
爪鲵端脑胚后发育的组织学研究
2016-08-31 02:21:28王欢欢邓昕旸梁传成王丽文
湖南师范大学自然科学学报 2016年3期
王 维,王欢欢,邓昕旸,梁传成,2,王丽文,2*
(1.沈阳师范大学化学与生命科学学院,中国 沈阳 110034;2.辽宁省高校生物系统进化与农业生态实验室,中国 沈阳 110034)
爪鲵端脑胚后发育的组织学研究
王维1,王欢欢1,邓昕旸1,梁传成1,2,王丽文1,2*
(1.沈阳师范大学化学与生命科学学院,中国 沈阳110034;2.辽宁省高校生物系统进化与农业生态实验室,中国 沈阳110034)
为探讨爪鲵端脑胚后发育的特征,及其在系统进化中的地位,采用脊椎动物神经标本制作技术和HE染色方法,对爪鲵胚后6个年龄组端脑的形态学与组织学进行研究.发现爪鲵端脑由嗅球和大脑半球组成,从幼体Ⅰ龄开始,爪鲵已具备了和成体端脑相似的结构.随发育的进行,爪鲵的端脑长和整个脑长都是逐渐增加的,端脑占整个脑的比例逐步加大.爪鲵的原始大脑皮质不发达,与原始海马和原始梨状区之间的界限不明显;原始海马是爪鲵大脑半球中最发达的结构,发育至Ⅱ龄时,出现内侧界沟将其与隔区分开;外侧界沟在Ⅰ龄时就将原始梨状区和纹状体分开,但发育至Ⅲ龄时,外侧界沟变得不明显,纹状体突增大.而东北小鲵没有明显的外侧界沟,这表明爪鲵的大脑半球虽然比较原始,但高于同类的东北小鲵.但与其他两栖动物相比,爪鲵的端脑结构还较为原始,在两栖类中属于较低等的类群.
爪鲵;端脑;胚后发育;组织学
爪鲵(Onychodactylusfischeri)隶属于两栖纲,有尾目,小鲵科,爪鲵属[1].由于爪鲵成体肺完全退化消失,指、趾具角质的爪,尾部形态更接近于爬行动物蜥蜴,因此爪鲵在进化地位上具有一定的特殊性.其分布范围狭窄,数量有限,先后被世界自然资源保护联盟红皮书、前苏联红皮书、中国濒危动物红皮书列为重点保护的濒危物种[2].有关爪鲵神经系统结构和发育的研究,仅限于对亚成体和成体爪鲵端脑组织学的描述[3-4].本文对爪鲵端脑的形态学与组织学的胚后发育进行了较为系统的研究,以期探讨爪鲵大脑半球发育的特征及进化地位,丰富有尾两栖动物发育的资料.对于神经系统发育的研究有助于我们更好地掌握爪鲵发育的特点,对拓宽爪鲵的生存环境、更好地对其进行保护有重要意义.
神经系统的发育是生物个体发育中的重要组成部分,而端脑作为中枢神经系统重要的组成部分,它的发育程度在一定程度上表明了物种的进化程度[5].目前对端脑的形态结构、细胞分化、基因表达和神经网络特定功能等的研究都取得了丰富成果[6-9].研究者应用HE和Nissl染色法、Golgi-Ⅱ eHHeke 浸银法,对有尾两栖类的大鲵[10]、北方山溪鲵[11]和东北小鲵[12-13]的端脑进行了组织学研究,认为有尾两栖类最发达的部位是原始海马,原始大脑皮层很不发达,只在背侧有少数的几层细胞,且难以与原始梨状区相区分,而爬行类的原始大脑皮质已经分化出3个明显的区域[14-15].但同鱼类端脑表皮相比,两栖类端脑表皮更为发达,非洲肺鱼的端脑则与两栖类动物相类似[16-18].
1 材料与方法
1.1实验材料
实验用处于3个不同发育阶段共6个年龄组的爪鲵49只[19],均在不同的季节采自辽宁省鞍山市岫岩县三家子镇华山村.于实验室饲养在6~10 ℃的恒低温双层养殖箱中[20].
1.2实验方法
常规方法解剖分离爪鲵完整的脑,常规石蜡切片,HE染色,用Olympus CX21光学显微镜观察, Moticam1300数码显微图象系统拍照,Illustrator软件对端脑区域进行重构并绘图.区域的命名参照Hoffman和张育辉的研究[9-10].用测微尺测量各年龄组爪鲵原始大脑皮层、原始海马和原始梨状区神经元分布区域的厚度.
实验结果用SPSS统计软件进行统计分析,绘制柱形图.数据用均数±标准差(X±SD)表示.P<0.05即认为差异极显著.
2 结果
爪鲵从幼体Ⅰ龄开始,已具备了和成体端脑相似的结构.随发育的进行,爪鲵的端脑长和整个脑长均是逐渐增加,端脑占整个脑的比例逐步加大.
2.1嗅球
Ⅰ龄和Ⅱ龄爪鲵嗅球的细胞分层现象不明显,Ⅲ~Ⅵ龄,嗅球细胞的分层逐步清晰,但各层细胞并无明确的界限.整体上看,嗅球腹外侧部细胞分布稀疏,向内逐渐紧密(图1).
2.2大脑半球
爪鲵的大脑半球由原始大脑皮层、原始海马、隔区、原始梨状区、纹状体等区域以及侧脑室和存在于侧脑室内的脉络丛等构成.爪鲵不同的发育阶段,大脑半球的各区域都有着不同的变化.
原始大脑皮层位于侧脑室的背侧部,其神经细胞在Ⅰ~Ⅲ龄爪鲵大脑半球内分布比较均匀,几乎占据侧脑室背侧的整个区域.Ⅳ~Ⅵ龄爪鲵的原始大脑皮层细胞聚集于侧脑室背侧相对狭窄的区域,上层细胞相对分散(图1,彩图见封三).就其细胞分布区域的厚度而言,随着发育的进行其皮层厚度增加,Ⅳ龄和Ⅴ龄的皮层厚度虽然较Ⅲ龄减小,但仍高于Ⅰ龄和Ⅱ龄,Ⅵ龄皮层厚度最高约197 μm(图2).

图1 爪鲵胚后发育各阶段大脑半球横切(HE染色)(o,嗅球;pal,原始大脑皮层;ph,原始海马;pr,原始梨状区; se,隔区;sp,纹状体;mls,内侧界沟;lls,外侧界沟)Fig.1 Transverse sections through the cerebral hemisphere of Onychodactylus fischeri in each age groups after embryo (HE-staining)( o, olfactory bulb; pal, primordial pallium; ph, primordial hippocampus; pr, primordial piriform; se, septum; sp, striatal complex; mls, medial limit sulcus; lls, lateral limit sulcus)
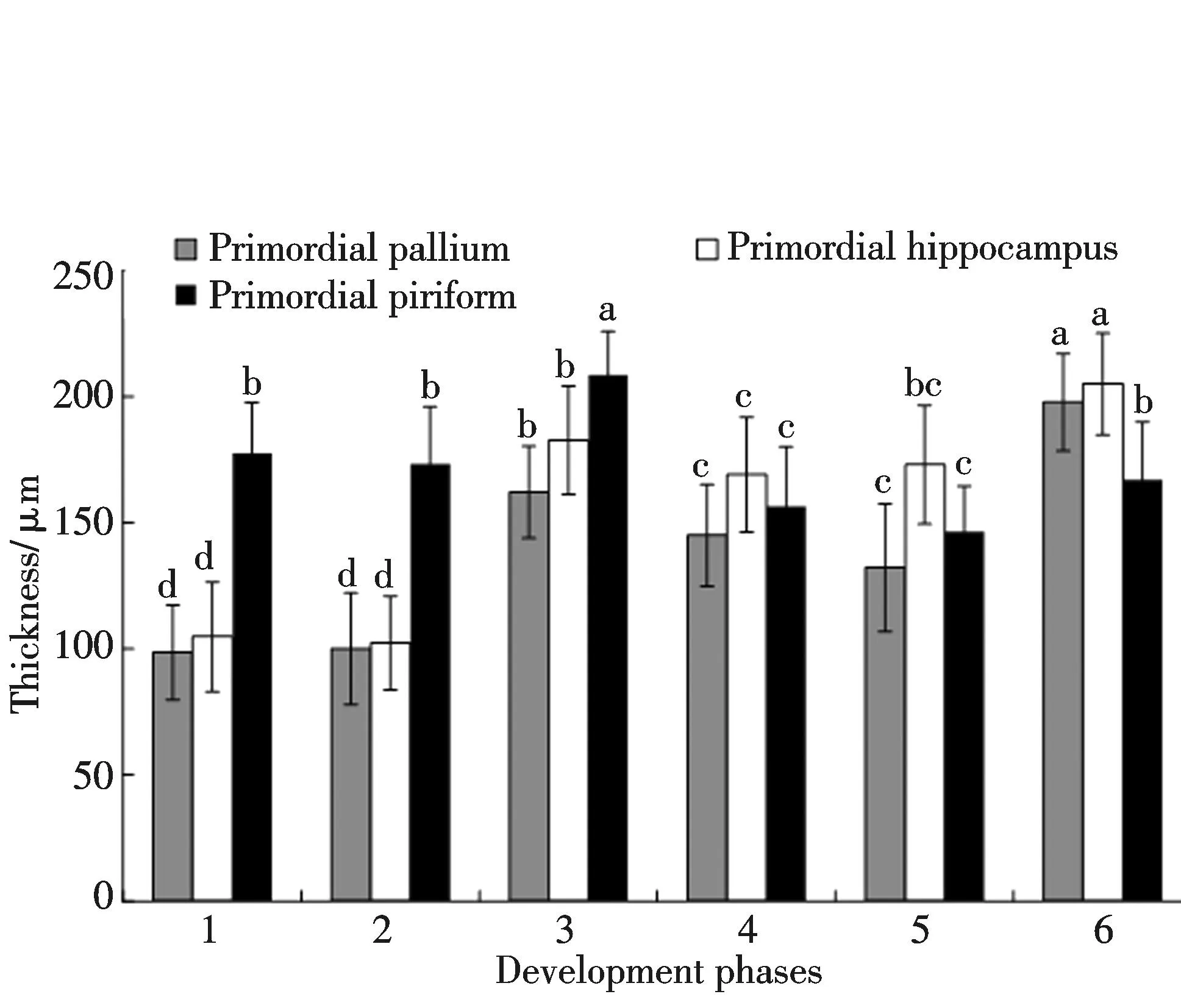
图2 爪鲵胚后发育各阶段端脑各细胞分布区域厚度变化比较(a, b, c, d表示差异显著)Fig.2 Comparison of cell distributing thickness changes of telencephalon of Onychodactylus fischeri in each age groups after embryo(a, b, c, d show significant difference)
原始海马位于侧脑室内侧壁的上部,背侧与原始大脑皮层相连.隔区则位于原始海马的下方.Ⅰ~Ⅵ龄爪鲵端脑的前端,原始海马与隔区无明显的分界(图1),随着侧脑室加宽加长,侧脑室内出现内侧界沟将原始海马和隔区分割开来.Ⅰ龄爪鲵的内侧界沟不甚明显(图1),从Ⅱ龄开始,在大脑半球的中段,侧脑室出现明显的内侧界沟(图1).从Ⅲ龄开始,内侧界沟逐渐移向中下部.在大脑半球的尾端,两脑室将要连通的部位,内侧界沟消失.原始海马的细胞分布区域厚度在各个年龄组的变化趋势与大脑皮层大致相似(图2).
原始梨状区位于侧脑室的外侧,其细胞分布区域厚度,Ⅲ龄大于Ⅰ龄和Ⅱ龄,Ⅳ龄和Ⅴ龄又减小,Ⅵ龄略增加,但仍不及Ⅲ龄(图2).纹状体是位于原始梨状区下方和侧脑室底部比较大的核团.Ⅰ龄和Ⅱ龄爪鲵的大脑半球上可见有外侧界沟将原始梨状区和纹状体分开(图1),在Ⅲ~Ⅵ龄,外侧界沟不明显,由纹状体突区分二者.幼体的纹状体突大于成体,Ⅱ龄爪鲵的纹状体突较为明显(图1),细胞厚度明显大于梨状区.
3 讨论
3.1嗅球
嗅球是低等动物感知外界环境的重要器官之一.爪鲵的嗅球与大脑半球并无明显分界,没有发现副嗅球.而一些无尾两栖类的嗅球与大脑半球界限清晰,且存在副嗅球[21].这说明爪鲵的端脑在进化上较无尾两栖类原始,没有分化出独立的嗅觉中枢[3].从系统进化的关系来看,鱼类较两栖类原始,但在鱼类的端脑中,嗅球以嗅茎和大脑半球相连,有独立的嗅觉器官[22],由此来看爪鲵的嗅觉器官甚至比鱼类原始.
3.2大脑半球
同大鲵、山溪鲵、东北小鲵以及无尾两栖类的原始大脑皮质相似[10-13],爪鲵的原始大脑皮质并不发达,与原始海马和原始梨状区之间的界限并不明显,细胞层较薄.而爬行类鳄鱼的大脑皮质已属于新皮层[23-24],皮层较厚,可细分为外网状层、细胞层和内网状层.草原沙蜥和敏麻蜥的皮质区更为发达[25],分为内皮层、背皮层和侧皮层三部分.由此可见爪鲵的皮质区仍处于较为原始的阶段.
爪鲵大脑半球中最发达的结构是原始海马,与山溪鲵[11]和东北小鲵[12-13]相似.原始海马区属于原脑皮,从大脑半球前端侧脑室位置出现,到第三脑室处逐渐消失.无尾两栖类的原始海马可以区分为两个细胞群,背侧群和背内侧群,分别包含不同类型的细胞.爪鲵的原始海马细胞很难区分为两部分,这也说明了爪鲵端脑的原始性.
原始海马和隔区及原始梨状区和纹状体的分界分别为内侧界沟和外侧界沟.无尾两栖类的两界沟清晰可见,隔区分为内侧隔核和外侧隔核两个核团[24].爪鲵的两个界沟不及无尾类清晰,隔区也没有再分成两个核团.而东北小鲵没有明显的外侧界沟,这表明爪鲵的大脑半球虽然比较原始,但就隔区与纹状体的分界上高于同类的东北小鲵.
[1]赵尔宓,胡其雄.中国有尾两栖动物的研究[M].成都:四川科学技术出版社, 1984.
[2]赵尔宓.中国濒危动物红皮书:两栖类和爬行类[M].北京:科学出版社, 1998.
[3]WANG H H, LI L Y, WANG L W,etal. Morphological and histological studies on the telencephalon of the salamander Onychodactylus fischeri[J]. Neurosci Bull, 2007,23(3):170-174.
[4]王欢欢,李丽妍,王丽文,等.亚成体和成体爪鲵端脑组织学初步研究[J].四川动物, 2008,27(2):223-226.
[5]杨岐生,林卿.脊椎动物前脑发育的研究进展[J].细胞生物学杂志, 1996,18(4):160-164.
[6]ENDO T, YOSHINO J, KADO K,etal. Brain regeneration in anuran amphibians [J]. Dev Growth Differ, 2007,49 (2):121-129.
[7]COUMAILLEAU P, KAH O. Cyp19a1 (Aromatase) expression in the xenopus brain at different developmental stages[J]. J Neuroendocrinol, 2014,26(4):226-236.
[9]周奕,邓志伟,艾卫敏,等. Aβ在两种阿尔茨海默病转基因小鼠模型脑杏仁核分布的比较研究[J]. 湖南师范大学自然科学学报, 2014,37(5):26-30.
[10]张育辉,梁刚.中国大鲵端脑的组织学研究[J].两栖爬行动物学研究,1995(4-5辑):30-34.
[11]蒋一淼,张育辉. 北方山溪鲵端脑细胞构筑的组织学观察[J].陕西师范大学学报(自然科学版), 2003,31(4):80-83.
[12]李丽妍,王丽文,那杰,等.东北小鲵中枢神经系统形态学与组织学初步研究[J].四川动物, 2005,24(3):287-289.
[13]ZHOU Y Y, SHAO R, LIANG C C,etal. Histological studies on the telencephalon of Hynobius leechii at the metamorphosis phase and the adult phase [J]. Neurosci Bull, 2009,25(4):196-202.
[14]唐宗湘,姜世英,施敏,等.蛤蚧端脑前背侧室嵴Golgi-Cox研究[J]. 解剖学研究, 2000,2(1):1-4.
[15]沈端文,张杏辉,姜世英,等.蛤蚧端脑前背侧室嵴头外侧区突触的电镜研究[J]. 电子显微学报, 1998,17(3):213-219.
[16]NORTHCUTT R G, WESTHOFF G. Connections of the medial telencephalic wall in the spotted african lungfish[J]. Brain Behav Evol, 2011,77(1):14-32.
[18]NORTHCUTT R G. Telencephalic organization in the spotted african lungfish, protopterus dolloi: a new cytological model[J]. Brain Behav Evol, 2009,73(1):59-80.
[19]毕远阁,廖承林,孙林,等.两栖动物年龄划分方法的比较研究[J].生物学通报, 2009,44(6):18-19.
[20]王丽文,梁传成,黄薇,等.环境温度对爪鲵体温及能量代谢的影响[J].动物学报, 2008,54(4):640- 644.
[21]MORENO N, MORONA R, LPEZ J M,etal. Lateral and medial amygdale of anuran amphibians and their relation to olfactory and vomeronasal information[J]. Brain Res Bull, 2005,66(4-6):332-336.
[22]MORGAN G C. The telencephalon of the sea catfish Galeichthys felis[J]. J Hirnforsch, 1975,16 (2):131-50.
[23]杨传秀,杨超,陈璧辉.初孵扬子鳄端脑基地中枢的组织学研究[J]. 安徽师范大学学报(自然科学版), 2002,25(3):270-273.
[24]CROSBY E C. The forebrain of alligator mississippiensis[J]. J Comp Neur, 1916,7(3):325-405.
[25]张服基,庞家庆,王跃招.两种蜥蜴大脑的组织学研究[J]. 两栖爬行动物学报, 1988(1-2):81-86.
(编辑WJ)
Histology Research on Postembryonic Development of Telencephalon in SalamanderOnychodactylusFischeri
WANG Wei1, WANG Huan-huan1, DENG Xin-yang1, LIANG Chuan-cheng1,2, WANG Li-wen1,2*
(1.College of Chemistry and Life Science, Shenyang Normal University, Shenyang 110034, China;2.Key Laboratory of Biological Evolution and Agricultural Ecology,Liaoning Provincial Department of Education, Shenyang 110034, China)
Objective: The characteristics of postembryonic development of the telencephalon in the salamanderOnychodactylusfischeriand its status in the evolution was discussed. Methods: The morphological and histological investigation on telencephalon ofOnychodactylusfischeriat six age groups were conducted using the vertebrate neural specimen making technology and HE-staining. Results: The telencephalon ofOnychodactylusfischericonsists of an olfactory bulb and cerebral hemisphere, from the GroupⅠ. It has a similar structure with the adult. With the development of the telencephalon, the whole length of brain and the length of telencephalon are gradually increased, and the proportion of the length of telencephalon in the whole length of brain increases gradually as well. The primordial cerebral cortex of theOnychodactylusfischeriis not developed, and the boundaries with primordial hippocampus and primordial piriform regions are not visible. The primordial hippocampus is the most developed structure in the cerebral hemisphere of theOnychodactylusfischerito the development of Group Ⅱ, and the medial limit sulcus appeared and separated it from the septal area. In the Group I, the primordial piriform region and striatum were separated by the lateral limit sulcus, but for the development to the Group Ⅲ, the lateral limit sulcus was not obvious as striatal complex projections were increased. While the Hynobius leechii does not have a clear lateral limit sulcus, which indicates that although the telencephalon of theOnychodactylusfischeriis relatively primitive, but higher than similar Hynobius leechii. When compared with other amphibians, the telencephalon structure of theOnychodactylusfischeriis relatively primitive, suggesting that theOnychodactylusfischeriin amphibians belongs to lower taxa.
salamander; telencephalon; postembryonic development; histology
10.7612/j.issn.1000-2537.2016.03.004
2015-11-29基金项目:辽宁省教育厅资助项目(2011196)*通讯作者,E-mail:wangliwen1956@163.com
Q959.5
A
1000-2537(2016)03-0018-04
猜你喜欢
数学年刊A辑(中文版)(2023年4期)2024-01-04 05:47:52
大自然探索(2020年7期)2020-08-06 15:25:32
中国妇幼健康研究(2019年2期)2019-03-26 07:14:46
中国化肥信息(2016年33期)2016-05-17 04:25:43
磁共振成像(2015年1期)2015-12-23 08:52:21
中国中西医结合影像学杂志(2015年1期)2015-06-06 10:45:40
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:38
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:38
山西医科大学学报(2014年6期)2014-11-21 07:39:10
中国神经精神疾病杂志(2014年1期)2014-03-01 03:23:27
