
紫外线胁迫对红色型豌豆蚜生物学特性的影响
2016-09-14 01:19:40袁伟宁朱仰艳孙小玲杨巧燕刘长仲
植物保护 2016年4期
袁伟宁, 朱仰艳, 孙小玲, 杨巧燕, 刘长仲
(甘肃农业大学草业学院, 草业生态系统教育部重点实验室, 兰州 730070)
紫外线胁迫对红色型豌豆蚜生物学特性的影响
袁伟宁,朱仰艳,孙小玲,杨巧燕,刘长仲*
(甘肃农业大学草业学院, 草业生态系统教育部重点实验室, 兰州730070)
由于臭氧层消耗,南北半球中高纬度地区紫外线(UV-B)胁迫增强,为了探求增强的UV-B胁迫对红色型豌豆蚜生物学特性的影响,以期明确UV-B辐射在豌豆蚜种群演替和种下分化中的作用。本研究以40 W紫外线灯连续辐射处理红色型豌豆蚜8代,每代从初产若蚜开始每天处理1次,连续照射6 d,分20、30、40、50和60 min 5个处理,并设置白炽灯对照。结果显示,同一世代,随着UV-B辐射时间延长,豌豆蚜体长逐渐减短,体重逐渐减轻,净增殖率、内禀增长率、周限增长率逐渐减小,平均世代周期逐渐延长。同一UV-B辐射时间下,随着世代增加,豌豆蚜体长、体重、净增殖率、内禀增长率、周限增长率均呈先上升后显著下降的趋势,平均世代周期呈先下降后显著上升的趋势。至F8代,不同辐射时间下豌豆蚜平均体长为对照的42.58%~81.94%,平均体重为对照的33.77%~73.12%,净增殖率为对照的6.94%~46.55%,周限增长率为对照的77.00%~87.94%,平均世代周期为对照的1.20~1.38倍,内禀增长率降低,且50~60 min处理下小于对照的1/2,种群数量增长放缓。以上说明长期UV-B胁迫对豌豆蚜的生长发育和种群增长最终都有抑制作用。
UV-B;豌豆蚜;生命表;种群参数;内禀增长率
生物体受紫外线损害的程度与紫外线的辐射波长相关,紫外线波段一般分为3部分:UV-A(320~400 nm),UV-B(280~320 nm),UV-C(200~280 nm)[1]。根据1979-1993年卫星测得的全球臭氧资料,由于对流层的污染,导致平流层臭氧量减少,致使南北半球中高纬度地区UV-B辐射显著增加[2],并对水陆生态系统的生物和化学链产生了广泛影响[3]。UV-B选择压力的增强,导致蚜虫(Aphidoidea)种下遗传分化普遍[4]。有报道称,紫外胁迫(UV-B)使蚜虫种群相关防御基因表达增加[5-6],上调表达生长因子[7],类似于蚜虫对光周期[8]、低温胁迫[9]、重金属胁迫[10]等的抗逆反应。蚜虫还有可能在UV胁迫下,通过体内共生菌影响热休克蛋白的表达[11]。豌豆蚜(AcyrthosiphonpisumHarris)是重要的牧草害虫,多聚集在寄主植物的幼嫩部位取食汁液,影响牧草生长发育、开花结实和产量[12],并且其分泌的蜜露可以引起叶片发霉, 影响牧草的质量,最为严重的是它是苜蓿花叶病毒的传播媒介,其造成的损失远远超过蚜虫的直接为害[13],因此豌豆蚜成为农牧业主要的防治对象之一。国际蚜虫基因组联盟(IAGC)已于 2010 年完成了豌豆蚜全基因组序列的测定工作, 并在《Insect molecular biology》发表了专辑[14]。在我国,绿色型豌豆蚜各地均有分布,而红色型分布范围较窄[15],2009年调查,红色型豌豆蚜主要分布在甘肃、青海、宁夏、新疆等地[16]。2013年,金娟等[17]对甘肃省内4个红色型豌豆蚜的地理种群遗传多样性进行了报道。杜军利等[18]认为,近几年我国豌豆蚜种群发生了很大变化,由于包括UV辐射等因素的影响,豌豆蚜已成为苜蓿(Medicagosativa)田的优势害虫,红色型所占的比例也在逐年上升。
虽然领域内学者开展了较多关于紫外辐射对蚜虫影响的研究,但多以高强度UV-B辐射做一次性处理[19-24]或只处理一代内的蚜虫[25],而自然环境中UV-B辐射强度较低,并一直对生物产生胁迫,前人处理条件与自然环境差异很大。本研究以豌豆蚜为材料,小剂量UV-B连续胁迫多代,以观察UV-B胁迫对豌豆蚜生物学特性的影响,从而探求UV-B辐射在豌豆蚜种群动态中的作用,为豌豆蚜生态遗传与进化及害虫综合治理提供理论依据。
1 材料与方法
1.1供试材料
红色型豌豆蚜于2015年4月采自甘肃农业大学苜蓿(M.sativa)试验基地。将无翅胎生雌蚜单头饲养在盆栽‘临蚕9号’蚕豆(ViciafabaLinn.)植株上进行单克隆系的培养,环境条件:温度(25±1)℃,湿度50%±10%,光照L∥D=16 h∥8 h。
1.2试验方法
1.2.1试虫饲养
采用“离体叶片”饲养法。在培养皿底部垫一张定性滤纸,滤纸中央铺一块与蚕豆叶片大小相仿的脱脂棉,厚度不超过2 mm,脱脂棉上平铺一片蚕豆叶片,背面朝上,叶柄用脱脂棉缠裹,在处理好的培养皿中注入蒸馏水,以滤纸和脱脂棉含满水为宜。单头接入若蚜后在温度为(25±1)℃,相对湿度50%±10%,光周期L∥D=16 h∥8 h的RH-250-G型光照培养箱中进行饲养。
1.2.2UV-B辐射若蚜
每世代将12 h内产的若蚜饲喂12 h后置于40 W紫外灯(波长313 nm)下垂直距离30 cm处,分别处理20、30、40、50、60 min,对照组(CK)白炽灯照射。每个处理60头,每天1次,连续处理6 d,连续试验8代。
1.2.3观察记载方法
对处理的蚜虫每隔12 h观察1次,记录每头蚜虫的蜕皮和死亡时间,产蚜量,直至成虫死亡。若蚜发育至4龄时测量其体重、体长、体宽。
1.3统计方法
种群参数:净增殖率R0=∑lxmx; 平均世代周期T=∑xlxmx/∑lxmx;内禀增长率rm=lnR0/T;周限增长率λ=exp(rm);式中,x为时间间隔(d),lx为在x期开始时的存活率,mx为在x期间平均单头产蚜量。
试验数据采用 SPSS 19.0 软件进行方差分析和 Duncan 多重比较,Excel 2007制表。
2 结果与分析
2.1连续UV-B胁迫对红色型豌豆蚜体长的影响
UV-B胁迫影响豌豆蚜体型的大小。相同世代中,红色型豌豆蚜在UV-B胁迫下,体长随着辐射时间延长而逐渐减短(表1)。20~30 min胁迫下F5~F8代显著低于对照(P<0.05),40~60 min胁迫下各代均显著低于对照(P<0.05)。在相同UV-B辐射时间处理下,随着豌豆蚜受辐射世代数的增加,体长均表现出先增长后减短的趋势。在UV-B 20~50 min处理下,F4代相对于F1代体长增长,但均未达到显著水平(P>0.05)。F5~F8代体长较F4代均明显减短(P<0.05),且随着代数的增加显著缩短(P<0.05),20、40、50、60 min 处理下F8代体长最短,30 min处理下虽然F7代体长最短,但F7代与F8代之间没有显著差异。20~50 min处理下,F8随着辐射时间的延长依次为对照的81.94%、79.03%、69.68%、61.61%。60 min处理下F4代开始逐渐减短,至F8代最小,为对照的42.58%。

表1 不同UV-B辐射时间连续胁迫红色型豌豆蚜8代的体长变化1)
1) 同列中不同大写字母表示同一辐射时间下不同世代之间差异显著(P<0.05),同行中不同小写字母表示相同世代不同辐射时间之间差异显著(P<0.05)。下同。
The different uppercase letters in the same column and the lowercase letters in the same row indicate significantly different at 5% level. The same below.
2.2连续UV-B胁迫对红色型豌豆蚜体重的影响
由表2可以看出,长时间UV-B胁迫对豌豆蚜的体重产生了较大影响。同世代豌豆蚜的体重随着UV-B胁迫时间的延长而逐渐减轻,F1~F4代40~60 min处理下显著减轻,F5~F8代各处理下均显著减轻,各代豌豆蚜的体重均以60 min处理下最轻。
同一胁迫时间下随着辐射处理代数的增加体重先增加后减少。20~50 min处理时间下体重均增加至F4代后逐渐减轻,F8代时显著减轻,随着辐射时间的延长依次为对照的73.12%、58.03%、44.59%、38.03%。60 min处理下豌豆蚜体重增大至F3代后逐渐减轻,F8代豌豆蚜体重最轻,为对照的33.77%。
表2不同UV-B辐射时间连续胁迫红色型豌豆蚜8代的体重变化
Table 2Effects of different time periods UV-B stress on weight of red pea aphid adult for 8 generations

世代Generation体重/mg WeightCK20min30min40min50min60minF1(3.11±0.18)aA(2.86±0.04)abB(2.61±0.04)bcAB(2.28±0.10)cdBC(2.10±0.23)dAB(1.48±0.07)eAF2(3.08±0.09)aA(2.89±0.04)aAB(2.79±0.24)aA(2.27±0.15)bBC(2.23±0.12)bAB(1.51±0.04)cAF3(2.98±0.13)aA(2.98±0.03)aAB(2.85±0.08)aA(2.37±0.15)bAB(2.28±0.07)bAB(1.59±0.04)cAF4(3.07±0.25)aA(3.10±0.15)aA(2.87±0.06)aA(2.63±0.06)abA(2.34±0.16)bA(1.28±0.04)cBF5(3.10±0.12)aA(2.86±0.07)bB(2.52±0.04)cAB(2.34±0.05)cABC(1.98±0.10)dAB(1.22±0.03)eBCF6(3.12±0.09)aA(2.83±0.03)bB(2.27±0.08)bBC(2.01±0.04)dC(1.88±0.09)dB(1.17±0.11)eBCF7(3.09±0.12)aA(2.51±0.05)bC(2.03±0.11)cCD(1.41±0.14)dD(1.28±0.07)deC(1.06±0.08)eCF8(3.05±0.19)aA(2.23±0.09)bD(1.77±0.13)cD(1.36±0.32)dD(1.16±0.06)deC(1.03±0.01)eC
2.3连续UV-B胁迫对豌豆蚜平均世代周期的影响
由表3可以看出,相同世代红色型豌豆蚜的平均世代周期随着UV-B辐射时间的延长表现出延长趋势。在UV-B的胁迫下,各世代豌豆蚜的平均世代周期除20 min处理外,其他处理的F3~F8代均是逐渐延长的。各处理下F1代平均世代周期虽然随着处理时间的延长而延长,但是差异不显著(P>0.05),F2代40~60 min处理下平均世代周期总体比20~30 min处理的长。F8代平均世代周期在各处理下均为最长,随辐射时间的延长,分别为对照的1.28、1.20、1.34、1.35、1.38倍。
平均世代周期在同一胁迫时间下随着世代增加均先缩短后延长。20~50 min处理下F4代组内最短, 60 min处理下F2代组内最短,但均较对照长。
2.4连续UV-B胁迫对豌豆蚜净增殖率的影响
由表4可以看出UV-B胁迫对红色型豌豆蚜的净增殖率产生了影响,受UV-B 胁迫后,与对照相比净增殖率显著下降(P<0.05)。同一世代中,胁迫时间越长,净增殖率越小。胁迫产生的影响积累至F8代时,各处理间差异显著(P<0.05),随着辐射时间的延长净增殖率分别为对照的46.55%、32.94%、 27.64%、22.63%、6.94%。
同一UV-B处理时间下净增殖率随着世代数增加先增大后减小,20~50 min处理下净增殖率以F4代最大,随后逐渐减小, 60 min处理下以F2代最大,随后递减。

表3 不同UV-B辐射时间连续胁迫红色型豌豆蚜8代的平均世代周期变化

表4 不同UV-B辐射时间连续胁迫红色型豌豆蚜8代的净增殖率变化
2.5连续UV-B胁迫对豌豆蚜内禀增长率的影响
由表5可以看出,在UV-B胁迫下,豌豆蚜内禀增长率呈降低趋势。同一世代下,随着辐射时间延长内禀增长率降低,除F4代20 min处理外,相同世代各处理的内禀增长率均低于对照,且各世代40、50、60 min处理下明显降低(P<0.05)。
同一UV-B处理时间下内禀增长率随着世代数的增加先增大后减小,20~50 min处理下内禀增长率增大至F4代后逐代显著减小(P<0.05),60 min处理下F2代后显著减小。50 min处理下,由于胁迫产生的遗传积累,F8代内禀增长率小于同世代对照的50%,60 min处理下仅为对照的26.20%。
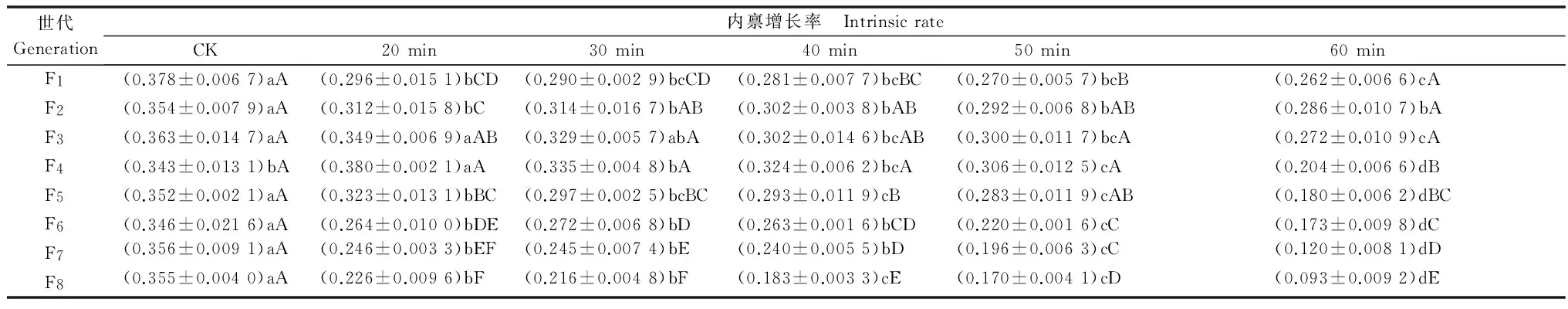
表5 不同UV-B辐射时间连续胁迫红色型豌豆蚜8代的内禀增长率变化
2.6连续UV-B胁迫对豌豆蚜周限增长率的影响
相同世代各处理下的周限增长率均是随着辐射时间的延长而降低,除F4代的20 min处理外,其他世代各处理均低于对照,50、60 min处理下各世代周限增长率明显降低(P<0.05)。
同一处理下周限增长率随着世代数的增加先上升后下降,20~50 min处理下F4代最高,60 min处理下F2代最高。各UV-B处理时间下F8代周限增长率均为最低,依处理时间长短分别为对照的87.94%、87.03%、 84.22%、83.10%、77.00%。因此长期UV-B胁迫不利于豌豆蚜种群数量的增长。

表6 不同UV-B辐射时间连续胁迫红色型豌豆蚜8代的周限增长率变化
3 结论与讨论
由于大气污染,导致臭氧层消耗,致使中高纬度地区UV-B辐射显著增加[2],增加的UV-B辐射对地球生态系统产生了影响[26],导致蚜虫种下遗传分化现象普遍[4],牧草害虫豌豆蚜也不例外,紫外胁迫上调了蚜虫表达生长因子[7],致使线粒体基因COⅠ-Ⅱ突变[21],并且UV-B辐射降低田间麦蚜复合种群数量[27-28]等。本研究采用小剂量的UV-B连续胁迫单克隆系红色型豌豆蚜8代,以观察UV-B胁迫对豌豆蚜生物学特性的影响,最终获得了连续多代种群生命表,统计出净增殖率、内禀增长率、平均世代周期和周限增长率等参数。研究结果表明,UV-B胁迫对豌豆蚜的体长、体重,净增殖率、内禀增长率、平均世代周期和周限增长率都产生了一定的影响。不同时间UV-B辐射连续胁迫红色型豌豆蚜8代后,F8代成虫平均体长显著减小,随着辐射时间的延长依次为对照的81.94%、79.03%、69.68%、61.61%、42.58%;成虫平均体重显著减轻,随着辐射时间的延长依次为对照的73.12%、58.03%、44.59%、38.03%、33.77%。在同一辐射时间处理下,两者均是随着世代数的增加先增大后减小,20~50 min处理下,体长和体重增大至F4代后逐代迅速减小,60 min处理下增大至F3代后逐代迅速减小。体长和体重是国外学者评价环境对昆虫影响的指标,比生命表更简单直接[29],因此在UV-B胁迫下,最终豌豆蚜体长缩短,体重减轻表明UV-B辐射抑制豌豆蚜的生长,与杜军利的研究结果相似[18],并且这种抑制是逐世代加强的,说明UV-B辐射产生的影响能够遗传积累[21,24],这种遗传积累很可能是由于紫外辐射诱发了基因变异而产生的[30-31]。处理组F1~F4代内体长和体重均有不同程度的增加,这可能与其抗逆反应过程中保护酶活性的上升有关[32],也可能是低剂量UV-B胁迫诱导豌豆蚜生理生化反应而产生的“毒物兴奋效应[33]”,这种效应与“毒物”剂量有关,高剂量下表现为抑制作用。净增殖力(R0)、平均世代周期(T)、内禀增长率(rm)和周限增长率(λ)是评价特定环境中种群变化的重要参数[34]。本研究中,红色型豌豆蚜受UV-B连续胁迫8代后,其F8代净增殖率与对照相比显著降低,随着辐射时间的延长分别为对照的46.55%、32.94%、 27.64%、22.63%、6.94%;F8代周限增长率相对于对照显著减小,分别为对照的87.94%、87.03%、 84.22%、83.10%、77.00%;由于胁迫产生的遗传积累,50~60 min 处理下F8代内禀增长率均小于对照的1/2,说明此时豌豆蚜种群增长呈衰退趋势;F8代平均世代周期相对于对照显著增长,随辐射处理时间延长分别为对照的1.28、1.20、1.34、1.35、1.38倍,以上说明UV-B胁迫抑制了豌豆蚜种群数量的增长,研究结果与Nigel等[3]对桃蚜的研究结果相似。处理组内随着世代的增加,净增殖率、内禀增长率和周限增长率均是先增大后减小,平均世代周期先缩短后显著延长,各种群参数在20~50 min处理下均以F4代为转折点,60 min处理下以F2代为转折点。出现这种现象可能与豌豆蚜的抗逆反应有关[5-6]。李进步等[35]认为,当遇到紫外辐射、高温等恶化环境时,蚜虫会出现体色分化应对,进一步通过调节生理代谢来提高机体能量,进而以改变其繁殖力来应对。
综上所述,长期UV-B胁迫对豌豆蚜的生长发育和种群增长都有抑制作用,具体表现为:体长减短,体重减轻,净增殖率、内禀增长率和周限增长率降低,平均世代周期延长。本试验所有试验结果均是在实验室理想条件下获得的,仅就连续多代UV-B胁迫下豌豆蚜的种群参数做了研究,对UV-B诱导产生负作用的机理探究不明确,但仍然可以为紫外诱导豌豆蚜种群演替和害虫综合防治提供理论依据。若要完全明确UV-B胁迫在豌豆蚜种群演替和进化中的作用,还需从分子机理、酶及相关蛋白和DNA、RNA等方面做进一步研究。
[1]王少彬,苏维瀚, 魏鼎文.太阳紫外线的生物有效辐射与大气臭氧含量减少的关系[J].环境科学学报,1993(1):114-120.
[2]Madronich S, McKenzie R L, Caldwell M M, et al. 地表紫外辐射的变化[J].李卫,译.AMBIO-人类环境杂志,1995,24(3):143-152.
[3]Nigel D P, Jason P M, McPherson M, et al. Ecological responses to UV radiation: interactions between the biological effects of UV on plants and on associated or ganisms [J]. Physiologia Plantarum, 2012, 145 (4):565-581.
[4]Thieme T.Adaptive significance of brown coloration inSitobionavenae[J]. IOBC/WPRS Bulletin, 1998, 21(2): 7-14.
[5]Yeen Y W, Chai L H, Phuoc D N, et al. Isolation of salinity tolerant genes from the mangrove plant,Bruguieracylindricaby using suppression subtractive hybridization (SSH) and bacterial functional screening [J]. Aquatic Botany,2007, 86 (2):117-122.
[6]杨传平,王玉成,刘桂丰,等.应用SSH 技术研究NaHCO3胁迫下柽柳基因的表达[J]. 遗传学报, 2004, 31 (9):926-933.
[7]Chu R, Varanasi U, Chu S, et al. Over expression and characterization of the human peroxi-somal acyl-CoA oxidase in insect cells[J]. Journal of Biological Chemistry, 1995, 27(4):4908-4915.
[8]Ramos S, Moya A, Martínez-Torres D. Identification of a gene overexpressed in aphids reared under short photoperiod [J].Insect Biochemistry and Molecular Biology,2003,33(3):289-298.
[9]Duman J G, Patterson J L.Role of thermal-hysteresis-proteins in low-temperature tolerance of insects and spiders [J].Cryobiology, 1978, 15 (6):683-684.
[10]Gazi Görür. Developmental instability in cabbage aphid (Brevicorynebrassicae) populations exposed to heavy metal accumulated host plants [J]. Ecological Indicators, 2006,6(4):743-748.
[11]Altincicek B, Knorr E, Vilcinskas A L.Beetle immunity: Identification of immune inducible genes from the model insectTriboliumcastaneum[J]. Developmental and Comparative Immunology, 2008, 32(4):585-595.
[12]武德功,杜军利,王森山,等.4个苜蓿品种对豌豆蚜的抗性评价[J].草业科学,2012(1):101-104.
[13]武德功, 贺春贵, 刘长仲, 等. 不同苜蓿品种对豌豆蚜的生化抗性机制[J]. 草地学报, 2011, 19(3):497-501.
[14]Charles H, Godfray J.The pea aphid genome [J]. Insect Molecular Biology, 2010, 19(2): 1-4.
[15]He C G, Zhang X G.Field evaluation of lucerne (MedicagosativaL.) for resistance to aphids in northern China [J]. Australian Journal of Agricultural Research, 2006, 57(4): 471-475.
[16]高有华, 刘长仲. 豆无网长管蚜的发育起点温度和有效积温测定[J].草原与草坪,2007(2):25-27.
[17]金娟, 王森山, 贺春贵. 西北不同地理种群红色型豌豆蚜的遗传多样性[J]. 草地学报, 2013, 21(2):406-412.
[18]杜军利, 武德功, 张廷伟, 等. 不同紫外辐射时间对2种色型豌豆蚜后代生物学特性的影响[J]. 草地学报, 2012, 20(5):961-966.
[19]胡祖庆,亢菊侠, 赵惠燕, 等. 不同紫外(UV-B)辐射时间对两种体色型麦长管蚜后代生物学特征的影响[J].生态学报, 2010, 30(7):1812-1816.
[20]姚建秀, 赵惠燕. 紫外条件诱导下麦长管蚜DNA的变异研究[J]. 西北农业学报, 2001, 10(1): 33-36.
[21]都二霞, 赵惠燕, 郭剑文. 紫外线诱导对桃蚜生态学及线粒体基因COⅠ-Ⅱ突变的影响[J]. 西北农林科技大学学报(自然科学版), 2007, 35 (9):123-126.
[22]张迎梅, 李元, 王勋陵, 等. 紫外辐射增强对麦二叉蚜发生量的影响[J]. 环境科学, 1999, 20(3):66-68.
[23]胡祖庆,赵惠燕,亢菊侠,等.紫外辐射对两种体色型麦长管蚜生物学特征的影响[J].应用生态学报,2009,20(4):1002-1006.
[24]都二霞, 郭剑文, 赵惠燕. 紫外辐射诱导桃蚜DNA变异[J]. 应用生态学报, 2006, 17(7): 1245-1249.
[25]周东, 杜一民, 杨杰, 等. 连续多代UVB照射对麦长管蚜保护酶活性的影响[J]. 昆虫学报, 2014, 57(7):762-768.
[26]Wilson S R, Solomon K R, Tang X.Changes in tropospheric composition and air quality due to stratospheric ozone depletion and climate change [J]. Photochemical & Photobiological Sciences, 2007, 6(3):301-310.
[27]李元, 王勋陵. 增强的UV-B辐射对麦田生态系统中种群数量动态的影响[J]. 生态学报, 2001, 21(1):131-135.
[28]张迎梅, 李元, 王勋陵, 等. 紫外辐射增强对麦二叉蚜发生量的影响[J]. 环境科学, 1999, 20(3):66-68.
[29]Caillaud C M, Dedryver C A, Simon J C.Development and reproductive potential of the cereal aphidSitobionavenaeon resistant wheat lines (Triticummonococcum)[J]. Annals of Applied Biology, 1994, 125(2):219-232.
[30]余多慰,柯惟中. 水溶液中DNA紫外辐射损伤的分子机制研究[J]. 光谱学与光谱分析, 2000, 20(3):311-314.
[31]郝卫东,根岸和雄.太阳光及紫外线B诱发基因突变的碱基特异性[J].北京医科大学学报,1997(2):121-123.
[32]李军, 赵惠燕, 赵学达. 不同强度紫外线对蚜虫生态学特征及有关酶活性的影响[J]. 西北农林科技大学学报(自然科学版), 2005, 33(4):61-64.
[33]Stebbing A R D. Hormesis-the simulation of growth by low levels of inhibitors [J].Science of the Total Environment,1982,22(3):213-234.
[34]刘长仲, 杜军利, 张廷伟, 等. 温度对三叶草彩斑蚜种群参数的影响[J]. 应用生态学报, 2012, 23(7):1927-1932.
[35]李进步,方丽平,孟玲,等.蚜虫体色多态性及生态进化功能[J].生态学杂志,2014(5):1404-1412.
(责任编辑:田喆)
Effects of UV-B stress on biological characteristics of red pea aphidAcyrthosiphonpisumHarris
Yuan Weining,Zhu Yangyan,Sun Xiaoling,Yang Qiaoyan,Liu Changzhong
(Pratacultural College of Gansu Agricultural University; Key Laboratory of Grassland Ecosystem of Ministry of Education, Lanzhou730070, China)
The depletion of ozone resulted in enhanced UV-B stress in middle and high latitudes of the northern or southern hemisphere. These situations generally made genetic divergence of aphids. This study sought to determine the effects of UV-B stress on biological properties of the red pea aphid and on species succession and subspecies differentiation. The newly born red pea aphids were treated for 8 generations by 40 W UV-B radiation, once a day, 6 times at 20 min, 30 min, 40 min, 50 min and 60 min. The results indicated that, with UV-B radiation time prolonged, the average body length of red pea aphids cut back, with average weight lightened, net reproductive rate, finite rate of increase and intrinsic rate reduced, and the mean generation time was prolonged in the same generation. Continuous UV-B radiation made average body length, average weight, net reproductive rate, finite rate of increase and intrinsic rate all increase in the first four generations, then reduce greatly with generation increasing, but the mean generation time declined at first, then increased. Up to F8, the average body length, average weight, net reproductive rate, finite rate of increase and mean generation time were 42.58%-81.94%, 33.77%-73.12%, 6.94%-46.55%, 77.00%-87.94%, and 1.20-1.38 times of the CK.However, the intrinsic rate decreased to 1/2 of the CK under the treatments of 50 min and 60 min, indicating that the population quantity was negative increase. All these data suggested that a long period of UV-B stress might lead to inhibitory effects on development and population growth of the red pea aphid.
UV-B;Acyrthosiphonpisum;life table;parameter of population dynamics;intrinsic rate of increase
2015-07-07
2015-09-22
国家自然科学基金(31260433) ;甘肃省农牧厅科技项目;高等学校博士学科点专项科研基金博导类资助课题(20136202110007)
E-mail:liuchzh@gsau.edu.cn
S 435.24
A
10.3969/j.issn.0529-1542.2016.04.011
猜你喜欢
天天爱科学(2022年9期)2022-09-15 01:12:28
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
安徽农业科学(2019年2期)2019-06-11 05:31:42
河北渔业(2018年7期)2018-09-15 08:59:42
健康必读·下旬刊(2018年6期)2018-07-24 09:50:24
大灰狼(2018年5期)2018-06-20 14:49:32
课外生活(小学1-3年级)(2017年5期)2017-06-10 10:38:03
故事作文·高年级(2016年9期)2016-10-25 08:50:53
故事作文·高年级(2015年1期)2015-09-07 08:21:57
