
热带次生林演替过程中环境因子对物种多样性的影响1)
2016-08-17 08:29龙成吴庭天杨小波龙文兴李东海
东北林业大学学报 2016年1期
龙成 吴庭天 杨小波 龙文兴 李东海
(海南大学,海口,570228) (热带作物种质资源保护与开发利用教育部重点实验室(海南大学))
热带次生林演替过程中环境因子对物种多样性的影响1)
龙成吴庭天杨小波龙文兴李东海
(海南大学,海口,570228)(热带作物种质资源保护与开发利用教育部重点实验室(海南大学))
为了解热带森林次生演替的群落环境特征变化对植物多样性的影响,以两种不同演替阶段的热带滨海次生林群落为研究对象,采用两种非参数检验以及Kendall秩相关分析的方法,对两群落内部的环境因子及其对植物多样性、均匀度以及物种周转率的影响进行研究。结果表明:各环境因子在群落间和群落内均存在差异。在热带常绿季雨林(TEMF)、灌木林(SHR)内,光合有效辐射(PAR)对物种多样性和周转率有显著影响。在两个群落内,土壤相对湿度对物种分布均起到重要影响,对SHR中物种分布的影响更强。在TEMF中,土壤温度、坡度及坡位对物种多样性和周转率限制显著;在SHR中,空气相对湿度成为限制物种分布的主要环境因子。
次生演替;环境因子;物种多样性;物种周转;物种均匀度;热带滨海次生林
In order to acquaint the effects of varying environmental factors on plant diversity during secondary succession of forests in tropics, our study was conducted in two types of tropical coastal forests which were called as tropical evergreen monsoon forest (TEMF) and shrubbery (SHR), respectively. Nine permanent plots of 50 m×50 m were established to investigate species composition and environmental factors. Meanwhile, two types of non-parametric tests and Kendall rank correlation analysis were used to study the difference of environmental factors between two communities as well as their effects on species diversity, species evenness, and species turnover. There were differences for each environmental factors between/within two types of communities. Species diversity and turnover were significantly affected by PAR within both TEMF and SHR. Species distributions within two types of communities, and in SHR especially were significantly influenced by soil relative moisture. In addition, species diversity and turnover were significantly limited by soil temperature, soil gradient, and slope position in TEMF, however, in SHR, air relative humidity became a main factor that could restrain plant species distributions.
热带次生林群落在我国热带地区分布广泛,它对林业生产以及生态系统服务有重要的作用[1-2]。森林群落物种生长速率较其他群落类型缓慢[3],尤其是在经历自然(极端的气候条件)或人为干扰(强烈的土地利用)后,次生森林群落物种组成的恢复速度远落后于群落结构的形成[4-5]。因此,如何恢复森林群落生物多样性,减少物种大量灭绝的危险,已成为当今热点议题[6-7]。森林群落演替通常伴随着群落内环境特征的改变和物种的取代[8-9]。在热带滨海次生林演替过程中,群落内变化的环境条件对物种的更新和共存起到怎样的作用,还尚未可知。因此,该研究不仅可以使我们了解滨海森林次生演替进程,还可以帮助我们揭示森林群落物种恢复和共存机制。
环境筛假说认为,物种在特定的环境下产生功能性适应,从而导致物种共存[10]。由此可见,环境因子在森林群落物种组成方面的重要作用。因此,近年来关于群落内环境条件对物种多样性影响的研究层出不穷。研究者们分别从纬度、海拔梯度、坡向、坡位、光照、温度、土壤以及岩石裸露度等[11-19]环境因子对植物多样性的影响进行了大量的研究。龙文兴等[20]针对海南岛霸王岭热带云雾林在雨季的环境特征进行了研究,说明环境因子对热带云雾林的植被分布具有重要影响。许涵等[21]通过对海南尖峰岭不同林型的热带雨林的研究,找出了与物种多样性相关联的环境因子。然而,这些研究多是基于森林同一演替阶段,并没有针对演替过程进行研究。海南岛东北部铜鼓岭自然保护区的热带滨海次生林,历史上经历了频繁的自然干扰(台风)和人为干扰(刀耕火种),演替过程较海南岛其他森林群落有所不同。因此,本研究以该类型森林两种不同演替阶段为研究对象,系统分析光合有效辐射、空气温度和相对湿度、土壤温度和相对湿度以及地形因子等环境特征,并进一步研究环境因子与物种多样性的关系,以期揭示热带滨海次生林演替过程中环境特征的变化及其对植物物种多样性的影响。
1 研究区概况
铜鼓岭国家级自然保护区位于海南岛东北部沿海的文昌市龙楼镇,区内分布的热带滨海次生林处于两种不同演替阶段,即热带常绿季雨林(TEMF)和灌木林(SHR),该地区相对稳定的气候顶级群落为热带常绿季雨林[9]。所有样地都是经人为和自然干扰后形成的次生林群落,均处于演替过程中。TEMF中优势物种为:鹅掌柴(Scheffleraheptaphylla(Linnaeus) Frodin)、假赤楠(SyzygiumbuxifolioideumHung T. Chang & R. H. Miao)、肖蒲桃(Syzygiumacuminatissimum(Blume) Candolle)、琼刺榄(Xantolislongispinosa(Merrill) H. S. Lo)和白茶树(Koilodepashainanense(Merrill) Airy Shaw)等。SHR中优势物种为:贡甲(Maclurodendronoligophlebium(Merrill) T. G. Hartley)、海南大风子(Hydnocarpushainanensis(Merrill) Sleumer)、柄果木(MischocarpussundaicusBlume)、云南银柴(Aporosayunnanensis(Pax & K. Hoffmann) F. P. Metcalf)和粗脉紫金牛(ArdisiacrassinervosaE. Walker)等。表1对两群落进行了概述。

表1 研究地概况
注:表中数值为“平均值±标准差”;同列不同小写字母表示差异显著(P<0.05)。
2 研究方法
2.1样地设置
在热带常绿季雨林样地和灌木林样地内分别设置9个50 m×50 m的固定样方,共18个,总面积4.5 hm2,样方编号为TEMF1~TEMF9和SHR1~SHR9。TEMF和SHR中处于上、中、下坡位的样方均为3个,用相邻格子法将每个样方划分成5 m×5 m的小样方,共计1 800个。调查小样方内所有胸径(DBH)大于等于1 cm的乔灌木以及层间藤本植物,乔灌木测定胸径、高度,层间藤本植物只测定胸径大小。
2.2数据采集
光合有效辐射。在晴朗无云天气,采集TEMF和SHR共18块样地内的光合有效辐射值(PAR)。采样时,沿每个样方对角线均匀地设置5个采样点,手持光合辐射仪,在每个点距地面1.3 m处测定PAR值。在TEMF和SHR内分别均匀设置5个取样点,从每天8:00—16:00,间隔2 h采集1次,连续监测4个月。数据采集时间从2015年5—8月份。
空气温度、空气相对湿度。每个样地用一个HOBO Pro温湿度自动记录仪置于样地中央,距地面高度1.3 m处,每隔2 h测定1次数据。以便分析空气温度和相对湿度日变化和月变化情况。数据采集时间从2015年5—8月份。
土壤温度、湿度的采集。在每个样地内沿对角线分别设置5个点,用土壤水矿化度和氧合状态监测装置测定土壤温度、湿度。在2015年5月至8月,在晴朗天气8:00—16:00,每隔2 h测量1次。
森林郁闭度的测定。在每个样地内,均匀设置100个观测点,对林分郁闭情况进行记录、测定。如果观测点上方被林冠遮挡,则记为“1”,否则记“0”。然后,计算被遮挡的观测点数占总观测点数的比例,即为林冠郁闭度。
地形数据。地形数据包括坡度、坡向、坡位和岩石裸露比例等。在每个样地内,用坡度坡向仪测定坡度和坡向,并且估测样地所处坡位和地表岩石裸露比例。
2.3植物物种多样性估计
以Shannon-Wiener(H)、Pielou指数(E)以及Sørensen相异性系数(CD)来估计物种多样性、物种均匀度和物种周转率。计算公式如下:

E=H/lnS;
CD=1-[2C/(S1+S2)]。
式中:S是出现在所有样方内的物种总数目;Pi是总物种中物种i的多度所占的比例;C是两个相邻样方之间共有种的数目;S1和S2分别是样方1和样方2中独有物种的数目。
2.4数据分析
本研究分别用Kruskal-Wallis检验和Wilcoxon符号秩检验比较同一群落内和不同群落间的光合有效辐射、空气温度、空气相对湿度、土壤温度、土壤相对湿度、坡度以及裸岩比例;环境因子与物种多样性、均匀度和周转率的关系采用Kendall秩相关分析;采用SPSS 19.0软件进行统计分析。
3 结果与分析
3.1林内光合有效辐射特征
由表2可知,TEMF和SHR一天中不同时间段的PAR最大值出现的时段不同。TEMF中,最高PAR出现在10:00—12:00间,而SHR中,12:00—14:00间PAR值则达到最大值。在8:00—10:00间,TEMF中的PAR显著大于SHR。在10:00—12:00间,群落间的PAR无显著差异。在12:00—14:00和14:00—16:00两个时间段内,SHR的PAR均显著大于TEMF。

表2 热带常绿季雨林(TEMF)和灌木林(SHR)光合有效辐射(PAR)日变化 μmol·m-2·s-1
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
由表3可知,TEMF内,同一时段的PAR均存在显著差异,在SHR中,相同时段的PAR均无显著差异。

表3 热带常绿季雨林(TEMF)和灌木林(SHR)内光合有效辐射的比较
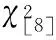
由表4可知,两群落内同时段的PAR在5月份至8月份变化情况相同,7月份达到最大值,随后逐渐下降。但是,在SHR中的8:00—10:00间,PAR的月最大值却出现在8月份。

表4 热带常绿季雨林(TEMF)和灌木林(SHR)日平均光合有效辐射 μmol·m-2·s-1

森林类型7月8:00—10:0010:00—12:0012:00—14:0014:00—16:008月8:00—10:0010:00—12:0012:00—14:0014:00—16:00TEMF(76.40±6.270)b(141.20±9.590)b(105.80±5.701)b(59.20±7.431)b(54.40±5.667)b(124.00±42.367)b(76.20±7.799)b(23.80±1.733)bSHR(107.20±14.247)a(241.40±80.402)a(542.20±78.949)a(278.20±22.326)a(142.80±15.932)a(209.00±49.745)a(488.00±49.805)a(228.20±72.559)a
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
3.2林内空气与土壤的温度、相对湿度特征
由表5可知,整体上,TEMF和SHR的空气温度最大值均出现在12:00—14:00。两群落在10:00—12:00、12:00—14:00和14:00—16:00空气温度均有显著差异。在8:00—12:00,两群落内空气温度上升速率无明显差异,但12:00后,SHR中空气温度明显高于TEMF。

表5 热带常绿季雨林(TEMF)和灌木林(SHR)空气温度(Ta)的日变化 ℃
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
两群落内空气相对湿度最小值出现在12:00—14:00。在12:00—14:00和14:00—16:00,群落间空气相对湿度存在显著差异。在12:00前,空气相对湿度随时间下降,但速率较小,在12:00之后,SHR中空气相对湿度的降低速率明显高于TEMF(见表6)。

表6 热带常绿季雨林(TEMF)和灌木林(SHR)空气相对湿度(RH)的日变化 %
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
在5至8月间,两群落内日平均空气温度与相对湿度的变化趋势相反。日平均空气温度在7月最高,而相应的空气相对湿度则最低。群落间每月的空气温度及湿度均呈现显著差异(见表7~8)。

表7 热带常绿季雨林(TEMF)和灌木林(SHR)月平均空气温度 ℃
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。

表8 热带常绿季雨林(TEMF)和灌木林(SHR)月平均空气相对湿度 %
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
TEMF和SHR中各样方间,除8:00—10:00时段的空气相对湿度无显著差异外,其他时段的空气温度和相对湿度均表现出明显的差异性(见表9~10)。
TEMF和SHR的土壤温度最大值均出现在12:00—14:00。群落间土壤温度仅在12:00—14:00和14:00—16:00时段存在显著差异。在8:00—12:00,TEMF中土壤温度的上升速率较SHR高,但12:00以后,SHR中土壤温度的升高速度却高于TEMF。群落间各时段的土壤相对湿度均差异显著,但群落内部变化较为平缓,基本呈现先下降后上升的趋势(见表11~12)。

表9 热带常绿季雨林(TEMF)和灌木林(SHR)内空气温度的比较

表10 热带常绿季雨林(TEMF)和灌木林(SHR)内相对湿度的比较

表11 热带常绿季雨林(TEMF)和灌木林(SHR)土壤温度(ST)的日变化 ℃
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。

表12 热带常绿季雨林(TEMF)和灌木林(SHR)土壤相对湿度(SM)的日变化 %
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
5月至8月间,两群落土壤温度和相对湿度的变化近似于空气温度和相对湿度的变化趋势相反。最高土壤温度和最低土壤相对湿度均出现在7月份。除7月份和8月份的土壤温度以外,群落间其他月份土壤温度以及相对湿度均呈现显著差异(见表13~14)。

表13 热带常绿季雨林(TEMF)和灌木林(SHR)月平均土壤温度 ℃
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。

表14 热带常绿季雨林(TEMF)和灌木林(SHR)月平均土壤相对湿度 %
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
群落内各时间段的土壤温度均存在显著差异;土壤湿度在TEMF和SHR中的8:00—10:00以及SHR中的10:00—12:00差异不明显,但其他时段均呈显著差异(见表15~16)。

表15 热带常绿季雨林(TEMF)和灌木林(SHR)内土壤温度的比较

表16 热带常绿季雨林(TEMF)和灌木林(SHR)内土壤湿度的比较
3.3地形特征和物种多样性
TEMF物种多样性、物种均匀度以及裸岩比例均显著大于SHR,然而,群落间坡度与物种周转率却无显著差异。但SHR的坡度还是大于TEMF,而物种周转率却小于TEMF。TEMF主要位于阳坡和半阳坡,而SHR主要位于阴坡和半阴坡。TEMF和SHR中处于上、中、下坡位的样方均为3个(见表17)。

表17 热带常绿季雨林(TEMF)和灌木林(SHR)地形因子与多样性指数的比较
注:表中数值为“平均值±标准差”;同列不同小写字母代表两者差异显著(P<0.05)。
3.4环境因子与物种多样性的关系
由表18可知,TEMF和SHR中,PAR与物种多样性及物种周转率显著正相关。空气相对湿度在两群落内与物种多样性和物种均匀度皆呈显著正相关,但仅在SHR中表现出与物种周转率的显著正相关。相似地,土壤温度仅在TEMF中与物种多样性和周转率呈显著正相关。土壤相对湿度在两类型群落内与物种多样性及周转率均呈显著正相关,但却与物种均匀度呈显著负相关。在TEMF中,物种多样性表现出与坡位的显著正相关,物种多样性与坡度显著负相关。无论在TEMF还是在SHR,空气温度与物种变量均无显著相关性。因此,可以看出,在TEMF中对物种多样性影响较大的环境因子为PAR、空气相对湿度、土壤温度及相对湿度、坡度与坡位等;而SHR中,PAR、空气相对湿度、土壤相对湿度对物种多样性有着重要影响。

表18 热带常绿季雨林(TEMF)和灌木林(SHR)中环境因子与物种变量的Kendall秩相关分析
注:括号内为检验数值P,*表示在0.05水平上显著相关,** 表示在0.01水平上显著相关。
4 讨论与结论
4.1群落光照特征与物种多样性
光照条件和光合作用之间的相互作用十分紧密,对于不同演替阶段群落中光照条件差异的研究,可以有效反映光因子在演替过程中的作用,以及对植物物种的影响。同时,光因子的差异也在一定程度上反映出不同演替阶段森林群落内部结构条件的差异[22]。TEMF林冠郁闭度大于SHR,这是森林演替过程必然导致的结果[22],也正由于这样的差异导致SHR中透过林冠的PAR整体上大于TEMF。然而,TEMF中8:00—10:00间的PAR却大于SHR,这一结果可能与两群落所处坡向条件有关。SHR主要位于西北坡向,而TEMF主要位于东南坡向。因此,在日出后的8:00—10:00,处于向阳坡的TEMF的PAR大于SHR。此后,随太阳高度角的增大,这种差异逐渐缩小,林冠的郁闭程度则成为林内光照情况的决定性因素,因此,在12:00—16:00,SHR内的PAR显著大于TEMF。此外,森林郁闭度及平均树高的差异也是导致TEMF中不同时段内各样方PAR存在显著差异的主要原因。整体上,5月至8月间,日平均PAR不同时段的变化呈趋势一致。然而,相较TEMF而言,SHR在8:00—10:00日平均PAR却在8月份达到最值,这同样可能由SHR群落的整体朝向导致。
在TEMF和SHR中,PAR与物种多样性和物种周转率均呈显著正相关。在两种群落中,PAR值显著地促进物种数量的增多以及转换速率,同时也是这二者的限制性因子。森林群落中的光因子对植物生长以及幼苗更新具有重要影响[18],随着群落演替的进行,森林群落结构的变化必然导致林冠下光照环境相应的改变[22],这种改变增加了群落内光因子的异质性程度,因而,在TEMF中能够创造出更多的更新生态位,使对光环境有不同反应及适应能力的物种得以共存[23]。物种对群落内光照异质性环境的长期适应,使其产生相应的趋光适应结构。例如,TEMF林下光照整体上低于SHR,因此,TEMF中林下层植物种的平均高度较SHR高,以便在林冠郁闭的林下获得水平方向上稀少的生长与更新不可或缺的光资源。一般来说,幼苗对光照条件的反应比成年植株更敏感[24]。SHR中的高光照条件且异质程度较低,可以使许多阳生物种的幼苗更新良好,但对于演替后期耐阴物种幼苗的更新却较为不利。而TEMF内较大的异质性光照条件可以为不同植物种的幼苗提供存活和更新机会,这就使TEMF中物种多样性水平高于演替早期的SHR。因此,在TEMF和SHR中,PAR对于物种多样性增大以及物种在群落内的迁移速率均起到促进作用,且该作用在光照条件异质性较强的TEMF中会表现得较为明显。但从森林群落长期演替来看,越接近演替末期,TEMF林冠层郁闭度越高,更加有利于耐阴物种的生长,但危及阳生物种的存活,即使少量林窗环境短暂出现,也不会改变在演替末期群落物种多样性下降的趋势。而SHR中,林冠下的光照条件由于林冠逐渐郁闭而稀缺,使植物向垂直方向趋光生长,这就导致林冠郁闭程度逐渐增大,从而使更多的耐阴物种得以“栖息”,因此,在未来的演替阶段,SHR中的物种多样性将有所增加[25]。
4.2群落空气温度、相对湿度与物种多样性
TEMF位于山顶周围,海拔高度、林冠郁闭度、植株平均高度均显著大于SHR。总体上,在5月至8月间,TEMF与SHR之间的日平均空气温度、相对湿度均存在显著差异,且变化趋势相同。在8:00—10:00,PAR差异显著,但空气温度的升高往往较为滞后[18],尤其在林冠郁闭度和植株高度较大的TEMF中更是如此。此后,空气温度则逐渐升高。然而,郁闭度的差异导致群落间空气温度上升速率不同,SHR空气温度在随后的各时段均显著高于TEMF。除此之外,TEMF海拔高度大于SHR,也会导致TEMF中的空气温度低于SHR[20]。TEMF和SHR中空气相对湿度的变化滞后于空气温度,且整体上与空气温度变化趋势相反,这与龙文兴等[20]的研究结果一致。在8:00—10:00和10:00—12:00,群落间空气相对湿度差异并不明显,但12:00—16:00,二者的差异性趋势逐渐显露。随着空气温度逐渐升高,相对湿度也就随之降低,较小的林冠郁闭度使SHR中空气相对湿度下降较TEMF快,但整体上变化趋势相同。尽管SHR中PAR差异并不显著,但差异依然存在;另外,植物也对群落内部空气温度具有一定调节作用[20]。因此,无论在TEMF还是SHR中空气温度都表现出空间异质性。林内空气温度的异质性特征对空气相对湿度影响较大。除8:00—10:00空气相对湿度无明显差异之外,两群落空气相对湿度在其他各时段均呈极显著差异。这表明空气温度对湿度具有调节作用[20],且林冠郁闭程度也间接影响空气相对湿度的变化过程。
在TEMF和SHR中,空气温度与物种多样性、均匀度以及周转率均无显著相关性。然而,空气相对湿度却对这些变量影响较大。这表明在森林生态系统中,较高的空气相对湿度是树木生长的重要条件之一[26],且研究中的空气温度并不直接影响物种多样性分布格局,这与温度可以影响高海拔森林群落植物生长、贮存水分和养分能力的观点相反[27]。但是,空气温度可以对空气相对湿度进行调节[20],间接影响群落内物种分布。在SHR中,空气相对湿度显著地促进物种周转速率,这说明该群落内部的植物物种对空气相对湿度的响应较TEMF敏感。一方面,可能由于空气相对湿度可以对森林中动植物特别是附生的苔藓、地衣、蕨类和兰科植物等提供充足养分和水分[28];另一方面,SHR中PAR的异质性及林冠郁闭度均不及TEMF,显著影响林内空气中水分的保持,致使SHR内环境较为干燥。因此,SHR中植物物种周转速率被空气相对湿度显著影响,但随着演替的进行,群落结构发生改变,演替后期的TEMF由于林冠高大,林内郁闭度高,空气相对湿度显著大于SHR,因而在TEMF中,该现象并不明显。经过环境筛长期筛选,SHR中的物种多属耐干旱型,但与TEMF中物种一样,其生长过程中需要充足的水分,以满足种子萌发[27]、植物体生理生化过程以及形态特性的需要[29]。因此,空气相对湿度皆促进物种多样性的提高。随着演替过程中群落内环境的变化,环境筛主要根据群落中空气温度与空气相对湿度的不同,筛选出不同的植物物种[30]。其中,适合在SHR中生长的物种,多为高度较矮小的灌木型植物,且较为耐旱,迁移与更新活动对水分变化更敏感。相反,由于TEMF空气相对湿度较SHR大,为植物生长提供更为丰富的水分条件[20],TEMF植株平均高度较大,又促进了光照条件异质性的增强,因此,耐旱植物与耐阴蔽植物被同时选择共存于群落中,表现出TEMF中的物种多样性显著大于SHR。随着森林演替的进行,林分郁闭程度相应增大,TEMF和SHR的空气相对湿度会随之增大,保证了更多物种存活所需水分条件。这时,SHR中的光照条件或许还可使阳生物种与耐阴物种共存,但TEMF内接近完全郁闭的环境,却不利于更多阳生物种幼苗存活。因此,在未来的演替期间,TEMF中的物种多样性会有所降低,而SHR却反而增大[25]。
4.3群落土壤温度、相对湿度和地形因子与物种多样性
地形因子,如坡度、坡位和裸岩比例等对土壤理化性质产生重要影响,间接影响植物物种的分布情况[16,31]。坡度愈大,则土壤水分流失愈严重,TEMF中土壤相对湿度大于SHR。因此,在SHR中,土壤相对湿度对物种多样性及周转速率的影响更为重要。SHR中的土壤温度显著高于TEMF,且在两群落内都表现出较强的异质性。较高的土壤温度,使土壤中水分蒸发速度加快,直接导致SHR土壤相对湿度显著小于TEMF,进而影响植株与种子内部生理生化过程[32]以及植物体外部形态特征[29],致使部分植株矮化、植物体生长发育和种子萌发受阻。但对于经过环境筛选择的耐干旱植物种来说[30],也许可以在SHR的土壤条件下更新繁衍,然而,这种高温低湿的土壤条件并不利于更多耐阴物种及其幼苗的存活,这就限制了SHR中物种多样性的发展。由环境筛选择在TEMF中生存的多数物种,耐土壤高温和土壤水分贫瘠的能力远不如SHR中的植物种,而过高的土壤温度反而会限制植物种子萌发[33-34],因此,TEMF中异质性土壤温度就成为影响物种丰富度和群落内物种周转的关键因素,甚至可能决定局部的物种组成特征[10,30],而在SHR中,由于环境的筛选,耐干旱瘠薄土壤条件的植物被保留下来,土壤温度对多数具有耐旱特性植物种子的影响并不明显,因此,土壤温度在SHR中对物种多样性以及周转不构成实质性影响。再者,TEMF的裸岩比例显著大于SHR,使TEMF内微地形条件较SHR复杂和异质,导致生态位进一步细化,例如,群落内裸岩比例的异质性可以使与裸岩比例较高和较低相关的物种同时存活下来[12]。综上,TEMF的物种较SHR丰富。此外,两群落内土壤相对湿度与物种均匀度显著负相关,这表明土壤水分条件较好的地区更适合于植物的聚集分布[35]。有研究表明,物种的聚集分布有利于发挥整体效应[36],从而更好地抵御环境胁迫条件,促进个体的生存和繁衍。在SHR中,土壤水分的贫瘠程度远胜于TEMF,因此,生长在SHR中的植物物种可能经过长期的坏境筛选,形成了以聚集分布方式更有效地利用有限的土壤水资源,抵御水分胁迫的影响。相应地,TEMF中物种均匀度就显著高于SHR。
[1]ASNER G P, RUDEL T K, AIDE T M, et al. A contemporary assessment of change in humid tropical forests[J]. Conservation Biology,2009,23(6):1386-1395.
[2]LONG W X, YANG X B, LI D H. Patterns of species diversity and soil nutrients along a chronosequence of vegetation recovery in Hainan Island, South China[J]. Ecological Research,2012,27(3):561-568.
[3]CHAI S L, TANNER E V J. 150-year legacy of land use on tree species composition in old-secondary forests of Jamaica[J]. Journal of Ecology,2011,99(1):113-121.
[4]BREARLEY F Q, PRAJADINATA S, KIDD P S, et al. Structure and floristics of an old secondary rain forest in Central Kalimantan, Indonesia, and a comparison with adjacent primary forest[J]. Forest Ecology Management,2004,195(3):385-397.
[5]CHAZDON R L, LETCHER S G, BREUGEL M, et al. Rates of change in tree communities of secondary Neotropical forests following major disturbances[J]. Philosophical Transactions of the Royal Society of Loudon,2007,362:273-289.
[6]GARDNER T A, BARLOW J, PERRY L, et al. Predicting the uncertain future of tropical forest species in a data vacuum[J]. Biotropica,2007,39(1):25-30.
[7]LAURANCE W. Have we overstated the tropical biodiversity crisis[J]. Trends in Ecology & Evolution,2006,22(2):65-70.
[8]HUANG W J, LIU J X, WANG Y P, et al. Increasing phosphorus limitation along three successional forests in southern China[J]. Plant and Soil,2013,364(1/2):181-191.
[9]钟义,杨小波,符气浩,等.海南岛铜鼓岭自然保护区的植被与植物资源(续)[J].热带生物学报,1991,9(2):1-7.
[10]LOHBECK M, LOURENS P, MARTNEZ-RAMOS M, et al. Changing drivers of species dominance during tropical forest succession[J]. Functional Ecology,2014,28(4):1052-1058.
[11]贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
[12]任学敏,杨改河,王得祥,等.环境因子对巴山冷山—糙皮桦混交林物种分布及多样性的影响[J].生态学报,2012,32(2):0605-0613.
[13]徐远杰,陈亚宁,李卫红,等.伊犁河谷山地植物群落物种多样性分布格局及环境解释[J].植物生态学报,2010,34(10):1142-1154.
[14]唐志尧,方精云,张玲,等.秦岭太白山木本植物物种多样性的梯度格局及环境解释[J].生物多样性,2004,12(1):115-122.
[15]杨京彪,郭泺,周萤,等.坡位对红松阔叶林物种多样性的影响[J].东北林业大学学报,2015,43(1):28-31.
[16]张顺平,乔杰,孙向阳,等.坡向、坡位对泡桐人工林土壤养分空间分布的影响[J].中南林业科技大学学报,2015,35(1):109-116.
[17]陈圣宾,宋爱琴,李振基.森林幼苗更新对光环境异质性的响应研究进展[J].应用生态学报,2005,16(2):365-370.
[18]臧润国.海南热带山地雨林森林循环不同阶段光、温环境的测定与分析[J].北京林业大学学报,2002,24(5/6):125-130.
[19]赵景学,曲广鹏,多吉顿珠,等.藏北高寒植被群落物种多样性与土壤环境因子的关系[J].干旱区资源与环境,2011,25(6):105-108.
[20]龙文兴,丁易,臧润国,等.海南岛霸王岭热带云雾林雨季的环境特征[J].植物生态学报,2011,35(2):137-146.
[21]许涵,李意徳,骆土寿,等.海南尖峰岭不同热带雨林类型与物种多样性变化的环境因子[J].植物生态学报,2013,37(1):26-36.
[22]丁圣彦,卢训令,李昊民.天童国家森林公园常绿阔叶林不同演替阶段群落光环境特征比较[J].生态学报,2005,25(11):2863-2867.
[23]SWAINE M D, WHITMORE T C. On the definition of ecological species groups in tropical rain forests[J]. Plant Ecology,1988,75(1/2):81-86.
[24]韩有志,王政权.森林更新与空间异质性[J].应用生态学报,2002,13(5):615-619.
[25]ROXBURGH S H, SHEA K, WILSON J B. The intermediate disturbance hypothesis: patch dynamics and mechanisms of species coexistence[J]. Ecology,2004,85(2):359-371.
[26]李猛,段文标,陈立新.红松阔叶混交林林隙光量子通量密度、气温和空气相对湿度的时空分布格局[J].应用生态学报,2009,20(12):2853-2860.
[27]朱教君,李智辉,康宏樟,等.聚乙二醇模拟水分胁迫对沙地樟子松种子萌发影响研究[J].应用生态学报,2005,16(5):801-804.
[28]HOLDER C D. Rainfall interception and fog precipitation in a tropical montane cloud forest of Guatemala[J]. Forest Ecology and Management,2004,190(2/3):373-384.
[29]郑盛华,严昌荣.水分胁迫对玉米苗期生理和形态特征的影响[J].生态学报,2006,26(4):1138-1143.
[30]KRAFT N J B, ADLER P B, GODOY O, et al. Community assembly, coexistence and the environmental filtering metaphor[J]. Functional Ecology,2015,29(5):592-599.
[31]张地,张育新,曲来叶,等.坡位对东灵山辽东栎林土壤微生物量的影响[J].生态学报,2012,32(20):6412-6421.
[32]韦莉莉,张小全,侯振宏,等.杉木苗木光合作用及其产物分配对水分胁迫的响应[J].植物生态学报,2005,29(3):394-402.
[33]崔建垣,李玉霖,赵哈林,等.差巴嘎蒿(Artemisiahalodendron)种子萌发对土壤温度、水分和埋深的响应[J].干旱区资源与环境,2009,23(9):151-154.
[34]李俊清,牛树奎.森林生态学[M].北京:高等教育出版社,2006.
[35]RAYBURN A P, MONACO T A. Linking plant spatial patterns and ecological processes in grazed great basin plant communities[J]. Rangeland Ecology and Management,2011,64(3):276-282.
[36]苏爱玲,徐广平,段吉闯,等.祁连山金露梅灌丛草甸群落结构及主要种群的点格局分析[J].西北植物学报,2010,30(6):1231-1239.
Effects of Environmental Factors on Plant Diversity during Succession Processes of Tropical Secondary Forests//
Long Cheng(Hainan University, Haikou 570228, P. R. China); Wu Tingtian, Yang Xiaobo, Long Wenxing, Li Donghai
(Key Laboratory of Protection and Development Utilization of Tropical Crop Germplasm Resources, Ministry of Education, Hainan University)//Journal of Northeast Forestry University,2016,44(1):37-43.
Secondary succession; Environmental factor; Species diversity; Species turnover; Species evenness; Tropical coastal secondary forest
龙成,男,1986年5月生,海南大学环境与植物保护学院,博士研究生。E-mail:longcheng0509@163.com。
2015年9月16日。
S718.5
1)国家自然科学基金项目(31060073);国家重点基础研究计划前期项目(2010CB134512);海南文昌等沿海地区(铜鼓岭国家级自然保护区)森林生态系统监测基地项目。
责任编辑:王广建。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
女报(2020年7期)2020-08-17
河南科学(2020年3期)2020-06-02
水土保持研究(2019年6期)2019-10-19
儿童故事画报(2019年8期)2019-08-14
艺术启蒙(2018年8期)2018-08-22
中国水土保持科学(2017年3期)2017-07-05
浙江大学学报(工学版)(2016年2期)2016-06-05
草业科学(2015年9期)2015-12-11
