
微生物抗药性进化的分子生物学证据及进化树构建方法
2016-08-27 06:39:18马亢刘海滨
甘肃科技纵横 2016年3期
马亢,刘海滨
(商丘师范学院生命科学学院,河南商丘476000)
微生物抗药性进化的分子生物学证据及进化树构建方法
马亢,刘海滨
(商丘师范学院生命科学学院,河南商丘476000)
摘要:微生物的抗药性一直是困扰人类的重大问题,尤其是最近超级细菌的出现更是为人类敲响了警钟——对微生物的抗药性研究已经刻不容缓。本论述梳理了近年来微生物进化方面的分子生物学证据,并介绍了两种最常用建立进化树的生物信息学方法,从而更加直观地找出他们的进化关系。
关键词:微生物;抗药性;进化;进化树
DOI10.3969/j.issn.1672-6375.2016.03.024
我们通过对微生物抗药性进化的研究来减缓其进化速度和找到其进化的规律性,以便更好地筛选相关药物,为人们的生活和生产活动服务。近年来,分子生物学迅速发展,使我们在分子水平能够找到微生物进化的过程以及他们之间的进化关系,借助一些生物学专业软件我们可以绘制出进化树,从而更加直观的找出他们的进化关系。找出微生物抗药性的进化规律是防止微生物进一步在抗药性上进化的有利武器。
1 微生物抗药性进化的分子生物学证据
1.116S rRNA构建系统发育树
16S rRNA是原核核糖体30S小亚基的组成部分,存在于所有细菌的基因组中[1]。其可以作为测量各类生物进化的工具的原因有以下几点:(1)rRNA是生物存活不可缺少的,普遍存在于真核和原核生物中,在生物进化历程中其稳定性比较高;(2)在16S rRNA分子中,有稳定度不同的序列区域,适用于测量进化距离不同的各类生物亲缘关系;(3)相对分子质量适宜序列分析。
1.2微生物抗药性的分子机制
1.2.1钙调磷酸酶机制
钙调磷酸酶是真核细胞信号传导途径中的关键调控因子,对于菌株应对环境压力(抗生素作用)是必不可少的。细胞内有一种帮助其他蛋白质折叠并运送它们到达细胞内适当的位置的分子——分子伴侣热激蛋白90(heat shock protein 90,HSP90)。钙调磷酸酶发挥正常功能必须借助HSP90对其亚基进行正确折叠。HSP90的转录出现变异将导致细菌出现抗药性[2]。PKA可以将转录因子CRZ1磷酸化,使其无法进入细胞核,造成转录异常。同时PKA出现异常,使相关基因转录受阻形成抗药性[3]。
1.2.2转运蛋白机制
一些抗生素进入细菌体内必须借助转运蛋白的运输,才能发挥杀菌作用。一些运输蛋白(如ABC运输蛋白)可以将药剂从膜内层转移至外层而排出细胞体外;另外一些运输蛋白(如CYP51蛋白)基因与药物作用时易发生点突变,造成编码蛋白与药物亲和力下降,使抗生素作用效果大大降低,导致抗药性。
1.2.3rpsL基因机制
链霉素作用于核糖体30S小亚基,直接抑制蛋白质的合成。其直接的位点为核糖体上的S12和16S RNA结构域,染色体上编码核糖体蛋白S12的高度保守的rpsL基因的单碱基突变,干扰了蛋白的生物合成,从而使病原细菌具有高水平抗性且能稳定遗传。一些研究对rpsL和16S rRNA基因序列进行分析和正反互补验证检测,证明了rpsL基因的第43位碱基突变造成抗药性突变[4]。
抗性基因常与质粒、转座子关联,链霉素的大量使用在一定程度上加剧了选择的进程,且自然界中本身存在的链霉素抗性基因库,二者的共同作用造成了抗性基因的大量传播。
基因重组是人类进化的一种可能的方式,微生物能大量进行基因重组,原核生物和真核生物中普遍存在的转座作用是微生物发生抗药性进化的一种重要机制[5]。
2 微生物进化树的构建
分析一个完整的进化树需要以下几个步骤:(1)对目的多序列目标进行排列;(2)构建一个进化树;(3)对进化树可信度进行评估。
在系统发育树的构建中应用最为广泛的是PHYLIP和MEGA两款生物软件,通过这两种软件的使用,我们可以得到较为清楚的生物进化关系。
2.1利用PHYLIP3.65软件构建菌株的系统发育树[6]
具体过程见图1所示:
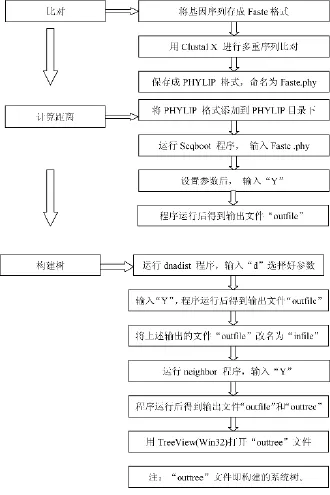
图1 PHYLIP3.65软件构建菌株系统发育树过程图
2.2利用MEGA3.1软件构建菌株的系统发育树[7]
具体过程见图2所示:
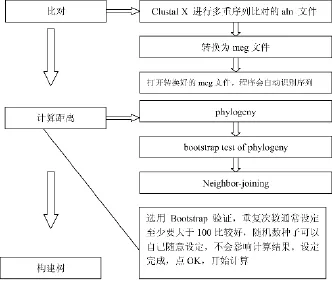
图2 MEGA3.1软件构建菌株系统发育树过程图
构建树的过程所耗时间和序列的数量和长短成正比。树枝上的数字表示bootstrap验证中该树枝可信度的百分比。
3 结束语
伴随着抗生素的大规模不当使用,微生物的抗药性逐年增加,这给人们的健康成本带来了极大的挑战。人们不断地筛选新的抗菌药物,但要从根本上解决抗药性的问题,我们必须了解微生物在抗药性方面的进化规律。微生物的抗药性和人类的抗生素筛选行为,可以看成是微生物为了适应人类制造的逆境(抗生素)而通过进化、突变等多种方式适应生存的结果,因此理顺微生物的进化模式,能够帮助我们找到更好的抗生素筛选策略,并能够从根本上研究解决微生物进化过快的问题。我们可以用分子生物学、基因组学、生物信息学的方法来研究其进化过程,找出规律,从而为进一步筛选药物以及合理使用药物提供科学依据。
参考文献:
[1]刘琳,刘洋,刘红娟.基于16S rDNA的系统发育分析在微生物进化关系中的应用[J].生物学通报,2008,43(11).
[2]Seung Oe Lim,Sung Gyoo Park.Expression of heat shock proteins(HSP27,HSP60,HSP70,HSP90,GRP78,GRP94)in hepatitis B virus-related hepatocellular carcinomas and dysplastic nodules[J].World Journal of Castroenerol 2002;11 (14):2072-2079.
[3]马宁,崔志峰.致病真菌抗药性机制的研究进展[J].中国药理学与毒理学杂志,2010,24(5).
[4]叶滔,马志强.植物病原真菌对甾醇生物合成抑制剂类(SBIs)杀菌剂的抗药性研究进展[J].农药学学报,2012,14 (1):1-16.
[5]Chiou CS,Jones AL.Molecular analysis of high-level streptomycin resistance in Erwinia amylovora[J].Phytopathology,1995,85:324-328.
[6]张光明,吴廷瑞.微生物与人类进化[J].医学与哲学,1999,20(7):20-22.
[7]陶士珩.生物信息学(普通高等教育十一五规划教材)[M].北京:科学出版社,2007.
中图分类号:Q939
文献标识码:A
收稿日期:2015-11-18
作者简介:马亢(1983-),男,回族,实验师,硕士,主要研究方向:生物化学与分子生物学。
猜你喜欢
天津市教科院学报(2021年5期)2021-11-10 07:32:40
生物学通报(2021年9期)2021-07-01 03:24:44
江苏农业科学(2016年8期)2017-02-15 19:54:11
出版广角(2016年14期)2016-12-13 02:16:10
道德与文明(2016年5期)2016-12-07 00:53:48
中小企业管理与科技·下旬刊(2016年11期)2016-11-18 08:19:35
中国科技博览(2016年19期)2016-10-19 11:47:05
科技视界(2016年9期)2016-04-26 12:23:48
科技视界(2016年7期)2016-04-01 09:39:11
淮阴工学院学报(2014年5期)2014-09-10 09:15:56
