
异钩藤碱通过mTOR非依赖性途径诱导SH-SY5Y细胞自噬发生
2016-08-25 09:45蔡雅卫王继业宁波市第二医院老年医学科浙江宁波500宁波市第二医院神经内科浙江宁波500浙江警察学院浙江杭州005
中国现代医生 2016年17期
徐 妍 严 旺 蔡雅卫 王继业.宁波市第二医院老年医学科,浙江宁波 500;2.宁波市第二医院神经内科,浙江宁波 500;.浙江警察学院,浙江杭州 005
异钩藤碱通过mTOR非依赖性途径诱导SH-SY5Y细胞自噬发生
徐妍1严旺2▲蔡雅卫1王继业3
1.宁波市第二医院老年医学科,浙江宁波315010;2.宁波市第二医院神经内科,浙江宁波315010;3.浙江警察学院,浙江杭州310053
目的 探讨异钩藤碱(Isorhynchophylline,IRN)人胶质瘤细胞SH-SY5Y细胞自噬的发生及其与mTOR信号通路的关系。方法 用不同浓度IRN(6、12和24 μmol/L)处理SH-SY5Y细胞24 h,免疫印迹法检测自噬标志性蛋白LC3-Ⅱ的表达,免疫荧光法观察微管相关蛋白轻链3(LC3)荧光斑点标记的自噬体形成,利用溶酶体稳定剂氯喹(CQ)以及自噬抑制剂3-MA进一步明确IRN作用的途径。Western蛋白印迹法检测mTOR相关蛋白和Beclin1的表达。结果IRN作用SH-SY5Y细胞24 h后,可显著上调LC3-Ⅱ表达,0、6、12、24 μM IRN作用后的相对表达量分别为(0.086±0.02)、(0.39±0.14)、(0.96±0.09)、(1.93±0.14)。自噬相关蛋白LC3-Ⅱ/LC3-Ⅰ的比值增高(P<0.01),并呈浓度依赖性,SH-SY5Y细胞内GFP-LC3荧光斑点标记的自噬体增多,在IRN不同浓度处理组(0、6、12、24 μM)中分别为2±1/细胞,4±1/细胞,8±1/细胞,和14±2/细胞。IRN联合CQ后可以促进LC3-Ⅱ的表达 (0.64±0.052)。IRN可以削弱a-Syn蛋白表达,在0、6、12、24 μM组中的相对表达量为 (0.71±0.07)、(0.69± 0.05)、(0.53±0.04)、(0.36±0.034),并呈浓度依赖性,联合3-MA后a-Syn表达上调(0.63±0.04),提示3-MA可以逆转由IRN对a-Syn的抑制效果,而CQ对a-Syn表达,与对照组比无显著影响。信号通路研究提示,对mTOR以及Beclin1表达不影响,Beclin1特异性siRNA作用后可以抑制IRN诱导的LC3-Ⅱ表达。 结论IRN可诱导SHSY5Y细胞自噬的发生,其机制为mTOR非依赖性自噬途径,Beclin1的表达可以影响IRN诱导的自噬。
异钩藤碱;SH-SY5Y细胞;自噬;Beclin1
[Abstract]Objective To explore the relationship between the occurrence of autophagy of human glioma cell SH-SY5Y induced by isorhynchophylline(IRN)and mTOR signaling pathway.Methods SH-SY5Y cells were treated by IRN with different concentrations(6,12 and 24 μmol/L)for 24h,and Western Blot method was used to test the expression of marker protein of autophagy LC3-Ⅱ.Immumofluorescence method was used to observe the formation of autophagosome marked by fluorescent spots in microtubule-associated protein LC3.Lysosome stabilizer CQ,and autophagy inhibitor 3-MA were used to further identify the action pathway of IRN.Western Blot method was used to test the expression of mTOR-associated proteins and Beclin1.Results After SH-SY5Y cells were treated by IRN,the expression of LC3-Ⅱwas significantly up-regulated.The relative expression amount after the action of 0,6,12 and 24 μM IRN was(0.086± 0.02),(0.39±0.14),(0.96±0.09),and(1.93±0.14)respectively.The ratio of autophagy-associated proteins LC3-Ⅱ/LC3-Ⅰwas increased(P<0.01),showing concentration dependence.Autophagosome marked by GFP-LC3 fluorescent spots were increased in SH-SY5Y cells,and was 2±1/cell,4±1/cell,8±1/cell and 14±2/cell respectively in the IRN treatment groups with different concentrations(0,6,12,24 μM).IRN combined with CQ was able to promote the expression of LC3-II(0.64±0.052).IRN was able to reduce the expression of a-Syn protein.The relative expression amount in 0,6,12 and 24 μM groups were(0.71±0.07),(0.69±0.05),(0.53±0.04),and(0.36±0.034)respectively,showing concentration dependence.After the combination with 3-MA,a-Syn expression was up-regulated(0.63±0.04),indicating that 3-MA was able to reverse the inhibiting effects of IRN on a-Syn.The effects of CQ on the expression of a-Syn were not sig-nificant compared with the control group.The study on signaling pathway indicated that the expression of mTOR and Becline1 was not affected,and with the action of Beclin1-specific siRNA,the expression of LC3-Ⅱinduced by IRN could be inhibited.Conclusion IRN is able to induce the occurrence of autophagy of SH-WY5Y cells,and the mechanism is independent autophagy pathway of mTOR.The expression of Beclin1 is able to affect the autophagy induced by IRN.
[Key words]Isorhynchophylline;SH-SY5Y cells;Autophagy;Beclin1
细胞自噬是常见的生物学现象,是对细胞内环境稳定的高度保守的生理过程,通过降解胞浆内细胞器,长寿命蛋白和有聚集倾向的蛋白,进而满足细胞对物质的需求,一定意义上也是细胞在饥饿或者环境压力下的一种自然生理过程。有多种神经退行性疾病与细胞内某些蛋白的异常聚集有关,如阿尔茨海默病(Alzheimer disease,AD)、帕金森病(Parkinson Disease,PD)和亨廷顿病疾病属于神经变性疾病,这些疾病的发生在一定程度上与大脑受累区域中异常蛋白的过度积聚有关。比如像a-突触核蛋白(a-Syn)的过度表达与和家族型帕金森的发生密切相关[1]。a-Syn的点突变则可以增加其聚集倾向,并可导致家族性PD的早期发病,在小鼠的研究中同时也验证了过量表达的a-Syn可以导致脑神经中多巴胺能神经元的丧失,以及细胞质内包涵体的形成,并最终导致进行性运动缺陷,普遍认为a-Syn寡聚体的积累或包涵体的形成会对神经元细胞产生毒性,最终可以导致神经元细胞直接死亡。此外,在老年痴呆患者中往往有Tau蛋白的过量聚集,Tau蛋白的过度磷酸化与AD发病的相关性密切[2]。目前认为,a-Syn、Tau蛋白以及其他类似的可以引起神经退行性病变的寡聚蛋白极大程度上依赖于细胞自噬途径的清除,这些蛋白在细胞内的体积往往较大,无法通过蛋白酶体的狭窄核心进行降解。
异钩藤碱(Isorhynchophylline,IRN)是从茜草科中药钩藤抽取提的具有较强药理活性的羟吲哚生物碱,除已被临床应用于治疗心血管疾病外,近年来研究发现,其对中枢神经系统具有广泛的药理学作用,存在多种生物活性,比如对缺血诱导引起的神经元损伤的保护作用[3];抑制LPS诱导的星形胶质细胞炎症因子的释放[4];显著降低大脑皮层的兴奋性,具有明显的镇静、安眠、降压、解痉等作用,是安神养血口服液中的主要成分之一[5],这些研究都从不同角度说明IRN可以较好通过血脑屏障而影响脑内神经元细胞[6]。目前尚无IRN在自噬方面的研究。本项目在2014年12月~2015年12月开展,着重考察IRN对神经胶质瘤细胞,SH-SY5Y细胞自噬的影响,探讨IRN是否可诱导SH-SY5Y细胞自噬性以及相关机制分析。
1 材料与方法
1.1细胞、试剂和主要仪器
神经母细胞瘤SH-SY5Y细胞购自中国科学院上海细胞库。新生牛血清,DMEM培养基和0.25%胰蛋白酶购自美国Life technology公司;异钩藤碱购自美国Sigma公司;mTOR通路相关抗体,美国Santa Cruz Biotechnology公司;兔抗人自噬微观相关蛋白1轻链3(microtubule-associatedprotein1lightchain3,LC3)抗体,p62抗体,兔抗人Beclin1抗体,以及二抗均购自Abcam;mTOR以及P70S6K磷酸化抗体购自CST公司;底物化学发光试剂,Fluor Save抗荧光淬灭剂,3-MA(3-Methyladenine)以及氯喹(CQ)均购自美国Millipore公司;其余试剂均为国产分析纯。
1.2细胞培养和药物处理
SH-SY5Y细胞培养在含有10%胎牛血清以及100 U/mL青-链霉素霉素的DMEM培养液中,置于含有5%CO2,37℃培养箱中,每隔2~3天换液。取对数生长期细胞进行实验。将SH-SY5Y细胞以1×104/mL密度接种于6孔或12孔板,置于培养箱中培养24 h后,IRN配置成10 mM储液,使用时用完全培养稀释,加入到细胞中使得终浓度分别为0、6、12和24 μM,放回培养箱孵育12~72 h用于后续的实验。
1.3GFP-LC3斑点计数
细胞爬片后,经IRN处理的细胞用PBS漂洗一遍后,4%多聚甲醛固定15min,含0.3%TritonX-100 PBS透化10 min,羊血清封闭30 min后,加入LC3抗体(1:200),4℃孵育过夜,PBS漂洗后加入FITC标记的二抗(1∶50)室温下避光反应1h,PBS漂洗后用Fluor Save试剂封片,荧光显微镜计数细胞内斑点情况。随机选择视野中至少50个细胞用于计数,计算斑点总数/细胞总数。
1.4免疫印迹
经不同浓度IRN处理后,冷PBS漂洗一遍,RIPA裂解细胞,收集蛋白并用BCA法测定蛋白浓度。按20 μg/泳道蛋白量进行SDS-PAGE电泳,转膜并用脱脂牛奶封闭后,加入抗mTOR磷酸化抗体(1∶1000)、P70S6K(1∶1000)、LC3(1∶1000)、Beclin1(1∶1000),p62 (1∶2000)和β-actin(1∶5000)抗体于4℃孵育过夜。取出膜用TBS-T漂洗3次后,将膜和相应2抗(1∶5000)在摇床上室温孵育1 h,取出膜后TBS-T漂洗3次后,加入ECL化学发光试剂显色后,用LI-COR免疫印迹成像系统进行扫描,以ImageJ软件分析各条条带积分吸光度值,以β-actin条带为内参,将目标条带吸光度数值除以β-actin条带吸光度数值后得到目标条带的相对表达量,所有免疫印迹实验数值均为3次独立实验后得到的均值,并表示为平均光密度值±SD值形式用于半定量分析。
1.5统计学分析
应用SPSS19.0统计学软件进行分析。实验结果数据为3次独立实验均值,计量结果用均数±标准差(x±s)表示,符合正态分布,则组间比较根据数据类型采用独立样本t检验,多组间比较用ANOVA方差分析;若不符合正态分布,则采用非参数 Mann-Whitney检验。P<0.05为差异具有统计学意义。
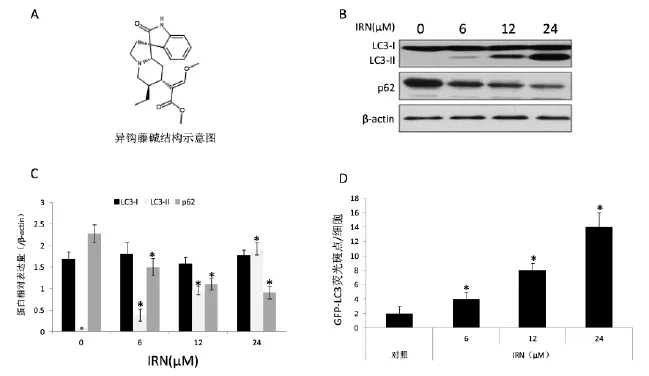
图1 IRN诱导SH-SY5Y细胞自噬的发生
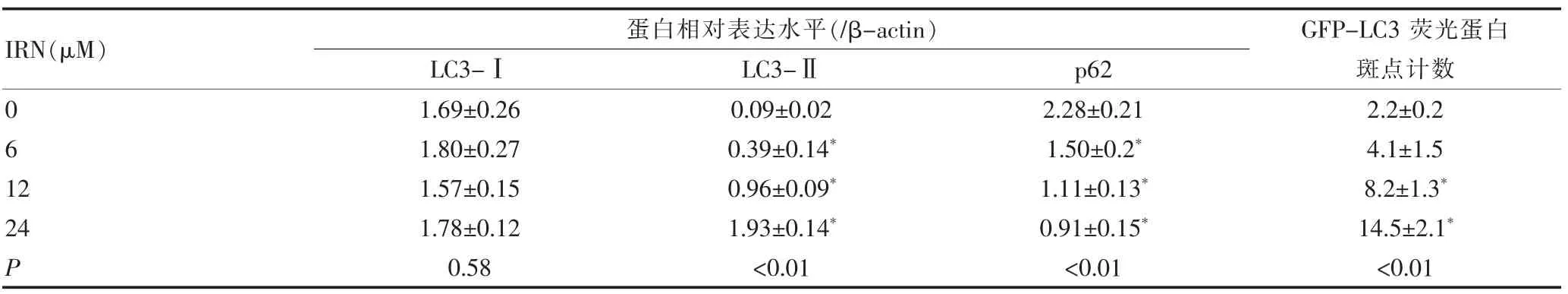
表1 不同浓度IRN对自噬相关蛋白相对表达水平的影响(x±s,n=3)
2 结果
2.1异钩藤碱引起SH-SY5Y细胞自噬相关蛋白表达
LC3-Ⅱ是自噬特异性标志物,其表达水平一定程度上反映细胞自噬体情况。如图1所示,不同浓度IRN作用SH-SY5Y细胞后可以上调LC3蛋白的表达,并且有剂量依赖性,即随着IRN浓度增加,表达水平也逐渐上调,对条带进行量化后,其数值在0、6、12、24 μM组中,LC3-Ⅰ的相对β-actin表达分别为(1.69±0.16)、(1.80±0.27)、(1.57±0.15)、(1.78±0.12),LC3-Ⅱ的表达分别为(0.086±0.02)、(0.39±0.14),(0.96±0.09)、(1.93±0.14)。而p62的表达水平则逐渐下调,其数值在0、6、12、24 μM组中分别为(2.28± 0.21)、(1.50±0.20)、(1.11±0.13)、(0.91±0.15)。提示IRN可以诱导细胞自噬发生。随后我们通过建立恒定表达GFP-LC3的细胞株,用来观察IRN处理以后GFPLC3斑点的形成情况,如图1C所示IRN作用24 h后,可以显著诱导大量GFP-LC3斑点的形成,GFPLC3荧光斑点数目在对照和IRN不同浓度处理组(6、12、24 μM)中分别为2±1/细胞,4±1/细胞,8±1/细胞,和14±2/细胞,IRN各浓度组与对照组比,差异具有统计学意义(n=3,P<0.05)。以上结果都证实IRN是可以引起神经元细胞SH-SY5Y自噬发生的诱导物质。
2.2异钩藤碱对自噬体成熟的影响
为了解IRN对自噬的诱导机制,我们进一步通过加入溶酶体抑制剂CQ来判断其是否可以影响自噬溶酶体成熟,如图2所示,通过CQ和IRN处理12 h后,对细胞内蛋白进行WB分析提示,CQ作用后可以显著的提高LC3的表达,LC3-Ⅰ表达水平从对照组的(0.92±0.0011)上调到(0.68±0.02),LC3-Ⅱ从对照组中的(0.11±0.03)上调到(0.29±0.04)。在CQ加入IRN后可以进一步提高LC3-Ⅰ(0.76±0.03)和LC3-Ⅱ(0.64±0.052)的表达,要显著高于单用CQ组(0.68± 0.02)的水平(n=3,P<0.05)。此外我们还分析了GFPLC3的斑点实验,结果发现IRN组中为14±2/细胞,而用5mM的3MA处理24 h后,可削弱IRN对GFP-LC3斑点的诱导效应,GFP-LC3斑点为3±1/细胞(n=3,P<0.05)。在单用CQ和CQ+IRN组中,GFP-LC3为21±2/细胞,和30±3/细胞,提示CQ刺激LC3表达,和IRN合用后,效果增强。该进一步证实,IRN是SH-SY5Y细胞的自噬诱导物,并与CQ有协同促进LC3-Ⅱ表达的作用,其诱导的自噬可以被3-MA阻断。
2.3异钩藤碱对a-Syn蛋白聚集的影响
为进一步了解IRN对当代聚集的影响,考察对a-Syn蛋白表达的影响。如图3A所示,随着IRN浓度的增加,a-Syn的表达水平逐渐下调,并呈现浓度依赖性,在0、6、12、24 μM组中的相对表达量为:(0.71± 0.07),(0.69±0.05),(0.53±0.04),(0.36±0.034)。随后对我们加入CQ和3-MA,结果显示3-MA可以上调a-Syn的表达(0.64±0.04),这也就提示,在神经元细胞中,抑制自噬可能会抑制积聚蛋白的降解。而在3-MA+IRN组中,可以上调a-Syn表达(0.63±0.04),提示3-MA对由IRN引起的a-Syn积聚有一定的削弱作用。而在CQ处理组中,a-Syn的表达有一定程度的上调(0.42±0.04),同时加入IRN和CQ后以后对a-Syn的表达并无显著影响(0.36±0.032,与单用CQ组比,n=3,P>0.05)。该结果提示3-MA对a-Syn积聚的影响较大,自噬是清理a-Syn的重要途径,IRN可以减少a-Syn的表达。
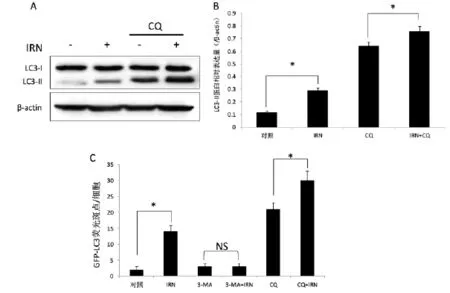
图2 IRN联合溶酶体稳定剂以及自噬抑制剂后对LC3表达的影响

表2 加入自噬抑制剂后对细胞内GFP-LC3荧光斑点数目的影响

图3 IRN对a-Syn蛋白表达的影响
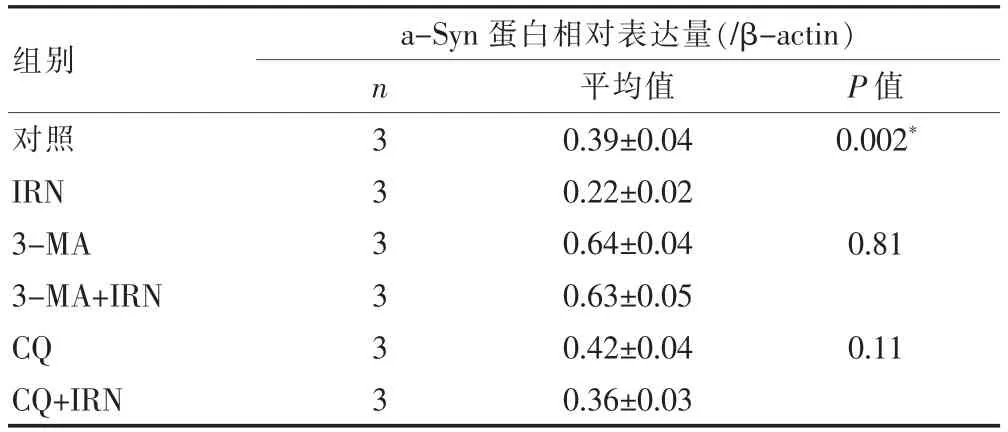
表3 IRN加入不同自噬抑制剂后对a-Syn蛋白表达水平的影响
2.4异钩藤碱诱导自噬的途径分析
为了进一步阐明IRN作用的分子机制。我们对自噬经典途径mTOR进行分析,结果如图4A所示,磷酸化mTOR以及其底物p70s6k都不受IRN的影响,而阳性药物雷帕霉素加入后可以大幅度抑制mTOR和pP70s6k磷酸化。此外,我们还检测在IRN处理后,其他的几个途径包括AKT、AMPK、MEK/ERK、JNK和ER应激途径,以及钙信号途径等,都未见有显著改变(数据未展示)。最后我们检测了Beclin1蛋白,该蛋白是自噬激活中的关键因子,上调Beclin1后可以诱导自噬的发生,结果如图4所示,IRN并不影响Beclin1蛋白的表达,但是用Beclin1特异性是siRNA干扰其表达以后可以显著抑制由IRN诱导的LC3-Ⅱ的表达,这一结果也提示,Beclin1蛋白在IRN诱导自噬中的作用也是非常关键的,提示IRN直接作用的蛋白可能需要Beclin1蛋白的协助形成自噬激活物后发挥作用。

图4 IRN对mTOR磷酸化以及Beclin1表达的影响
3 讨论
细胞自噬过程中,常常伴随有LC3蛋白的表达,LC3即Light chain3,最初被认为是微管相关蛋白1A 和1B即MAP1LC4,随后在酵母中发现,和该蛋白高度同源的Atg8/Aut7/Cvt5对于自噬至关重要,在人体中LC3有3种亚型,LC3A、LC3B和LC3C。在自噬开始前以无活性的形式存在,LC3通过羧基端即C端与自噬相关的特异性基因4(Atg4)结合,剪切为LC3-Ⅰ在胞浆内表达,随后通过泛素化系统与脂酰乙醇胺(PE)发生酯化反应,得到LC3-Ⅱ(LC3-PE),这其中有Atg7和Atg3的参与。LC3-Ⅱ分子可以特异地结合在自噬膜泡的内膜和外膜上,LC3-Ⅱ的含量与自噬泡数量呈正相关,从而可以用于评估自噬活性[7]。因此,其常作为研究细胞自噬的标志分子。本研究结果显示,不同浓度IRN处理SH-SY5Y细胞24h可浓度依赖性地上调LC3-Ⅱ表达,自噬相关蛋白LC3-Ⅱ/LC3-Ⅰ的比值增高,IRN 6~24 μmol·L-1处理SH-SY5Y细胞24 h后,GFP-LC3荧光斑点标记的自噬体增多,提示自噬体大量形成。这些结果都证明,IRN可能通过激活自噬相关蛋白LC3导致SH-SY5Y细胞发生自噬。目前,有关自噬在神经退行性疾病的发生发展中的作用报道不一[8,9],在亨廷顿病研究中发现,依赖神经元细胞自噬途径可加速突变亨廷顿蛋白清除,减少蓄积,同时诱导细胞自噬的发生可减缓神经的退行性变[10,11]。在阿尔茨海默病中的研究发现,神经元细胞中的自噬体富含Aβ和APP羧基末端偏殿,以及早老素依赖性的γ-分泌酶,抑制神经元自噬将有助于减少Aβ40和Aβ42额分泌,而诱导自噬则可以促进前述2种蛋白的分泌。但是从另一种角度,自噬也是清除Aβ的一种途径,较多的细胞和动物实验认为,诱导自噬可以减少Aβ的聚集。因此自噬在AD的发病机制中的作用存在争议[12,13]。在海藻酸钠诱导大鼠纹状体神经元兴奋性损伤研究中,自噬的激活可促进神经元死亡,并可被自噬抑制剂所削弱。以上研究提示,细胞自噬的发生在神经系统疾病中既有保护作用又可能有损伤作用,和具体的疾病和神经元有关。有研究显示,IRN可以保护PC12细胞对抗由β-淀粉样蛋白引起的细胞凋亡[14],在多项动物实验中显示,IRN可以改善由β-淀粉样蛋白引起的大鼠认知损伤,主要与抑制神经元细胞凋亡和tau蛋白形成有关[15],IRN可以改善D-半乳糖引起的小鼠学习和记忆损伤[16],这些研究都从不同角度证实了IRN在神经元保护中的作用和价值,结合本研究得到的结果,我们认为IRN在神经元退行性疾病中可能主要以保护作用为主。
多巴胺能神经元是PD患者大脑中受影响主要细胞,a-Syn在期中枢神经细胞中的过表达可以导致多种生物体多巴胺能神经元的变性。a-Syn降解主要通过蛋白酶体、自噬以及伴侣分子介导的自噬即CMA等途径,但是仅有2个自噬途径可以降解a-Syn。具体说,已经有报道,a-Syn可以抑制cma并其只有巨自噬才能降解该蛋白。自噬也被认为可用于对抗突触核蛋白病的治疗策略,本研究中结果提示IRN可以显著削弱a-Syn的表达,后续研究将进一步考察IRN 对a-Syn突变体以及a-Syn/Synphilin-1聚集体的影响,这些结果在既往针对雷帕霉素、海藻糖以及17-AAG中的研究中未见报道。Beclin1是酵母自噬相关的特异性基因6也就是Atg6在人体中的同源体,主要可表达在高尔基体、线粒体、内质网以及细胞浆内[17],参与调控自噬前体生成和自噬体的形成。因此,Beclin1表达增强也可作为评价自噬水平升高的重要指标。尽管IRN作用后不影响Beclin1表达,但是siRNA干扰Beclin1后可以显著削弱IRN诱导的LC3-Ⅱ上调,提示Beclin1参与IRN诱导的自噬,后者可能会影响到自噬体其他相关蛋白表达。本研究组将在后续实验中进一步明确IRN对Beclin1相关蛋白的影响。以期进一步证实自噬在IRN诱导SH-SY5Y细胞自噬中的作用及与Beclin1的关系。
细胞自噬性死亡的发生受多种因子的调控,主要可以分为mTOR依赖性通路和mTOR非依赖性通路[18],mTOR作为一个丝苏氨酸蛋白激酶,能够感知细胞内ATP、生长因子、胰岛素的水平,进而也就是感受细胞内营养和能量的变化,参与组成TORC1和TORC2,并可以受上游多种不同信号通路的影响,比如经典的PI3K、MAPK等通路,影响自噬的发生[19]。mTOR的活化和抑制主要受到TSC1/TSC2的控制。而mTOR非依赖性通路则主要包括RAS-MAPK、Ca-AMPK、p53、PTEN以及内质网应激等[20]。同时我们的研究也提示,IRN通过mTOR非依赖性途径诱导细胞自噬,并排除了其他几种mTOR非依赖性通路的可能,但是未对p53以及PTEN等通路进行检测,后续研究有必要进一步分析其作用的具体蛋白,IRN通过mTOR非依赖性通路发挥作用,也在一定程度上消除了未来临床应用中与mTOR途径相关的副作用和并发症发生的可能性。
本研究结果表明,IRN可诱导SH-SY5Y细胞发生自噬性死亡,溶酶体稳定后可以促进IRN诱导的LC3-Ⅱ表达,自噬抑制剂3-MA作用后则削弱其表达,分子途径分析认为,IRN诱导的自噬为mTOR非依赖型,对Beclin1蛋白的表达无影响,可能通过活化与Beclin1相关蛋白,与Beclin1结合后活化自噬有关。综上所述,IRN作为一种可以通过血脑屏障的传统中药钩藤的活性组分,不仅能够减少异常蛋白的积聚,而且有潜在的保护作用,IRN有进一步研究的价值,并有望将结果应用于临床。
[1]Lynch-Day MA,Mao K,Wang K,et al.The role of autophagy in Parkinson's disease[J].Cold Spring Harb Perspect Med,2012,2(4):a009357.
[2]宋明洁,戴雪伶,姜招峰.tau蛋白过磷酸化与阿尔茨海默病[J].生命科学,2013,(3):315-319.
[3]罗小金.异钩藤碱对阿尔茨海默病及其他中枢神经系统疾病的药理作用研究进展[J].医学研究生学报,2015,28(10):1112-1115.
[4]宋宇,刘继平,石富国,等.异钩藤碱对脂多糖诱导的星形胶质细胞炎性介质释放的抑制作用[J].药学与临床研究,2011,19(4):311-314.
[5]王捷,龙禹,焦爱军,等.HPLC测定安神养血口服液中3种生物碱延胡索乙素、钩藤碱和异钩藤碱含量[J].中成药,2010,32(2):231-235.
[6]汪江涛,丁伯平.钩藤碱和异钩藤碱药理作用研究进展[J].中国医药导刊,2012,14(7):1224-1225.
[7]倪志华,张玉明,邓传怀,等.LC3基因在细胞自噬过程中的表达研究[J].湖北农业科学,2015,54(20):4932-4936.
[8]姜凌,常诚.自噬与神经退行性疾病[J].卒中与神经疾病,2014,21(2):125-128.
[9]郑晨曦,张建平,姚伟伟,等.线粒体自噬与神经退行性疾病[J].中华神经外科疾病研究杂志,2015,(3):286-288.
[10]孙鑫,王进,秦正红,等.自噬及其在亨廷顿舞蹈病发病机制中的作用[J].中国神经免疫学和神经病学杂志,2011,18(1):54-57.
[11]蒋小岗,邬君超,林芳,等.靶向自噬途径作为亨廷顿舞蹈病的潜在治疗策略[J].中国药理学与毒理学杂志,2011,25(2):220-222.
[12]金贺,王蓉.阿尔茨海默病:自噬与β淀粉样肽关系的研究进展[J].中国比较医学杂志,2015,(8):68-71,75.
[13]史新秀.自噬在阿尔茨海默病发病中的作用及研究进展[J].中风与神经疾病杂志,2014,31(9):850-853.
[14]Xian YF,Lin ZX,Mao QQ,et al.Isorhynchophylline protects PC12 cells against Beta-amyloid-induced apoptosis via PI3K/Akt signaling pathway[J].Evid Based Complement Alternat Med,2013,2013:163057.
[15]Xian YF,Mao QQ,Wu JC,et al.Isorhynchophylline treatment improves the amyloid-β-induced cognitive impairment in rats via inhibition of neuronal apoptosis and tau protein hyperphosphorylation[J].J Alzheimers Dis,2014,39(2):331-346.
[16]Xian YF,Su ZR,Chen JN,et al.Isorhynchophylline improves learning and memory impairments induced by D-galactose in mice[J].Neurochem Int,2014,76:42-49.
[17]Menzies FM,Fleming A,Rubinsztein DC.Compromised autophagy andneurodegenerativediseases[J].NatRevNeurosci,2015,16(6):345-357.
[18]李治国,张继红,张锦华.mTOR通路对神经母细胞瘤SH-SY5 Y细胞自噬作用的影响[J].现代肿瘤医学,2014,(10):2278-2280.
[19]Kim YC,Guan KL.mTOR:A pharmacologic target for autophagy regulation[J].J Clin Invest,2015,125(1):25-32.
[20]Steelman LS,Chappell WH,Abrams SL,et al.Roles of the Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity to therapy-implications for cancer and aging[J].Aging(Albany NY),2011,3(3):192-222.
Occurrence of autophagy of SH-SY5Y cells induced by isorhynchophylline via independent pathway of mTOR
XU Yan1YAN Wang2CAI Yawei1WANG Jiye3
1.Department of Geriatrics,Ningbo Second Hospital,Ningbo 315010,China;2.Department of Neurology,Ningbo Second Hospital,Ningbo315010,China;3.Zhejiang Police College,Hangzhou 310053,China
R742.5
A
1673-9701(2016)17-0028-07
浙江省医药卫生科技计划基金项目支持(2014 KYA198)
▲
(2016-03-31)
猜你喜欢
新作文·小学低年级版(2022年3期)2022-08-30
农技服务(2021年7期)2021-09-24
智慧少年·故事叮当(2021年5期)2021-08-23
数学年刊A辑(中文版)(2018年2期)2019-01-08
今日农业(2019年15期)2019-01-03
商情(2017年38期)2017-11-28
中国洗涤用品工业(2017年2期)2017-04-16
中国医疗美容(2015年1期)2015-07-12
小星星·阅读100分(高年级)(2014年10期)2014-10-29
中国民族民间医药(2012年18期)2012-11-20
