级存活数量的分布分析表明,存活植株与龄级之间变化关系存在着一种显著的波动规律,这种波动是种群与环境相互选择的过程。
很多学者在生命表中,对不同龄级数量进行匀滑修正,以避免死亡率为负值[14]。由于本研究数据源自天然种群,并且静态生命表是以时间推空间,以横向导纵向,故在生命表的编制中,会出现死亡率为负的现象,这虽与生命表的数学假设相违背,但仍能作为重要的生态学依据反映种群的发展和衰落[15,22]。表3中,除第1龄级的标准化存活量(lx)为最高值(1000)外,种群第2、4、5龄级出现了死亡率为负的现象,说明2、4、5龄级幼树和小树数量不足。因此,必须补充相应的负值数量,才能维持红椿种群平衡,这表征了种群受环境筛选和红椿生理特点制约。
从群落特征分析,红椿所在群落属于亚热带大陆性季风气候常绿落叶阔叶林。群落上层优势种以红椿为主。群落林分湿度大,层次明显,乔木层树冠较连续,郁闭度高(0.85)。乔木层主要树种为绒毛钓樟、黑壳楠、竹叶楠、鳞木稠李、红果黄肉楠、紫弹树、小叶青冈等,均是阔叶乔木,树冠较大,对空间、环境资源都具有一定的竞争要求。大型藤本较多,如乌泡子数量极其丰富,整个样地统计为78株。其他如小叶爬崖香(Piperarboricola)、棠叶悬钩子、砚壳花椒、绣球藤(Clematismontana)、石岩枫(MallotusrepandusMuell)等,已进入乔木层上层。环境条件决定了红椿个体间及与其他种群间对光照、水分、养分和空间生态位竞争,种群自疏作用增强,环境筛的选择强度增大,对红椿幼树及成年植株造成一定程度的损失。红椿种群中大树的保存,主要来自于“林窗机遇”或人为干扰因素。
对红椿及其生存环境的人为干扰主要有砍伐和开垦等。对个体尤其是幼树的砍伐,常使其达不到成熟年龄就死亡,导致红椿径级结构失调或变化剧烈,种群不能正常发展。通过对湖北全省红椿资源调查发现,红椿种群全部在相应的群落边缘。红椿幼苗在每个样方中均存在,但第2、4龄级的植株大多存在于群落边缘,能获得较强的光照,得以存活。因此,对现存种群及生境保护和适当的人工辅助措施是红椿种群恢复的关键。
[1]张钦弟,张金屯,苏日古嘎,等.宠泉沟向然保护区华北落叶松种群生命表与谱分析[J].应用与环境生物学报,2010,16(1):1-6.
[2]殷东生,张凤海,山永清,等.色木槭次生林种群结构动态分析[J].林业科技,2011,11(6):19-22.
[3]谢宗强,陈伟烈,路鹏,等.濒危植物银杉的种群统计与年龄结构[J].生态学报,1999,19(4):523-528.
[4]胡喜生,洪伟,吴承祯,等.木荷天然种群生命表分析[J].广西植物,2007,27(3):469-474.
[5]李妍,李登武,韩东辰.贺兰山东坡青海云杉种群动态[J].浙江农林大学学报,2014,31(1):50-56.
[6]中国树木志编委会.中国主要树种造林技术[M].北京:中国林业出版社,1981.
[7]郑万钧.中国树木志:第1卷[M].北京:中国林业出版社,1983:797.
[8]吴际友,程勇,王旭军,等.红椿无性系嫩枝扦插繁殖试验[J].安徽林业科技,2011,38(4):5-8.
[9]龙汉利,冯毅,向青,等.四川盆周山地红椿生长特性研究[J].四川林业科技,2011,32(3):37-41.
[10]邹高顺.珍贵树种红椿与毛红椿引种栽培研究[J].福建林学院学报,1994,14(3):271-276.
[11]陈绍林,张志华,廖于实,等.星斗山自然保护区秃杉原生种群生境现状及保护对策[J].安徽农业科学,2008,36(11):4624-4625.
[12]刘毅,孙云逸,陈世明,等.湖北星斗山国家级自然保护区生物多样性分析与保护[J].湖北林业科技,2007(6):42-45.
[13]戴其生,张梅林,徐玉伟,等.红楝子人工造林试验初报[J].安徽林业科技,1997(1):33-34.
[14]江洪.云杉种群生态学[M].北京:中国林业出版社,1992:8-26.
[15]吕海英,王孝安,李进,等.珍稀植物银砂槐中国分布区的种群结构与动态分析[J].西北植物学报,2014,34(1):177-183.
[16]康华靖,陈子林,刘鹏,等.大盘山自然保护区香果树种群结构与分布格局[J].生态学报,2007,27(1):389-396.
[17]杨宁,邹冬生,杨满元,等.贵州雷公山秃杉的种群结构和空间分布格局[J].西北植物学报,2011,31(10):2100-2105.
[18]张兴旺,张小平,郝朝运,等.安徽琅琊山青檀种群结构与空间分布格局研究[J].广西植物,2011,31(6):782-788.
[19]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
[20]LEAK W B.Age distribution in Virgin red spruce and Northern Hard woods[J].Ecology,1975(56):1451-1454.
[21]洪伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.
[22]吴承祯,洪伟,谢金寿.珍稀濒危植物长苞铁杉种群生命表分析[J].应用生态学报,2000,11(3):333-336.
[23]张文辉,王延平,康永祥,等.濒危植物太白红杉种群年龄结构及其时间序列预测分析[J].生物多样性,2004,12(3):361-369.
[24]卢杰,李江荣,郭其强,等.藏东南林芝云杉种群结构与数量动态[J].林业资源管理,2014(4):68-73.
[25]程红梅.大蜀山短毛椴种群生命表与生存分析[J].浙江大学学报:农业与生命科学版,2010,36(3):341-347.
[26]朱慧,洪伟,吴承祯,等.天然更新的檫木林根系生物量的研究[J].植物资源与环境学报,2003,12(3):31-35.
[27]万志兵,程国源,丁强强,等.绩溪光皮桦次生林的种群动态[J].东北林业大学学报,2014,42(1):27-30.
[28]杜兴乔,刘映良.荔波瘤果茶种群生命表分析[J].安徽农业科学,2009,37(33):16613-16615.
[29]郭华,王孝安.子午岭马栏林区油松种群结构分析[J].陕西师范大学学报:自然科学版,2012,40(1):67-71.
[30]薛瑶芹,张文辉,何景峰.黄龙山白桦种群结构及其时间序列预测分析[J].西北农林科技大学学报:自然科学版,2008,36(12):121-127.
[31]孟宪帅,韦小丽.濒危植物花榈木野生种群生命表及生存分析[J].种子,2011,30(7):66-68.
[32]谢伟东,陈建华,赖家业,等.珍稀濒危植物蒜头果种群生命表分析[J].中南林业科技大学学报,2009,29(2):73-76.
Dynamics and Estimation ofToonaciliataRoem.Population in Shengjiaba,Enshi City
WANG Yang,MIN Shui-fa,JIANG Xiong-bo,CAO Ying-nan,ZHANG Min,FU Cui-lin,BAI Lin
(Hubeivocationalcollegeofecologicalengineering,Wuhan430200,Hubei,China)
By a field survey and statistic data analyses of a typical sampling plot ofToonaciliatain Shengjiaba of Enshi City,the age-class survival curve figure ofT.ciliata′snatural population was drawn;the quantitative analysis of the individual number of different age classes and its dynamic condition were conducted;the life table in specific time of the population was made,and the survival curve,the mortality rate curve and the life-expectation table were analyzed.The result indicated:First,the survival number of age-class 1 and age class 7 covered 28.57% and 16.07% respectively,and the largest age-class (above 27 cm in diameter) covered merely 10.71% of the total.The dynamic change index of the age structure of the populationVpi=27.84%>0,indicating the population was in growth form.Second,the analysis of static life table demonstrated that the former tendency of population mortality was stronger than that of the latter,and that the mortality peak fell between age-class 1 and 4.The survival curve of the population was not in accordance with any Deevey curves,indicating the intensification of environment sieve and competition between populations.Third,time-dynamic expectation of the population to 3rd,5th and 7th age classes showed the surging trend of aging individuals in the population.Forth,the mortality rate curve and the disappearance index curve ofT.ciliatapopulation were basically the same,and more specifically,the curves fluctuated severely,with the mortality rate and the disappearance index of 3rd and 4th age-classes at maximum.The research showed that the growth and development ofT.ciliatapopulation conformed to its biological characteristics,intraspecific competition and environmental disturbance,and therefore,the key to the restoration ofT.ciliatapopulation lies in the protection of current populations and its habitats,as well as appropriate human assistances.
Toonaciliata;static life table;population dynamics;population structure;estimation
10.13428/j.cnki.fjlk.2016.02.004
2015-07-05;
2015-08-01
湖北省科技厅公益性研究项目(40 2012DBA40001)
汪洋(1968—),男,湖北武汉人,湖北生态工程职业技术学院副教授,从事植物生态学和遗传教学与科研。E-mail:13296698026@163.com。
江雄波(1975—),男,湖北武汉人,湖北生态工程职业技术学院高级工程师,从事林业及栽培研究。E-mail:stxycyb@163.com。
S718.54
A
1002-7351(2016)02-0021-06
猜你喜欢
中老年保健(2022年5期)2022-11-25
今日农业(2022年15期)2022-09-20
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
中老年保健(2021年4期)2021-08-22
今日农业(2021年5期)2021-05-22
科学之谜(2020年6期)2020-08-11
生物学教学(2018年3期)2018-08-08
中学生物学(2018年8期)2018-03-01
食品工业科技(2014年13期)2014-03-11
