
豌豆不同耐铝品种根尖细胞壁果胶及其甲基酯化度的差异
2016-08-30 02:51李学文李亚林吴礼树萧洪东冯英明刘家友
植物营养与肥料学报 2016年3期
李学文, 李亚林, 杨 锦, 吴礼树, 萧洪东,冯英明, 刘家友, 喻 敏*
(1佛山科学技术学院园艺系, 广东佛山 528000; 2 华中农业大学资环学院, 湖北武汉 430070 )
豌豆不同耐铝品种根尖细胞壁果胶及其甲基酯化度的差异
李学文1, 2, 李亚林1, 2, 杨 锦1, 2, 吴礼树2, 萧洪东1,冯英明1, 刘家友1, 喻 敏1*
(1佛山科学技术学院园艺系, 广东佛山 528000; 2 华中农业大学资环学院, 湖北武汉 430070 )
【目的】研究豌豆不同品种耐铝性和根尖根段耐铝性与果胶及其甲基酯化间的关系,为进一步揭示植物耐铝机理以及耐铝性状的遗传改良提供依据。【方法】以豌豆品种Hyogo和Alaska为试验材料,采用Hoagland培养方式,测定了不同品种不同根段果胶含量、 果胶甲基酯化度和果胶甲酯酶活性,研究了其差异及原因。【结果】在15和30 μmol/L铝浓度胁迫条件下,豌豆品种Alaska根相对伸长率均显著高于品种Hyogo,同时有根尖05 mm和510 mm段有更少的胼胝质生成和累积,在30 μmol/L浓度下不同根段间均达到显著差异,同时品种Hyogo根尖02.5 mm和2.55.0 mm段铝含量均显著高于品种Alaska,说明品种Alaska和品种Hyogo间存在耐铝性差异,其中品种Alaska耐铝性高于品种Hyogo,即品种Hyogo为铝敏感品种,品种Alaska是耐铝品种。比较两者不同根段(02.5 mm、 2.55.0 mm和5.010.0 mm)的铝含量与果胶含量、 果胶甲基酯化度、 PME活性间的关系,发现耐铝品种不同根段中的铝含量均小于敏感品种,并且在02.5 mm和2.55.0 mm段间达到显著性差异; 根尖不同根段果胶糖醛酸含量大小依次为02.5 >2.55.0>5.010.0 mm,耐铝品种Alaska根尖细胞壁果胶和未甲酯化果胶含量均显著低于Hyogo,并且02.5 mm根段差异最大。根尖不同根段果胶甲基酯化度从根尖向上逐渐降低,并且耐铝品种Alaska高于铝敏感品种Hyogo,其中02.5 mm段间的差异达到显著水平;在对两个品种果胶甲基酯化酶(PME)活性进一步分析发现,PME活性大小依次为02.5>2.55.0>5.010.0 mm,两品种02.5 mm和2.55.0 mm根段间均达到显著差异。【结论】铝敏感品种Hyogo在02.5 mm和2.55.0 mm根段具有较高 PME活性和较低果胶甲基酯化程度。豌豆根尖果胶含量和甲基酯化度尤其是02.5 mm根段是豌豆耐铝性差异的重要原因;Alaska根尖细胞壁的果胶含量低和果胶甲基酯化度高(尤其是02.5 mm段)是其耐铝的重要机制。
豌豆; 耐铝性; 细胞壁果胶; 甲基酯化度; 果胶甲基酯化酶
铝是地壳中含量最丰富的金属元素,约占地壳总重量的7%,通常以难溶性的硅酸盐或氧化铝形式存在,对植物无毒害作用。当土壤的pH低于5.0时,部分铝就会以离子形式释放到土壤溶液中,微摩尔级的铝就可以在短时间内对根系的伸长产生抑制作用。铝毒是酸性土壤中限制作物生长的众多胁迫因素中最重要的因子。最近几十年来,科学家们虽然已对植物铝毒和耐铝性机理做了大量研究,但对其生理和分子机制仍缺乏足够的了解[1-2]。铝毒对植物的危害首先表现在抑制植物根系的生长及对养分和水分的吸收,从而最终影响作物的产量。Ryan等提出根尖是铝毒的主要作用位点,也是铝抑制根系生长的作用靶[3]。铝毒导致植物根尖生长受到抑制的原因主要是细胞伸长受到抑制;Al3+与根表皮和皮层细胞的细胞壁结合,对细胞壁的结构和功能(如细胞壁的伸展性等)产生影响[4]。植物根尖细胞壁作为植物响应铝毒环境的第一道屏障,既是铝毒害的最初位点,也是铝积累的主要部位[1, 5],在铝毒害机理和植物耐铝机制中起到重要作用。根系中30%90%的铝累积于根尖细胞壁,而细胞壁对铝的结合能力主要取决于细胞壁果胶含量及其果胶甲基化度[6-9],果胶甲基酯化度受果胶甲基酯酶(PME)调控。 Sivaguru 和Horst[10]对玉米根尖不同根段分别进行铝诱导时发现,玉米根尖12 mm根段比01 mm和23 mm根段受铝影响更大,如胼胝质的形成和铝的累积更多,而12 mm根段为过渡区(transition zone),恰好位于分生区和伸长区之间,提出过渡区远端是玉米根尖的敏感部位,同时也表明根尖不同区域可能对铝的敏感性不同。Yang等[7]对比不同耐铝性水稻品种发现,铝敏感型品种 Zhefu802 根尖010 mm段PME活性、 细胞壁多糖(果胶,半纤维素,半纤维素2)和果胶的去甲基化程度均显著高于耐铝型品种Nipponbare。导致根尖不同区域铝敏感性差异的原因是什么?是否与根尖细胞壁果胶含量及其甲基化程度有关?这种机制是否也引起不同品种耐铝差异?本研究以耐铝豌豆品种和铝敏感豌豆品种为材料,通过分析根尖不同根段果胶含量和甲基酯化程度及PME活性等,研究根尖细胞壁果胶含量和性质与根不同区域及不同豌豆品种耐铝差异的关系,以期更加深入地揭示铝对植物的毒害机理及植物耐铝机制。
1 材料与方法
1.1供试品种
豌豆(Pisumsativum)耐铝品种Alaska和铝敏感品种Hyogo由Yoko Yamamoto教授(Okayama University)馈赠。
1.2种子萌发
采用雾培法进行催芽[11]。
1.3测定方法
1.3.1 铝处理及其根伸长率的测定选取根长相对一致(约8.010.0 mm)的幼苗各3040株,测量根长后,转移至塑料烧杯中,分别用0、 15、 30 μmol/L AlCl3(含0.5 mmol/L CaCl2, pH 4.5) 处理24 h,再次分别测量根长。铝处理前根长分别记为LCK0和LAl0,铝处理24 h相对应根长分别记为LCK24和LAl24。根相对伸长率(root relative elongation rates,RER) 按下列公式计算:
1.3.2 胼胝质含量的测定剪取距根尖 05.0 mm、 5.010.0 mm的根段各5株,用4 mL 1 mol/L NaOH 研磨提取,收集研磨液于10 mL离心管中,于80℃水浴20 min,4800 rpm离心,收集上清液,加入苯胺蓝混合反应液,于50℃水浴20 min,冷却后采用荧光分光光度法(970CRT,上海),在λex=400/30nm,λem=485/40nm, 灵敏度为2,测定胼胝质含量[12]。1.3.3 铝处理及其铝含量的测定剪取距根尖 02.5 mm、 2.55.0 mm、 5.010.0 mm的根样各7株,用1 mL 2mol/L HCl 振荡提取48 h,4800 rpm 离心,收集上清液,采用ICP-AES法 (IRIS-Advantage, Thermo Elemental, MA, USA)测定铝含量。
1.3.4 PME的测定选取根长相对一致(约2025 mm)的幼苗,收集02.5 mm、 2.55.0 mm和5.010.0 mm根段,用于样品提取,参考Jolie等[13]的方法提取粗酶,按照Li等[8]的方法测定酶活性。
1.3.5 细胞壁果胶含量的测定参考Takayuki等[4]和Heim等[14]的方法稍作修改,对02.5 mm、 2.55.0 mm和5.010.0 mm根段进行细胞壁粗提取。在-20℃冷冻30 min,研磨使其达到均匀的状态,移入离心管,加入5 mL超纯水(预冷)淋洗,在4500 rpm条件下离心15 min,弃上清液,沉淀再用超纯水重复淋洗2次。然后将沉淀用5 mL丙酮(预冷)淋洗,室温下振荡浸提30 min,在4500 rpm条件下离心15 min,弃去上清液,沉淀再用丙酮重复淋洗2次。沉淀物即为细胞壁,真空干燥后备用。
用50 mmol/L Na2CO3(含20 mmol/L CDTA)提取细胞壁果胶,采用苯二酚比色法[15]测定果胶中糖醛酸含量,以D-半乳糖醛酸(Mr=212.16)为标准物,制备标准曲线。
1.3.6 细胞壁果胶甲基酯化度的测定吸取100 μL细胞壁果胶待测液,加入50 μL 1.5 mol/L NaOH,进行皂化,常温条件反应30 min,然后加入55 μL 0.75 mol/L H2SO4,参考PME测定方法[8],测定皂化反应后产生的甲醇。 以甲醛为标准物,制备标准曲线。
未甲基化果胶含量=果胶含量×(1-DM)。
1.4数据处理
结果为3次独立试验的平均值,采用Excel 2010和SAS 9.13进行数据处理和统计分析。
2 结果与分析
2.1铝对豌豆不同品种根伸长的影响
经15 μmol/L和30 μmol/L AlCl3溶液(含0.5 mmol/L CaCl2pH4.5)处理豌豆幼根24 h后,Alaska和Hyogo两个豌豆品种的根相对伸长率如图1所示。在不同浓度铝胁迫下,Alaska根相对伸长率均显著高于Hyogo,进一步验证Alaska和Hyogo分别为耐铝品种和铝敏感品种[16]。

图1 豌豆耐铝品种(Alaska)和铝敏感品种(Hyogo)根相对伸长率的差异Fig.1 Difference in root relative elongation rates under AlCl3 concentrations in Al resistant cultivar Alaska and Al sensitive cultivar Hyogo
2.2铝对豌豆不同品种根尖胼胝质含量的影响

图2 豌豆耐铝品种(Alaska)和铝敏感品种(Hyogo)不同根段的胼胝质含量差异Fig.2 Difference of callose content in root segments of Al resistant cultivar Alaska and Al sensitive cultivar Hyogo[注(Note): 柱上不同字母表示在0.05 水平上差异显著 Different letters above the bars mean significant difference at 0.05 level. ]
经15 μmol/L和30 μmol/L AlCl3溶液(含0.5 mmol/L CaCl2pH 4.5)处理根尖24 h后,Alaska和Hyogo根系不同根段胼胝质含量结果如图2所示。15 μmol/L AlCl3条件下,Hyogo不同根段胼胝质含量有大于Alaska的趋势,但未达到显著差异;30 μmol/L AlCl3浓度时,Hyogo根尖05.0 mm和5.010.0 mm段胼胝质含量均显著大于Alaska,分别是Alaska的2.3倍和2.7倍。
2.3豌豆不同品种根尖铝含量的差异

图3 豌豆耐铝品种(Alaska)和铝敏感品种(Hyogo)不同根段的铝含量差异Fig.3 Difference of aluminum content in root segments of Al resistant cultivar Alaska and Al sensitive cultivar Hyogo[注(Note): 柱上不同字母表示在0.05 水平上差异显著 Different letters above the bars mean significant difference at 0.05 level. ]
经30 μmol/L AlCl3溶液处理根尖24 h后,同一品种不同根段铝含量有从根尖向上递减的趋势(图3),其中Alaska品种 02.5 mm根段铝含量显著大于2.55.0 mm根段,Hyogo品种则 02.5 mm根段铝含量和2.55.0 mm根段中无明显差异,但均显著高于5.010.0 mm根段。不同耐铝性豌豆品种间根尖铝含量存在明显差异,Hyogo品种根尖02.5 mm和2.55.0 mm根段铝含量显著高于Alaska品种,分别为1.2倍和1.8倍, 5.010 mm根段差异不明显。
2.4豌豆不同品种根尖细胞壁果胶甲基酯化度的差异
果胶是铝结合到细胞壁的主要吸附位点,果胶结合铝的能力取决于果胶含量和果胶甲基酯化度。新生果胶一般都是高度酯化,只有脱甲基后,形成果胶酸,才能与铝结合,即果胶甲基酯化度越低,结合的铝吸附位点越多[6]。不同品种不同根段细胞壁中果胶甲基酯化度如图4所示。同一品种不同根段果胶甲基酯化度有从根尖向上依次递减的趋势,Alaska果胶甲基酯化度02.5 mm根段显著高于2.55.0 mm和5.010 mm根段,而Hyogo品种则差异不显著。不同豌豆品种间果胶甲基酯化度的差异主要是02.5 mm根段,Alaska品种 02.5 mm根段果胶甲基酯化度显著高于Hyogo品种,Alaska品种 2.55.0 mm和5.0 mm10.0 mm根段果胶甲基酯化度有高于Hyogo品种的趋势,但未达显著水平。

图4 豌豆耐铝品种(Alaska)和铝敏感品种(Hyogo)不同根段的果胶甲基酯化度的差异Fig.4 Difference of pectin methyl esterification degree in root segments of Al resistant cultivar Alaska and Al sensitive cultivar Hyogo
2.5豌豆不同品种根尖细胞壁中果胶和未甲酯化果胶含量的差异
果胶主要是由聚半乳糖醛酸构成,测定了不同根段的细胞壁中糖醛酸含量(图5)。同一品种不同根段果胶糖醛酸含量有从根尖向上依次递减的趋势,Hyogo品种各个根段间糖醛酸含量差异显著,Alaska品种 02.5 mm和2.55.0 mm根段无显著差异;不同豌豆品种间根尖糖醛酸含量存在显著差异,Hyogo品种不同根段糖醛酸含量均显著高于Alaska品种,分别是Alaska品种的1.8倍、 1.5倍和1.7倍。这些结果表明,铝敏感品种根尖细胞壁糖醛酸含量高于耐铝品种,尤其是02.5 mm根段两品种差异更大。
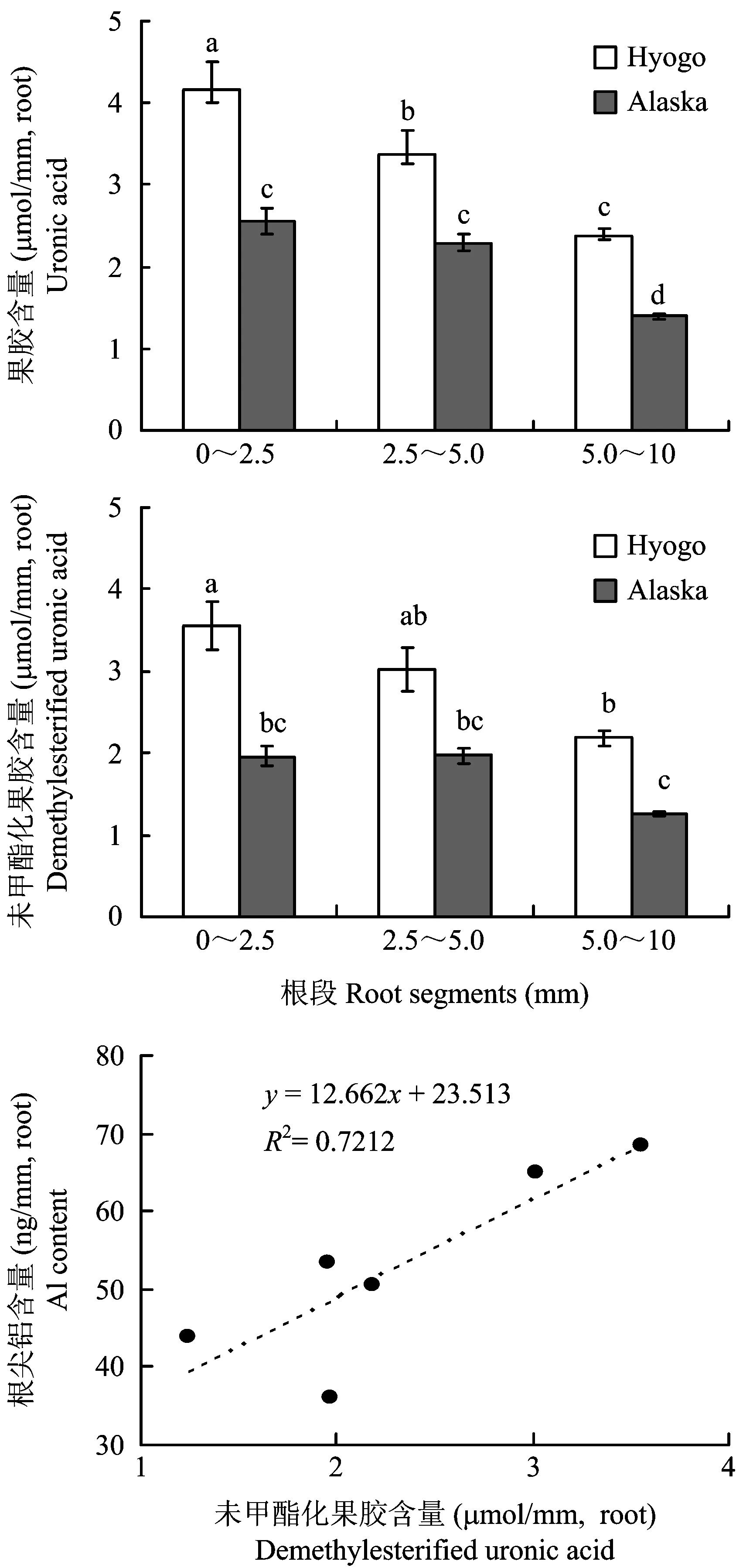
图5 豌豆耐铝品种(Alaska)和铝敏感品种(Hyogo)不同根段的糖醛酸含量的差异Fig.5 Difference of uronic acid content in root segments of Al resistant cultivar Alaska and Al sensitive cultivar Hyogo[注(Note): 柱上不同字母表示在0.05 水平上差异显著 Different letters above the bars mean significant difference at 0.05 level. ]
同一品种不同根段未甲酯化果胶糖醛酸含量有从根尖向上依次递减的趋势,Hyogo品种根尖不同根段未甲酯化果胶含量均高于Alaska品种,并且02.5 mm和5.010 mm根段差异达到显著水平,整体趋势与果胶含量保持一致。根尖未甲酯化果胶糖醛酸含量与根尖铝含量呈正比(图5),相关性达到显著水平,说明根尖铝含量与根尖未甲酯化果胶糖醛酸含量密切相关。
2.6豌豆不同品种根尖PME活性的差异
果胶甲基酯化度受果胶甲基酯化酶的控制,PME能使果胶去甲基化后转变为含有大量游离羧基的果胶酸,从而调节根尖细胞壁的果胶甲基酯化程度,间接地影响了植物对铝毒的耐性。Hyogo和Alaska品种根尖不同根段的PME活性均是从根尖向上活性依次降低,其中在02.5 mm和2.55.0 mm根段Hyogo品种显著高于Alaska品种(图6)。
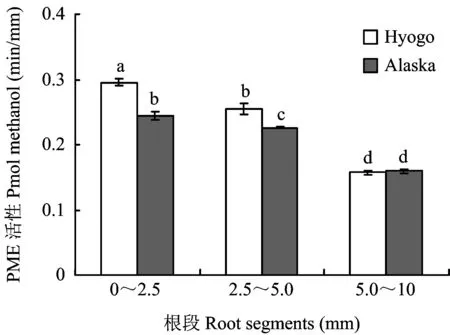
图6 豌豆耐铝品种(Alaska)和铝敏感品种(Hyogo)不同根段的PME活性的差异Fig.6 Difference in enzyme activity of PME in root segments of Al resistant cultivar Alaska and Al sensitive cultivar Hyogo
3 讨论与结论
3.1豌豆不同品种和不同根段的耐铝性差异
植物根系伸长的抑制是铝毒最敏感和最易观察到的典型症状,而胼胝质作为植物应激反应的一部分,在植物受伤或各种胁迫时能够快速、 大量地合成,由于铝毒作用会诱导胼胝质的大量产生,因而通常认为胼胝质含量是衡量根尖铝毒害程度的敏感指标[17]。本研究结果表明,在15 μmol/L和30 μmol/L AlCl3胁迫下,Alaska主根的根相对伸长率均大于Hyogo,并且胼胝质含量小于品种Hyogo,说明两品种在耐铝性上确实存在差异,其中Hyogo属铝敏感品种,Alaska属耐铝品种,结果与Kobayashi等[16]的一致。Sivaguru等提出根尖过渡区远端为铝敏感区域[10],在豌豆中,根尖02.5 mm根段包括根冠、 分生区和过渡区(李学文等,未发表资料),主要进行细胞有丝分裂和为细胞伸长做准备;2.55 mm根段为伸长区,主要进行植物细胞的伸长; 510 mm根段为成熟区,主要进行水和养分的吸收。本研究结果中,不同根段中的铝含量表现出由根尖向上依次递减,02.5 mm段根尖铝含量显著高于其他根段,是根尖铝敏感根段。
3.2豌豆不同根段铝敏感性与果胶含量和甲酯化程度的关系
植物根系的不同部位对铝毒的敏感性存在差异[18],耐铝性差异与果胶含量和果胶甲基酯化度密切相关[6-7, 19]。本研究表明,不同根段间果胶糖醛酸含量和果胶甲基酯化度在不同根段均存在差异,根尖不同根段中果胶含量和果胶甲基酯化程度均为02.5 mm>2.55.0 mm>5.05.0 mm,根段铝敏感性与果胶糖醛酸正相关,与果胶甲基酯化度负相关,与前人研究[6]并不完全一致。不同根段果胶含量和甲基酯化度的差异可能与不同根段所对应的根区及根区功能相关。越靠近根尖部位,植物生理活动越活跃,细胞的有丝分裂和细胞伸长过程都伴随大量细胞壁的生成,而新生果胶均为高度酯化,在PME作用下脱甲基,伴随细胞伸长成熟,果胶甲酯化度降低。因此我们认为不同根段间果胶含量和果胶甲基酯化度的差异是由不同根段及根段所对应区域的功能所决定的,鉴于02.5 mm区域功能和果胶含量最高,尽管其果胶甲基酯化度较高,未甲基酯化果胶依然最高,表明细胞壁果胶吸附铝的位点较多,分析表明铝累积与未甲基酯化果胶相关性高于与总果胶的相关性,因此根尖根段铝敏感性与未甲酯化果胶含量呈正比,02.5 mm根段铝敏感性最高。
3.3豌豆不同品种耐铝差异与果胶含量和甲酯化程度的关系
本研究表明,两个品种果胶糖醛酸含量和果胶甲基酯化度存在差异,Hyogo品种根尖02.5 mm和2.55.0 mm根段果胶含量显著高于Alaska品种,02.5 mm段果胶甲基酯化度显著低于Alaska品种。品种Alaska根尖02.5 mm和2.55.0 mm根段因为具有较低果胶含量和较高果胶甲酯化度,所以表现出更低未甲酯化果胶含量,因此在铝毒条件下表现出低铝累积和低胼胝质含量为耐铝品种。这与对玉米和水稻等耐铝性/铝敏感性的研究结果一致[6]。Eticha等[19]发现不同耐铝性玉米品种根尖细胞壁果胶含量和果胶甲基酯化度存在明显差异,铝敏感品种中果胶含量高于耐铝品种,果胶甲酯化度低于耐铝品种,指出铝敏感型品种 Lixis 根尖中高果胶含量和低比例的甲基化果胶可能是其铝敏感的原因。
3.4PME和果胶甲基酯化度的关系
PME广泛存在于高等植物中,它的作用是水解果胶分子中甲酯化的羧基,通过脱甲基化来降低甲基酯化度(Pectin-COOCH3+2H2OPectin-COO++H3O+ +CH3OH)[20]。本研究结果表明,不同根段间PME活性存在显著差异,依次是02.5 mm>2.55.0 mm>5.010.0 mm,不同品种间也存在差异,Hyogo 根尖02.5 mm和2.55.0 mm根段均高于Alaska。因此Hyogo果胶甲基酯化度低于Alaska。这与Schmohl等[21]的研究结果一致,表明根尖细胞壁PME活性越高,果胶甲基酯化程度越低,说明根段或品种间果胶甲基酯化度的差异是PME活性不同引起的。
总之,根段或品种间耐铝性差异是由根尖未甲酯化果胶含量决定的,而未甲酯化果胶含量又受根尖细胞壁果胶含量和果胶甲酯化度影响。根尖(尤其是02.5 mm根段)低果胶含量和低果胶甲基酯化酶活性导致的高果胶甲基酯化度是豌豆不同耐铝品种的重要耐铝机制。本研究揭示果胶和PME在植物耐铝机制中的作用,为进一步揭示植物耐铝机理以及耐铝性状的遗传改良提供依据。
[1]Horst W J, Wang Y, Eticha D. The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: a review[J]. Annals of Botany, 2010, 106(1): 185-197.
[2]Ryan P R, Tyerman S D, Sasaki T,etal. The identification of aluminium-resistance genes provides opportunities for enhancing crop production on acid soils[J]. Journal of Experimental Botany, 2011, 62(1): 9-20.
[3]Delhaize E, Ryan P R, Randall P J. Aluminum tolerance in wheat (TriticumaestivumL.) II. Aluminum-stimulated excretion of malic acid from root apices[J]. Plant Physiology, 1993, 103(3): 695-702.
[4]Hoson T, Soga K, Wakabayashi K,etal. Growth and cell wall changes in rice roots during spaceflight[M]. Springer, 2003, 19-26.
[5]Ma J F. Syndrome of aluminum toxicity and diversity of aluminum resistance in higher plants[J]. International Review of Cytology, 2007, 264: 225-252.
[6]Schmohl N, Horst W J. Cell wall pectin content modulates aluminium sensitivity ofZeamays(L.) cells grown in suspension culture[J]. Plant, Cell & Environment, 2000, 23(7): 735-742.
[7]Yang J L, Li Y Y, Zhang Y J,etal. Cell wall polysaccharides are specifically involved in the exclusion of aluminum from the rice root apex[J]. Plant Physiology, 2008, 146(2): 602-611.
[8]Li Y Y, Yang J L, Zhang Y J,etal. Disorganized distribution of homogalacturonan epitopes in cell walls as one possible mechanism for aluminium-induced root growth inhibition in maize[J]. Annals of Botany, 2009, 104(2): 235-241.
[9]Yang J L, Zhu X F, Peng Y X,etal. Cell wall hemicellulose contributes significantly to aluminum adsorption and root growth in Arabidopsis[J]. Plant Physiology, 2011, 155(4): 1885-1892.
[10]Sivaguru M, Horst W J. The distal part of the transition zone is the most aluminum-sensitive apical root zone of maize[J]. Plant Physiology, 1998, 116(1): 155-163.
[11]Yu M, Ming Feng Y, Goldbach H E. Mist culture for mass harvesting of root border cells: aluminum effects[J]. Journal of Plant Nutrition and Soil Science, 2006, 169(5): 670-674.
[12]Köhle H, Jeblick W, Poten F,etal. Chitosan-elicited callose synthesis in soybean cells as a Ca2+-dependent process[J]. Plant Physiology, 1985, 77(3): 544-551.
[13]Jolie R P, Duvetter T, Houben K,etal. Carrot pectin methylesterase and is inhibitor from kiwi fruit: Study of activity, stability and inhibition[J]. Innovative Food Science & Emerging Technologies, 2009, 10(4): 601-609.
[14]Heim D R, Skomp J R, Waldron C,etal. Differential response to isoxaben of cellulose biosynthesis by wild-type and resistant strains ofArabidopsisthaliana[J]. Pesticide Biochemistry and Physiology, 1991, 39(2): 93-99.
[15]Blumenkrantz N, Asboe-Hansen G. New method for quantitative determination of uronic acids[J]. Analytical Biochemistry, 1973, 54(2): 484-489.
[16]Kobayashi Y, Yamamoto Y, Matsumoto H. Studies on the mechanism of aluminum tolerance in pea (PisumsativumL.) using aluminum-tolerant cultivar ‘Alaska’and aluminum-sensitive cultivar ‘Hyogo’[J]. Soil Science and Plant Nutrition, 2004, 50(2): 197-204.
[17]Wissemeier A H, Klotz F, Horst W J. Aluminium induced callose synthesis in roots of soybean (GlycinemaxL.)[J]. Journal of Plant Physiology, 1987, 129(5): 487-492.
[18]Horst W J, Schmohl N, Kollmeier M,etal. Does aluminium affect root growth of maize through interaction with the cell wall-plasma membrane-cytoskeleton continuum?[J]. Plant and Soil, 1999, 215(2): 163-174.
[19]Eticha D, Stass A, Horst W J. Cell-wall pectin and its degree of methylation in the maize root-apex: significance for genotypic differences in aluminum resistance[J]. Plant, Cell & Environment, 2005, 28(11): 1410-1420.
[20]Grsic-Rausch S, Rausch T. A coupled spectrophotometric enzyme assay for the determination of pectin methylesterase activity and its inhibition by proteinaceous inhibitors[J]. Analytical Biochemistry, 2004, 333(1): 14-18.
[21]Schmohl N, Pilling J, Fisahn J,etal. Pectin methylesterase modulates aluminium sensitivity inZeamaysandSolanumtuberosum[J]. Physiologia Plantarum, 2000, 109(4): 419-427.
Differences of pectin content and methylation in root cell wall of pea(Pisumsativum) cultivars with different Al tolerance
LI Xue-wen1, 2, LI Ya-lin1, 2, YANG Jin1, 2, WU Li-shu2, XIAO Hong-dong1, FENG Ying-ming1, LIU Jia-you1, YU Min1*
(1DepartmentofHorticulture,FoshanUniversity,Foshan,Guangdong528000,China;2CollegeofResourceandEnvironment,HuazhongAgriculturalUniversity,Wuhan430070,China)
【Objectives】 The study investigated the relationship between changes of cell wall pectin, pectin methyl esterification degree and aluminum tolerance in different cultivars of pea (Pisumsativum) to provide theoretical evidences how genetic differences affected aluminum tolerance during plant growth. 【Methods】 All cultivars were cultivated by mist under 24℃ growth chamber for 24 h after seed soaking. The root was divided into three segments: 0-2.5 mm, 2.5-5.0 mm and 5.0-10.0 mm. Pectin content, methyl esterification degree and the activity of pectin methylesterase (PME) were measured in different root segments of cultivar Hyogo and cultivar Alaska. Relative elongation rate (RER), callose and Al content were also tested to compare Al tolerance in different root zones and cultivars of pea. 【Results】 The RER of Alaska cultivar were significantly higher than Hyogo under Al concentration of 15 μmol/L and 30 μmol/L. And, the callose and Al of 0-5 mm or 5-10 mm segments in Alaska cultivar were accumulated much less than Hyogo. This result confirmed that the Alaska was an Al resistant cultivar and Hyogo was an Al sensitive cultivar. Alaska showed lower Al contents in all segments. In the meantime, the different Al levels between two species were showed significantly in 2.5-5.0 mm and 5.0-10.0 mm segments. Uronic acid content in cell wall pectin was found decreasing sequentially from segment 0-2.5 mm to 5.0-10.0 mm in both pea species. Alaska was measured lower cell wall pectin and un-methylated pectin compared with Hyogo, especially in segment 0-2.5 mm. The activity of PME also decreased successively from segment 0-2.5 mm to 5.0-10.0 mm in both Alaska and Hyogo. The degree of pectin methylation was significantly higher in 0-2.5 mm root segment than the other root segments in Alaska. Alaska had relatively lower content of uronic acid, lower activity of PME, and higher degree of pectin methylation, especially in 0-2.5 mm root segment. 【Conclusions】The key factor for pea cultivars shows Al tolerance or Al sensitive could be pectin content and methylation level in 0-2.5 mm root segment. Hyogo is highly sensitive to Al, which contained higher pectin in 0-2.5 mm root segment. The relatively lower pectin content and high methylation, especially in 0-2.5 mm, of Alaska associated with Al tolerance characterize.
pea; Al tolerance; cell wall pectin; degree of pectin methylation; pectin methylesterase
2014-11-25接受日期: 2015-07-20网络出版日期: 2015-12-08
国家自然科学基金资助项目(31172038);教育部科技研究重点资助项目(210154);广东省省级重大科研项目(2014KZDXM061)资助。
李学文(1987—),男,山东临沂人,博士研究生,主要从事植物营养生理研究。
Tel: 0757-85501986, E-mail: xuewen_1024@163.com。*通信作者 Tel: 0757-85501986, E-mail: yumin@fosu.edu.cn
Q945.78
A
1008-505X(2016)03-0729-07
猜你喜欢
酿酒科技(2021年5期)2021-06-06
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
三农资讯半月报(2020年18期)2020-10-14
酿酒科技(2019年1期)2019-02-25
数码世界(2018年1期)2018-12-23
中成药(2018年6期)2018-07-11
浙江工业大学学报(2017年5期)2018-01-22
中国酿造(2017年10期)2017-11-17
酿酒科技(2017年9期)2017-09-20
广东农业科学(2017年5期)2017-08-29
