
利用钾吸收基因表达评价葡萄叶面喷施钾肥效果和喷施浓度
2016-08-24 09:10:01贾海锋纠松涛王梦琦
植物营养与肥料学报 2016年4期
张 成, 贾海锋, 王 剑, 纠松涛, 王梦琦
(南京农业大学园艺学院, 江苏南京 210095)
利用钾吸收基因表达评价葡萄叶面喷施钾肥效果和喷施浓度
张 成, 贾海锋*, 王 剑, 纠松涛, 王梦琦
(南京农业大学园艺学院, 江苏南京 210095)
【目的】叶面喷施钾肥可以快速、 高效地为葡萄补充钾营养,促进葡萄的高产和优质,已经被广泛应用于葡萄生产中。本实验通过对葡萄叶面喷施不同种类及浓度钾肥,测定葡萄叶片和果实等生理指标变化,以及钾吸收相关基因的表达变化,从生理和基因水平上评价这些钾肥的喷施效果,为葡萄生产中钾肥的施用提供一定指导。【方法】本实验以‘夏黑’葡萄为试材,选择两个葡萄生长关键时期盛花期和果实膨大期分别对葡萄叶片喷施0.2%、 0.5%和0.8%三种浓度的K2SO4、 K2CO3、 K2SO4·2MgSO4和KCl。然后对葡萄叶片和新梢的生长率,坐果率,叶绿素含量,单粒重,可溶性固形物等生理指标进行统计分析,并利用荧光定量PCR技术分析4个钾吸收相关基因VvHAK13、VvKEA2、VvSIRK和VvSORK的表达情况。【结果】叶面喷施4种钾肥后,葡萄叶片和新梢的生长率,坐果率,叶绿素含量,单粒重,可溶性固形物等各项生理指标均有不同程度的提升。钾肥种类不同,最适喷施浓度不同,同种钾肥在葡萄盛花期和果实膨大期的最适喷施浓度也有所不同,四种钾肥在果实膨大期的最适喷施浓度普遍高于盛花期。四个钾吸收相关基因在喷施不同种类及浓度钾肥后也表现出不同的表达模式,总体来讲VvKEA2、VvSIRK、VvSORK的表达上调,而VvHAK13的表达下调。果实膨大期,需喷施较高浓度的钾肥,钾吸收相关基因才表现出较强烈的响应,而在盛花期则只需喷施较低浓度的钾肥。综合生理指标和基因表达两方面结果,得出4种钾肥效果依次为K2SO4·2MgSO4>K2SO4>K2CO3>KCl。盛花期K2SO4和K2CO3的最适喷施浓度为0.5%,K2SO4·2MgSO4和KCl的最适喷施浓度为0.2%; 果实膨大期K2SO4、 K2SO4·2MgSO4和KCl的最适喷施浓度为0.5%,K2CO3的最适喷施浓度为0.8%。【结论】葡萄叶面喷施钾肥可以有效促进葡萄叶片和果实的生长发育,四种钾肥的效果依此为: K2SO4·2MgSO4>K2SO4>K2CO3>KCl。盛花期K2SO4和K2CO3的最适喷施浓度为0.5%,K2SO4·2MgSO4和KCl的最适喷施浓度为0.2%; 果实膨大期K2SO4、 K2SO4·2MgSO4和KCl的最适喷施浓度为0.5%,K2CO3的最适喷施浓度为0.8%。适宜的喷施浓度可以有效提高钾吸收相关基因的表达,是其提高钾吸收利用的机理之一。
葡萄; 叶面喷肥; 钾肥; 钾吸收基因; 基因表达
作为喜钾植物,葡萄对钾营养的需求巨大,叶面喷肥钾肥具有技术简单、 用量少、 见效快、 利用率高等优点,在葡萄生产中逐渐被广泛应用[1-3]。钾素在提高葡萄抗逆性、 抗病性,促进果实发育,提高产量,增进品质等方面作用显著[4-7],如Alt1ndili等[8]报道葡萄喷施1%2%的KNO3可以显著提高葡萄的产量和质量,郑秋玲等[9]研究表明叶面喷施KH2PO4能使赤霞珠葡萄枝条中可溶性糖及淀粉的含量分别提高74.7%和27.7%。近年来,葡萄全基因的测序为基因信息应用于农业生产提供了全新的思路和条件[10-12],初建青等利用葡萄N代谢相关基因的表达信息判断施肥效果[13-14],王晨等利用葡萄花果发育相关基因的表达信息描绘了葡萄物候期,并用来指导葡萄精确施肥[15]。葡萄生长发育不仅受营养状况影响,还受各种外界环境的影响,基因信息可以精准反映植物营养吸收、 转运、 代谢等生理状态。通过分析葡萄叶面喷施钾肥后其叶、 花、 果等生理指标的变化以及钾吸收、 转运相关基因的表达,我们可以从宏观和微观两个层面全面而精准地评价不同钾肥的作用效果,并筛选出适合葡萄叶面喷施的钾肥种类及浓度。本文拟通过葡萄叶面喷施钾肥实验,从葡萄新梢生长、 叶片生长、 叶绿素含量、 坐果率、 果实单粒重、 可溶性固形物等生理指标,以及4个钾吸收相关基因: 钾离子转运体13(probable K+transporter 13,HAK13)、 钾离子反向转运体2(K+efflux antiporter 2,KEA2)、 shaker家族内向整流钾离子通道(inward rectifying shaker-like K+channel,SIRK)、 shaker家族外向整流钾离子通道(shaker-like potassium channel,SORK)的表达量两方面综合分析,评价不同种类钾肥及其浓度的作用效果,以期为葡萄生产中钾肥的施用提供一定参考。
1 材料与方法
1.1材料来源
本研究所用实验材料为5年生‘夏黑’(V.vinifera×V.labruscaSumerblack)葡萄,由江苏省农博园提供。 K2SO4、 K2CO3、 KCl购自南京化学试剂有限公司,K2SO4·2MgSO4购自青海中信国安科技发展有限公司。反转录试剂盒购自南京基天生物技术有限公司。荧光定量染料SYBR GreenⅠ购自上海浦迪生物科技有限公司。所用引物由北京鼎国昌盛生物技术有限责任公司合成(表1)。
1.2试验设计
试验采用两个喷施时期和三个喷施浓度,以喷清水为对照。喷施时期为盛花期(2014年5月25日)和果实膨大期(2014年7月16日),三个喷施浓度为0.2%、 0.5%、 0.8%的K2SO4、 K2CO3、 K2SO4·2MgSO4和KCl溶液(质量比)。选长度和粗度基本一致的葡萄植株,于上午7点对葡萄叶面和叶背分别喷施钾肥,喷施量以叶面滴水为度。每个处理3棵葡萄,分别于喷施后0、 6、 12、 24和48 h采第15个节位长枝条上的第9、 10节位的成熟叶片,液氮处理后-40℃保存备用。
1.3测定项目与方法
1.3.1 生理指标测定每个处理随机选取10个枝条,分别于处理前和处理10天后测量新梢长度和成熟叶的长与宽,计算新梢生长率和叶片大小增长率。盛花期喷施钾肥后,每个处理随机选取10个花序,统计花蕾数,待坐果后统计坐果数,计算坐果率。坐果率(%)=(坐果数/花蕾数)×100。
喷施处理10天后采第15个节位长枝条上的第9、 10节位的成熟叶片测定叶绿素含量。测定和计算方法参考郝再彬等[16]和叶济宁[17]的方法。
果实成熟时(8月10日),每处理随机选取10穗葡萄,用电子天平(0.01g)称量单果重,并用可溶性固形物含量浓度检测仪(LH-T32,杭州陆恒生物)测量葡萄可溶性固形物含量。
测得的各项生理数据采用SPSS进行方差分析。
1.3.2 RNA的提取与CDNA的合成‘夏黑’葡萄叶片总RNA的提取、 消化参照张彦苹等[18]和Chang等[19]的方法。mRNA的纯化采用Promega公司生产的PloyATtract® mRNA Isolation System IV试剂盒进行。cDNA合成以mRNA为模板,引物P01反转录合成cDNA第一条链,引物P02延伸加帽子,空气加热条件下42℃保温1 h,75℃保温10 min,冰上冷却2min后,-70℃保存备用。
1.3.3 荧光定量PCR应用Bio-Rad My-IQ 2荧光定量PCR仪进行实时定量。反应体系按SYBR GreenⅠ(TOYOBO)说明书进行。以葡萄看家基因UBI作为内参。扩增体系含1 μL cDNA,上下游引物各0.8 μL(引物信息详见表1),10 μL反应MIX,7.4 μLddH2O,总体系20μL。反应程序为95℃变性1 min,95℃变性10 s,Tm退火20 s,72℃延伸30 s,40个循环; 反应结束后分析荧光值变化曲线以及融解曲线。试验设3次重复,试验数据用LinRegPCR[21]和Excel软件分析,采用2-△△CT计算方法,以0 h为对照进行相对表达量分析。

表1 荧光定量PCR引物
2 结果与分析
2.1叶面喷施不同浓度钾肥对坐果率的影响
钾肥可以促进葡萄花果发育,对葡萄坐果率有重要影响。本实验结果显示叶面喷施四种钾肥对葡萄坐果率的影响均达到了显著水平。其中K2SO4·2MgSO4效果最佳,在0.2%的喷施浓度下坐果率高达48.3%,比对照增长19.8%。K2SO4效果次之,其喷施浓度为0.5%时坐果率最高,为46.33%,比对照提高14.9%。K2CO3和KCl效果最差,其喷施浓度均为0.5%时坐果率最高,分别为43.63%和43.30%,分别比对照提高8.1%和7.4%(表2)。

表2 不同钾肥不同喷施浓度下葡萄的坐果率(%)
注(Note): 数据后不同字母表示差异达到5%显著水平 Values followed by different letters mean significant difference among treatments at 5% level.
2.2叶面喷施不同浓度钾肥对叶片和新梢生长的影响
葡萄叶片和新梢是葡萄的重要营养器官,叶面喷肥可以直接作用于叶片和新梢,对其生长有着巨大影响。本实验结果显示盛花期和果实膨大期喷施钾肥均可以显著影响叶片和新梢的生长。四种钾肥均以K2SO4·2MgSO4效果最佳,其在盛花期喷施浓度为0.2%时叶片增长率和新梢增长率最高,果实膨大期喷施浓度为0.5%时叶片增长率最高,喷施浓度为0.2%时新梢增长率最高。K2CO3效果次之,其在盛花期的喷施浓度为0.5%时新梢和叶片的增长率最高,在果实膨大期的喷施浓度为0.8%时新梢和叶片增长率最高。K2SO4和KCl效果最差,这两种钾肥在盛花期的喷施浓度为0.2%时新梢和叶片的增长率较高,而在果实膨大期的喷施浓度为0.5%时新梢和叶片的增长率较高(表3)。

表3 不同钾肥不同喷施浓度下葡萄叶片和新梢增长率(%)
注(Note): 数据后不同字母表示差异达到5%显著水平 Values followed by different letters mean significant difference among treatments at 5% level.
2.3叶面喷施不同浓度钾肥对叶片叶绿素含量的影响
四种钾肥在不同喷施浓度下对葡萄叶片叶绿素含量的影响均达到了显著水平。其中K2SO4·2MgSO4效果最显著,盛花期喷施浓度为0.2%时叶绿素含量最高,果实膨大期喷施浓度为0.5%时叶绿素含量最高。K2SO4和K2CO3效果次之,盛花期两种钾肥喷施浓度均为0.5%时葡萄叶片叶绿素含量最高,果实膨大期K2SO4喷施浓度为0.5%时叶绿素含量最高,K2CO3喷施浓度为0.8%时叶绿素含量最高。KCl效果最差,其在盛花期和果实膨大期的喷施浓度均为0.5%时叶绿素含量最高(表4)。
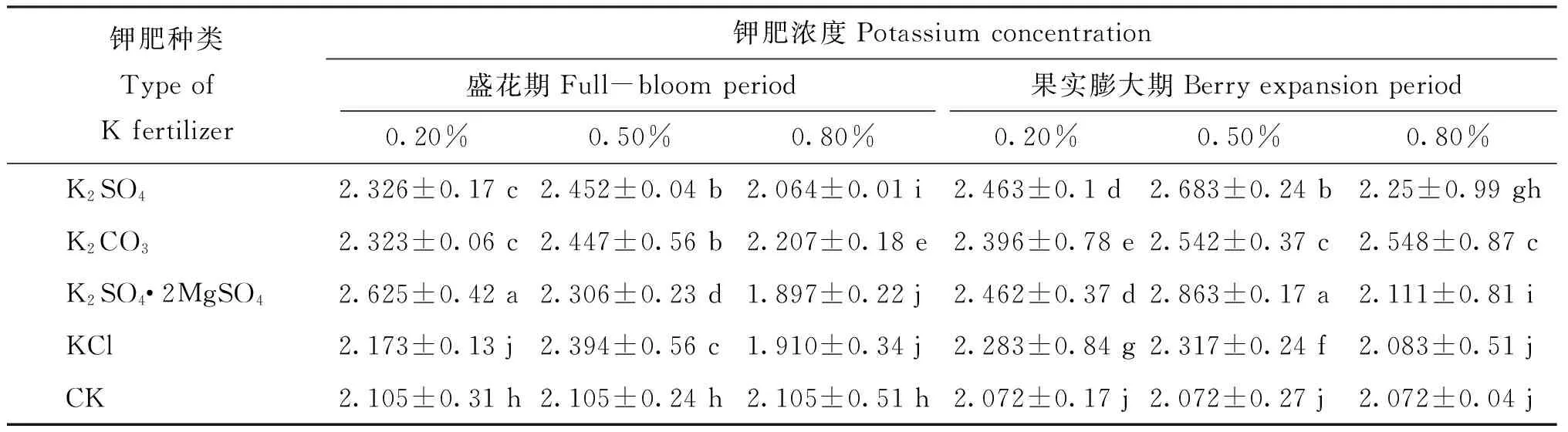
表4 不同钾肥不同喷施浓度下叶片叶绿素含量(mg/g, FW)
注(Note): 数据后不同字母表示差异达到5%显著水平 Values followed by different letters mean significant difference among treatments at 5% level.
2. 4叶面喷施不同浓度钾肥对葡萄果实品质的影响
葡萄果实可溶性固形物含量主要指葡萄果实中可溶性糖类物质的积累量,葡萄单果重则指葡萄单粒果实的平均重量,这两项指标是评价葡萄果实品质的重要参考。本实验结果显示葡萄叶面喷施不同种类及浓度钾肥对这两项指标的影响差异较大。K2SO4·2MgSO4效果最显著,其在盛花期的喷施浓度为0.2%时果实单粒重和可溶性固形物均达到最高,在果实膨大期喷施浓度为0.5%时两项指标均达到最高。K2SO4和K2CO3效果次之,其中K2SO4在两个时期的喷施浓度均为0.5%时葡萄果实单粒重和可溶性固形物达到最高,而K2CO3在盛花期的喷施浓度为0.5%时果实单粒重和可溶性固形物含量最高,在果实膨大期喷施浓度为0.8%时单粒重最高,喷施浓度为0.5%时可溶性固形物含量最高。KCl效果最差,其对单粒重和果实可溶性固形物的影响均不显著,甚至对果实可溶性固形物产生了不利影响,例如在果实膨大期三个喷施浓度均降低了葡萄果实的可溶性固形物含量(表5)。

表5 不同钾肥不同喷施浓度下葡萄单粒重和可溶性固形物含量
注(Note): 数据后不同字母表示差异达到5%显著水平 Values followed by different letters mean significant difference among treatments at 5% level.
2.5叶面喷施不同浓度钾肥对钾吸收相关基因表达的影响
2.5.1 叶面喷施4种钾肥对VvHAK13表达的影响VvHAK13属于高亲和性钾离子转运体KUP/HAK/KT家族,不仅负责对K+的吸收,对K+在植物体内的运输也有着重要作用。本实验结果显示叶面喷施4种钾肥均对VvHAK13的表达产生了不同程度的抑制作用,VvHAK13的表达主要呈现先降低后升高和不断降低两种模式。在盛花期,K2SO4和K2CO3的喷施浓度不超过0.5%,K2SO4·2MgSO4和KCl的喷施浓度不超过0.2%时,VvHAK13的表达呈先降低后升高趋势,超过时,VvHAK13的表达则呈不断降低趋势。在果实膨大期K2SO4、 K2SO4·2MgSO4和KCl的喷施浓度不超过0.5%时VvHAK13的表达均呈先降低后升高趋势,超过0.5%时VvHAK13的表达则呈不断降低趋势,而K2CO3在三个喷施浓度下VvHAK13的表达均呈先降低后升高趋势(图1)。

图1 不同钾肥不同喷施浓度下VvHAK13的表达Fig.1 Relative expression of VvHAK13 under different K fertilizers with different spraying concentrations
2.5.2 叶面喷施4种钾肥对VvKEA2表达量的影响VvKEA2编码K+/H+反向转运体,属于CPA2家族,主要参与钾的装载,如在液胞中,KEA可能通过质子泵介导H+/K+的交换,从而完成钾的装载。本研究结果显示,两个时期喷施4种钾肥均明显促进了VvKEA2的表达。喷施K2SO4·2MgSO4对VvKEA2表达的促进作用最明显,K2SO4和K2CO3次之,KCl最差。盛花期K2SO4和K2CO3喷施浓度为0.5%时VvKEA2的表达量最高,而K2SO4·2MgSO4和KCl的喷施浓度为0.2%时VvKEA2的表达量最高,在最适喷施浓度下该时期VvKEA2的表达量呈先提高后降低趋势。果实膨大期K2CO3的喷施浓度为0.8%时VvKEA2的表达量最高,其余三种钾肥的喷施浓度均为0.5%时VvKEA2的表达量最高,在最佳喷施浓度下该时期VvKEA2的表达均随时间延长不断提高,说明该时期钾肥的效果比盛花期更加持久(图2)。
2.5.3 叶面喷施4种钾肥对VvSIRK表达量的影响VvSIRK属于Shaker钾离子通道中内向整流K+通道基因,主要负责钾离子的低亲和吸收。本实验研究结果显示叶面喷施K2CO3对VvSIRK的影响最为明显,其次是K2SO4·2MgSO4、 K2SO4和KCl。叶面喷肥处理后VvSIRK在两个时期的表达均呈先升高后降低趋势。盛花期四种钾肥的喷施浓度均为0.2%时VvSIRK的表达量达到最高。果实膨大期K2SO4·2MgSO4的喷施浓度为0.2%时VvSIRK的表达量最高,K2SO4和KCl的喷施浓度为0.5%时VvSIRK的表达量最高,而K2CO3的喷施浓度为0.8%时VvSIRK的表达量最高。相比于其他3个基因,VvSIRK的表达受到低浓度钾肥的影响较为明显,尤其在盛花期,几种钾肥均在0.2%的喷施浓度下使VvSIRK的表达量达到最高(图3)。这可能与SIRK属于低亲和钾离子吸收通道,主要负责低浓度钾离子的吸收有关。
2.5.4 叶面喷施4种钾肥对VvSORK表达量的影响
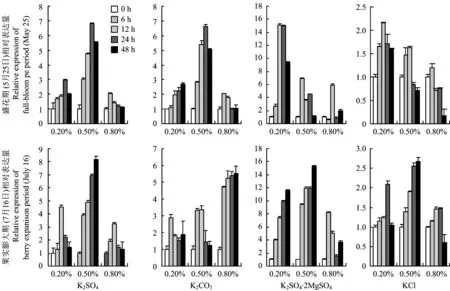
图2 不同钾肥不同喷施浓度下VvKEA2的表达Fig.2 Relative expression of VvKEA2 under different K fertilizers with different spraying concentrations

图3 不同钾肥不同喷施浓度下VvSIRK的表达Fig.3 The expression of VvSIRK under different K fertilizers with different spraying concentrations
VvSORK属于Shaker钾离子通道中外向整流K+通道,在植物地上部K+的积累中起着重要的作用。实验结果表明叶面喷施K2SO4·2MgSO4对VvSORK的影响最明显,其次是K2SO4、 K2CO3和KCl。盛花期K2SO4和K2CO3的喷施浓度为0.5%时VvSORK的表达量最高,K2SO4·2MgSO4和KCl的喷施浓度均为0.2%时VvSORK的表达量最高。果实膨大期K2SO4、 K2SO4·2MgSO4和KCl的喷施浓度均为0.5%时VvSORK的表达量最高,而K2CO3的喷施浓度为0.8%时VvSORK的表达量最高。盛花期喷施K2SO4和KCl后VvSORK的表达量先升高后降低,而在果实膨大期喷施后VvSORK的表达量则随时间延长不断升高。喷施K2CO3和K2SO4·2MgSO4后,VvSORK表现出独特的表达模式,喷施K2CO3后VvSORK的表达量先升高后降低再升高,而喷施K2SO4·2MgSO4后VvSORK的表达则先降低,随后又开始大幅度升高(图4)。
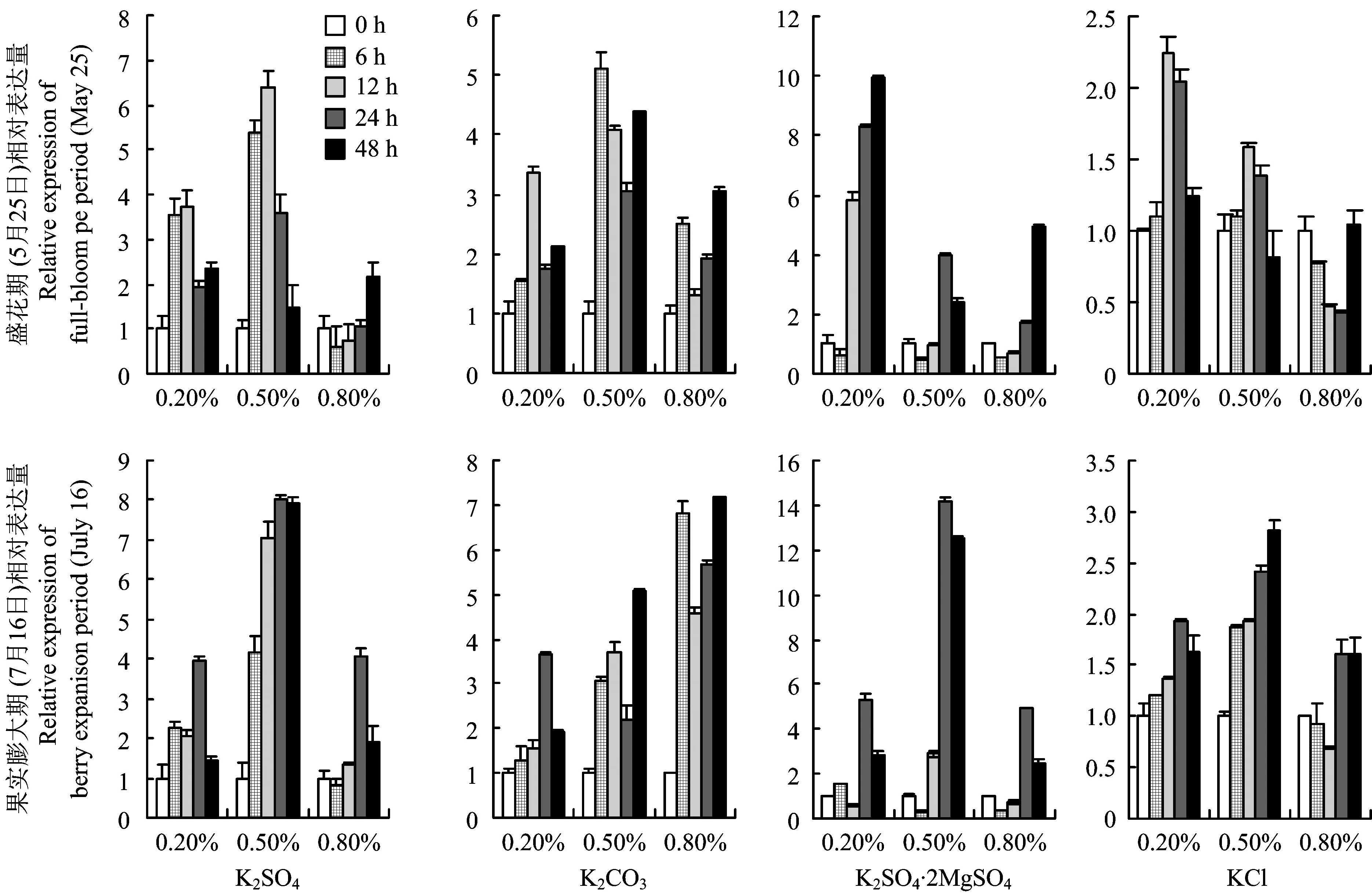
图4 不同钾肥不同喷施浓度下VvSORK的表达Fig.4 The expression of VvSORK under different K fertilizers with different spraying concentrations
3 讨论
钾是植物细胞中最丰富的离子,在植物的生长和发育中起重要作用,如促进酶的活化、 蛋白质合成、 光合作用,维持渗透压、 调节叶片和气孔运动和细胞伸长等[20-25]。葡萄钾吸收、 转运机理的研究主要集中在根际施钾对葡萄浆果及葡萄酒品质的影响上[26-30],对于葡萄叶面喷施钾肥相关理论的研究尚且不多。叶面喷施钾肥技术已经在棉花[31-33]、 小麦[34]、 马铃薯[35]等作物上得到了广泛应用和深入研究,研究结果均表明叶面喷施钾肥可以有效提高作物的产量、 品质、 抗病性等。本研究表明对葡萄叶面喷施钾肥不仅可以促进葡萄叶片和新梢的营养生长,为后期果实的发育提供充足的物质基础,也可提高葡萄的坐果率和单粒重、 可溶性固形物等品质指标,有助于提升葡萄的产量和质量。

葡萄生长发育不同时期对营养的需求有所差异,因此各时期的施肥效果也不同,其中施肥效果最好的时期应为植物营养的最大效率期[41]。据Abha等的研究报道,在葡萄坐果30天后叶面喷施K2SO4,随着K2SO4浓度的增大,葡萄果实可溶性固形物总量及总糖含量明显增加[42]。马文娟等[43]、 张志勇等[44]研究显示,果实膨大期葡萄钾累积量最大,该时期葡萄浆果快速生长、 糖分快速积累,此时施用钾肥,既可满足葡萄成熟对钾的需求,又有助于提高葡萄品质。Pratelli等研究发现VvSIRK在葡萄果实快速发育时期具有较高的表达量[45],说明该时期葡萄植株需要从外界吸收较多的钾元素,以满足葡萄浆果快速膨大对营养元素的需求。本实验结果与前人研究较为相似。在葡萄果实膨大期喷施钾肥效果更佳,且肥效更加持久。在果实膨大期喷施钾肥后,葡萄叶片和果实各项生理指标提升幅度明显高于盛花期,该时期三个重要钾吸收基因VvSIRK、VvKEA2、VvSORK在最佳喷施浓度下均能保持较长时间的高表达水平,且表达量随时间延长不断升高,说明该时期喷施钾肥比盛花期喷施具有更高更持久的肥效。此外,果实膨大期葡萄对钾肥喷施液浓度要求更高,例如KCl和K2SO4·2MgSO4在该时期的最适喷施浓度为0.5%,而在盛花期则为0.2%。
本实验中所选的4个钾吸收相关基因所编码的蛋白是葡萄钾离子吸收、 转运的重要通道或载体,其表达量反映了葡萄植株中钾离子吸收运输的活跃程度,叶面喷施钾肥后,这些基因表达越活跃,说明其对钾的吸收效率越高,因此也越有利于葡萄叶片和果实等的生长发育。本实验结果也证明了这一观点,钾吸收基因的表达和葡萄生理指标的变化具有较高的一致性,叶面喷施钾肥后,基因响应剧烈、 持续时间长的,叶片和果实等生理指标提升也更明显。这也证明了利用基因表达水平评价钾肥肥效的有效性。实验中喷施K2CO3和K2SO4·2MgSO4后VvSORK呈现出独特的表达模式,其表达量先下降后上升,或者先上升后下降再上升。Lacombe等的研究表明植物中SORK通道受到细胞内外pH的强烈影响[46],而K2CO3溶液呈弱碱性,K2SO4·2MgSO4溶液呈弱酸性,这可能是造成VvSORK独特表达模式的原因。这种现象也提示我们在对作物进行叶面喷肥的时候,肥料喷施液的pH也会影响植物对肥料的吸收效果,应当给予充分考虑。此外,VvHAK13和其他三个钾吸收相关基因不同,其表达受到了叶面喷施钾肥的抑制。Maser等,Ahn等[47-48]的研究表明缺钾可诱导KUP /HAK/KT家族高亲和性成员的表达,可能是因为缺钾时,KUP /HAK/KT家族高亲和性成员参与到不同组织或细胞间K+的转运以平衡K+的分布。然而当K+供应充分时,其转运、 平衡K+的作用降低,这可能是造成叶面喷施钾肥后VvHAK13表达量降低的原因。缺钾可以诱导VvHAK13的表达,这一特性使得VvHAK13可被当作“信号基因”用来预测葡萄对钾素的需求程度。当该基因表达量较高时,说明葡萄植株缺少钾元素,则可以通过叶面喷施钾肥快速补充钾元素。当其表达量较低时,则说明葡萄植株中钾元素充足,不需补充钾素。然而钾素对VvHAK13的抑制可能涉及更为复杂的机制,需要进一步研究。
4 结论
葡萄叶面喷施钾肥可以有效促进葡萄叶片和果实的生长发育,四种钾肥的效果依此为: K2SO4·2MgSO4>K2SO4>K2CO3>KCl。盛花期K2SO4和K2CO3的最适喷施浓度为0.5%,K2SO4·2MgSO4和KCl的最适喷施浓度为0.2%; 果实膨大期K2SO4、 K2SO4·2MgSO4和KCl的最适喷施浓度为0.5%,K2CO3的最适喷施浓度为0.8%。
适宜的喷施浓度可以有效提高钾吸收相关基因的表达,是其提高钾吸收利用的机理之一。
[1]赵祥奎, 张序, 姜远茂. 秋季叶施尿素对甜樱桃产量与品质的影响[J]. 山东农业大学学报(自然科学版), 2007, 38(3): 369-372.
Zhao X K, Zhang X, Jiang Y M. Studies on the effect of foliar spray of urea on the yield and quality of sweet cherry (Prunus.AviumL)[J]. Journal of Shandong Agricultural University (Natural Science Edition), 2007, 38(3): 369-372.
[2]贺普超. 葡萄学[M]. 北京: 中国农业出版社, 1999.
He P C. Viticulture[M]. Beijing: China Agriculture Press, 1999.
[3]孙周平, 刘涛, 蔺姗姗, 等. 雾培对番茄植株生长、 产量和品质的影响[J]. 沈阳农业大学学报, 2006, 37(3): 488-490.
Sun Z P, Liu T, Lin S S,etal. Effect of aeroponics on the growth, yield and quality of tomato plant[J]. Journal of Shenyang Agricultural University, 2006, 37(3): 488-490.
[4]Conradie W J, Saayman D. Effects of long-term nitrogen, phosphorus, and potassium fertilization on Chenin blanc vines. II. Leaf analyses and grape composition[J]. American Journal of Enology and Viticulture, 1989, 40(2): 91-98.
[5]Freeman B M, Kliewer W M. Effect of irrigation, crop level and potassium fertilization on Carignane vines. II. Grape and wine quality[J]. American Journal of Enology and Viticulture, 1983, 34(3): 197-207.
[6]Possner D R E, Kliewer W M. The localization of acids, sugars, potassium and calcium in developing grape berries[J]. Vitis, 1985, 24(4): 229-240.
[7]Delgado R, Martín P, del álamo M,etal. Changes in the phenolic composition of grape berries during ripening in relation to vineyard nitrogen and potassium fertilization rates[J]. Journal of the Science of Food and Agriculture, 2004, 84(7): 623-630.
[9]郑秋玲, 韩真, 王慧, 翟衡. 不同叶面肥对赤霞珠葡萄果实品质及树体贮藏养分的影响[J]. 中外葡萄与葡萄酒, 2009, 7: 13-16, 19.
Zheng Q L, Han Z, Wang H, Zhai H. Effects of different foliar fertilizers on fruit quality and storage nutrient of Cabernet Sauvignon grapevine[J]. Sino-Overseas Grapevine & Wine, 2009, 7: 13-16, 19.
[10]Boss P K, Thomas M R. Association of dwarfism and floral induction with a grape ‘green revolution’ mutation[J]. Nature, 2002, 416(6883): 847-850.
[11]Jaillon O, Aury J M, Noel B,etal. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla[J]. Nature, 2007, 449:463-467.
[12]房经贵, 刘洪, 杨光. 基因组学研究时代的葡萄酿造明星—黑比诺[J]. 中外葡萄与葡萄酒, 2008, 4: 57-59.
Fang J G, Liu H, Yang G. Wine grape star in the genomics era—pinot noir[J]. Sino-Overseas Grapevine & Wine, 2008, 4: 57-59.
[13]初建青, 王文艳, 房经贵, 等. 叶面喷施尿素对葡萄氮代谢相关基因表达的影响[J]. 植物营养与肥料学报, 2012, 18(2): 405-416.
Chu J Q, Wang W Y, Fang J G,etal. Effect of foliar applied urea on the expression of genes related to nitrogen metabolism in Fujiminori grapevine[J]. Plant Nutrition and Fertilizer Science, 2012, 18(2): 405-416.
[14]初建青, 岳林旭, 房经贵, 等. 尿素对葡萄5个氮代谢相关基因表达的影响[J]. 园艺学报, 2013, 40(2): 221-230.
Chu J Q, Yue L X, Fang J G,etal. Effects of fertilizer application on expression of genes related to nitrogen metabolism inFujiminorigrapevine[J]. Acta Horticultural Sinica, 2013, 40(2): 221-230.
[15]Wang C, Han J, Shangguan L F,etal. Depiction of grapevine phenology by genes expression information and test of its workability in guiding fertilization[J]. Plant Molecular Biology Reporter, 2014, 32(5): 1070-1084.
[16]郝再彬, 苍晶晶, 徐仲, 等. 植物生理实验[M]. 哈尔滨: 哈尔滨工业大学出版社, 2004. 46-49.
Hao Z B, Cang J J, Xu Z,et.al. Plant physiology experiment[M]. Harbin: Harbin Institute of Technology Press, 2004. 46-49.
[17]叶济宁. 关于叶绿素含量测定中的Arnon计算公式[J]. 植物生理学通讯, 1985, (6): 69.
Ye J N. Arnon formula in the measurement of chlorophyll content[J]. Plant Physiology Communications, 1985, (6): 69.
[18]张彦苹, 王晨, 于华平, 等. 适于葡萄不同组织RNA提取方法的筛选[J]. 西北农业学报, 2010, 19(11): 135-140.
Zhang Y P, Wang C, Yu H P,etal. Screening of RNA extraction methods for various grapevine organs and tissues[J]. Acta Agriculturae Boreali-Occidentalis Sinica, 2010, 19(11): 135-140.
[19]Chang S J, Puryear J, Cairney J. A simple and efficient method for isolating RNA from pine trees[J]. Plant Molecular Biology Reporter, 1993, 11(2): 113-116.
[20]黄显淦, 王勤, 赵天才. 钾素在我国果树优质增产中的作用[J]. 果树科学, 2000, 17(4): 309-313.
Huang X G, Wang Q, Zhao T C. Effects of potassium fertilizers on improving quality and production of fruit crops[J]. Journal of Fruit Science, 2000, 17(4): 309-313.
[21]Maathuis F J M, Sanders D. Mechanisms of potassium absorption by higher plant roots[J]. Physiologia Plantarum, 1996, 96(1): 158-168.
[22]Maathuis F J, Ichida A M, Sanders D,etal. Roles of higher plant K+channels[J]. Plant Physiology, 1997, 114(4): 1141.
[23]Smart C J, Garvin D F, Prince J P,etal. The molecular basis of potassium nutrition in plants[J]. Plant and Soil, 1996, 187(1): 81-89.
[24]Thiel G, Wolf A H .Operation of K+-channels in stomatal movement[J]. Trends Plant in Science, 1997, 2(9) : 339-345 .
[25]唐晓东, 张洁, 韩胜芳, 王冬梅. 采收后喷施KH2PO4对四倍体“玫瑰香”葡萄枝条成熟的影响[J]. 河北农业大学学报, 2011, 34(1): 33-37.
Tang X D, Zhang J, Han S F, Wang D M. Effect of foliar application of KH2PO4after harvest on the maturation of branch for tetraploid grape (VitisviniferaL.cv"Muscat Hamburg")[J]. Journal of Agricultural University of Hebei, 2011, 34(1): 33-37.
[26]Davies C, Shin R, Liu W,etal. Transporters expressed during grape berry (VitisviniferaL.) development are associated with an increase in berry size and berry potassium accumulation[J]. Journal of Experimental Botany, 2006, 57(12): 3209-3216.
[27]Hale C R. Relation between potassium and the malate and tartrate contents of grape berries[J]. Vitis-Berichte ueber Rebenforschung mit Dokumentation der Weinbauforschung, 1977, 16: 9-19.
[28]Ruhl E H. Uptake and distribution of potassium by grapevine rootstocks and its implication for grape juice pH of scion varieties[J]. Animal Production Science, 1989, 29(5): 707-712.
[29]Morris J R, Sims C A, Cawthon D L. Effects of excessive potassium levels on pH, acidity and color of fresh and stored grape juice[J]. American Journal of Enology and Viticulture, 1983, 34(1): 35-39.
[30]Ruhl E H, Fuda A P, Treeby M T. Effect of potassium, magnesium and nitrogen supply on grape juice composition of Riesling, Chardonnay and Cabernet Sauvignon vines[J]. Animal Production Science, 1992, 32(5): 645-649.
[31]Oosterhuis D M, Wullschleger S D, Maples R L,etal. Foliar-feeding of potassium nitrate in cotton[J]. Better Crops with Plant Food, 1990, 74(3): 8-9.
[32]Sawan Z M, Mahmoud M H, El-Guibali A H. Response of yield, yield components, and fiber properties of Egyptian cotton (GossypiumbarbadenseL.) to nitrogen fertilization and foliar-applied potassium and mepiquat chloride[J]. Journal of Cotton Science, 2006, 10(4): 224-234.
[33]Weir B L. Effect of foliar applied potassium on cotton in the San Joaquin Valley of California [A]. Dugger P, Richter D. Proceedings of beltwide cotton conference [C]. Memphis, TN: National Cotton Council, 1999. 1307-1309.
[34]Kettlewell P S, Cook J W, Parry D W. Evidence for an osmotic mechanism in the control of powdery mildew disease of wheat by foliar-applied potassium chloride[J]. European Journal of Plant Pathology, 2000, 106(3): 297-300.
[35]Brar M S, Kaur N. Effect of soil and foliar applied potassium and nitrogen on yield of potato (Solanumtuberosum) in alluvial soils of Punjab, India[J]. The Indian Journal of Agricultural Sciences, 2006, 76(12):740-743.
[36]周淑惠, 方建苗, 陆宏,等. 含氯肥料对葡萄产量、 品质影响研究初报[J]. 葡萄栽培与酿酒, 1991, 4(3): 17-21.
Zhou S H, Fang J M, Lu H,etal. The influence of chlorine fertilizers on the yield and quality of grape[J]. Viticulture and Oenology, 1991, 4(3):17-21.
[37]朱本岳, 杨玉爱, 叶正钱, 陈雅云. 葡萄施钾效应的研究[J]. 浙江农业大学学报, 1995, 21(4): 429-430.
Zhu B Y, Yang Y A, Ye Z Q, Chen Y Y. Effect of potassium application in grapevine[J]. Journal of Zhejiang Agricultural University, 1995, 21(4): 429-430.
[38]Nahdi H, Benzina N, Mhiri A. Stalk necrosis and magnesium-potassium nutrient balance of grapevines in Tunisia[C]. Annales de l'Institut National de la Recherche Agronomique de Tunisie. 1993, 66(1-2): 153-168.
[39]张漱茗, 闫华, 刘施辉, 闫晓松. 钾及钾镁肥配合对酿酒葡萄产量、 品质的效应[J]. 葡萄栽培与酿酒, 1998, (2): 9-11.
Zhang S M, Yan H, Liu S H, Yan X S. Effect of compound fertilizer of potash and magnesium on yield and quality of winegrape[J]. Viticulture and Oenology, 1998, (2): 9-11.
[40]亓桂梅. 葡萄钾、 镁肥试验初报[J]. 葡萄栽培与酿酒, 1993, (4): 12-14.
Qi G M. The experiment on grapevine K and Mg fertilizers[J]. Viticulture and Oenology, 1993, (4): 12-14.
[41]杜会英, 薛世川, 孙忠富. 不同用量腐植酸复合肥对葡萄叶片养分积累及其生理指标的影响[J]. 中国生态农业学报, 2007, 15(1): 49-51.
Du H Y, Xue S C, Sun Z F. Effects of different application rates of humic acid compound fertilizer on leaf nutrient accumulation and physiological mechanism of grape[J]. Chinese Journal of Eco-Agriculture, 2007, 15(1): 49-51.
[42]Abha J, Singh R P, Vinod K. Effect of ammonium sulphate and potassium dihydrogen phosphate on yield and quality of grape cv[J]. Perlette Recent-Horticulture, 1995, 2(2): 37-39.
[43]马文娟, 同延安, 王百祥, 杨莉芳. 葡萄树主要生长期内钾素的吸收与累积规律[J]. 西北农林科技大学学报(自然科学版), 2013, 41(9): 127-132.
Ma W J, Tong Y A, Wang B X, Yang L F. Annual change of potassium content and accumulation in grape tree[J]. Journal of Northwest A&F University (Natural Science Edition), 2013, 41(9): 127-132.
[44]张志勇, 马文奇. 酿酒葡萄‘赤霞珠’养分累积动态及养分需求量的研究[J]. 园艺学报, 2006, 33(3): 466-470.
Zhang Z Y, Ma W Q. Studies on the requirement and accumulative trend of nutrients in wine grape ‘Cabernet Sauvignon’[J]. Acta Horticulturae Sinica, 2006, 33(3): 466-470.
[45]Pratelli R, Lacombe B, Torregrosa L,etal. A grapevine gene encoding a guard cell K+channel displays developmental regulation in the grapevine berry[J]. Plant Physiology, 2002, 128(2): 564-577.
[46]Lacombe B, Pilot G, Gaymard F,etal. pH control of the plant outwardly-rectifying potassium channel SKOR[J]. FEBS Letters, 2000, 466(2): 351-354.
[47]Maser P, Thomine S, Schroeder J I,etal. Phylogenetic relationships within cation transporter families ofArabidopsis[J]. Plant Physiology, 2001, 126(4): 1646-1667.
[48]Ahn S J, Shin R, Sehachtman D P. Expression of KT/KUP genes inArabidopsisand the role of root hairs in K+uptake[J]. Plant Physiology, 2004, 134: 1135-1145.
Effectiveness and concentration of foliar-application of potassium fertilizers on grapevine evaluated by expression of potassium uptake related genes
ZHANG Cheng, JIA Hai-feng1*, WANG Jian1, JIU Song-tao, WANG Meng-qi1
(CollegeofHorticulture,NanjingAgriculturalUniversity,Nanjing210095,China)
【Objectives】 Foliar application of potassium(K) fertilizer has been widely adopted in grape production as an effective K supplement measurement. In this study, four types of K compounds were sprayed in three different concentrations at full-blooming stage and berry expansion stage of grapes, to study the responses of the physiological traits and the expression variation of K uptake related genes, and to evaluate the effectiveness of these K fertilizers and concentrations for practical grape production. 【Methods】 Grapevine cv.‘Summer Black’ (hybrid ofV.vinifera×V.labrusca) vines were used as experimental material, 0.2%, 0.5% and 0.8% solutions of K2SO4, K2CO3, K2SO4·2MgSO4and KCl were sprayed at the full-bloom stage and berry expansion stage. The growth rate of leaves and shoots, fruit setting rate, leaf chlorophyll content, single berry weight and total soluble solid content were measured, and the expression variation of the four K uptake related genes,VvHAK13,VvKEA2,VvSIRKandVvSORKwere analyzed using real-time quantitative PCR. 【Results】 All the foliar applications of K fertilizers are effective to improve the physiological indexes to different extents, and K2SO4·2MgSO4shows the best result, following are K2SO4, K2CO3, KCl. The optimum spraying concentrations are 0.2%-0.5% at full blooming stage and 0.5%-0.8% at berry expansion stage. Relatively higher K fertilizer concentration is required at berry expansion stage than at full blooming stage. The four K uptake related genes show different expression responses to the treatments. In general, the expression ofVvKEA2,VvSIRKandVvSORKare up-regulated, but that ofVvHAK13 down-regulated. In berry expansion stage, the four K uptake related genes show more remarkable responses to spraying concentrations of 0.5%-0.8%, but do the same to spraying concentration of 0.2%-0.5% at full-bloom stage.【Conclusions】 Spraying K fertilizer is effective to up-regulate the expression of K-related genes, which are reflected by the increased physiological indexes. Among the four tested fertilizers, K2SO4·2MgSO4shows the best result, following are K2SO4, K2CO3and KCl. Spraying concentration of 0.2%-0.5% is recommended in full-bloom stage and 0.5%-0.8% in the berry expansion stage of grape.
grapevine; foliar application; potassium fertilizer; potassium uptake gene; gene expression
2015-05-27接受日期: 2016-01-25
国家自然科学基金(31401847); 江苏省自然科学基金(BK20140707); 国家博士后基金(2014M561663); 中央高校特别资助(KJQN201541)资助。
张成(1988—), 男, 河南南阳人, 硕士, 主要从事果树营养研究。 E-mail: 2013104041@njau.edu.cn
E-mail: jiahaifeng@njau.edu.cn
S601; S663.1
A
1008-505X(2016)04-1091-11
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:08:14
气象水文海洋仪器(2022年2期)2022-07-08 09:48:48
农产品市场周刊(2022年4期)2022-05-21 15:15:28
农产品市场周刊(2022年4期)2022-03-23 22:34:20
今日农业(2021年8期)2021-07-28 05:56:04
河北果树(2020年2期)2020-01-09 11:15:07
今日农业(2019年11期)2019-08-15 00:56:32
今日农业(2019年11期)2019-08-15 00:56:32
山东林业科技(2019年2期)2019-06-03 10:11:02
中国果业信息(2017年4期)2017-05-09 08:36:39
