
杉木人工林凋落物中微生物数量对间伐的响应1)
2016-08-17 08:29肖文娅费菲刁娇娇关庆伟
东北林业大学学报 2016年1期
肖文娅 费菲 刁娇娇 关庆伟
(南京林业大学,南京,210037)
杉木人工林凋落物中微生物数量对间伐的响应1)
肖文娅费菲刁娇娇关庆伟
(南京林业大学,南京,210037)
为了明晰凋落物中微生物数量对间伐强度的响应程度,以南京溧水林场间伐7 a后25年生的杉木(Cunninghamialanceolata(Lamb.) Hook.)人工林为对象,研究了不同间伐强度对凋落物中真菌、细菌、放线菌3类微生物数量季节性变化的影响。结果表明:不同强度间伐杉木人工林下凋落物中微生物的数量随季节变化差异显著,微生物数量在春季期间对间伐响应最为显著。与对照相比,春季内弱度间伐下细菌数量显著减少了61.05%(P<0.01);而春季内中度间伐下的真菌和细菌数量分别减少了25.32%(P<0.01)和36.14%(P<0.01),放线菌则增加了30.97%(P<0.05);春季内强度间伐下真菌量和放线菌数量分别增加了77.64%(P<0.05)和59.35%(P<0.01),细菌数量减少了42.11%(P<0.01)。其中,凋落物层温度及林下植被层Margalef指数对微生物数量的影响最为显著。微生物数量的影响会导致凋落物向土壤中输送的养分不同,对提高林分生产力有重要意义。
间伐强度;凋落物;真菌;细菌;放线菌;季节变化
In order to clarify the response of the microbial number to thinning intensities in the litter layer, the microorganisms’ number of fungi, bacteria and actinomycetes in litter was determined during season changes in different thinning intensities in a 25-year old Chinese fir (Cunninghamialanceolata(Lamb.) Hook.) plantation. The numbers of microorganisms in the litter were significantly different depending on the season in the different thinning intensity, especially in spring. Compared with the control, the number of bacteria in spring significantly was reduced by 61.05% (P<0.01) in the light intensity thinning. The number of fungi in litter was reduced by 25.32% (P<0.01) in middle intensity thinning in spring, and the number of bacteria was reduced by 36.14% (P<0.01), while the number of actinomycetes was increased by 30.97% (P<0.05). In high intensity thinning, the numbers of fungi and actinomycetes were increased by 77.64% (P<0.05) and 59.35% (P<0.01), respectively. The number of bacteria in spring decreased by 42.11% (P<0.01) in high intensity thinning. Among all the factors, the impact of litter layer temperature and the Margalef index of the forest floor on the numbers of microorganisms is the most significant. The differences of the numbers of microorganisms in litter will lead to different nutrients that delivered to the soil, and improve the forest productivity.
森林凋落物是森林植物在生长发育过程中新陈代谢的产物,在促进生态系统养分循环方面起着重要作用[1]。凋落物分解是生态系统物质循环和能量转换的主要途径,对维持森林土壤肥力有重要作用[2]。而凋落物的分解与其中的微生物种群数量关系密切[3]。
微生物数量及生物量的变化受凋落物质量、树种组成、土壤水热状况等多种因素的影响[4],与凋落物和土壤中的养分状况[5]和元素周转速率密切相关[6]。物种组成[7-8]不同,凋落物的质和量就不同,适于微生物生长的营养源也不同,因而微生物的种类和数量也不同[9]。凋落物组成类型的增加对凋落物中微生物数量提升作用显著[10]。光、热、水、气的分布以及土壤状况,对凋落物中微生物的结构以及多样性也会产生一定影响[11]。在以往的研究中,对凋落物层微生物的研究主要集中于群落及数量的改变[3,12],或单一的研究某一因子对微生物数量的作用[6,13],影响微生物数量的直接和间接因子往往非常复杂,而综合分析3大类群微生物量与凋落物量、化学组成、温度、湿度及林下植被层多样性关系的研究还较少。
间伐作为一种重要的森林经营技术,其通过改变林下微环境[14-15]、根系分泌和凋落物分解输入到土壤中的有机质数量与质量[16-17],最终影响微生物群落。目前,间伐对凋落物影响的研究主要集中于物质循环与分解[18-19],且有关杉木人工林中凋落物中可培养微生物数量的研究还相对鲜见。
因此,文中以南京溧水林场间伐7 a后25年生的杉木人工林为对象,调查了弱度、中度和强度间伐下凋落物中细菌、真菌和放线菌数量的季节性变化,分析了它们与温湿度、林下植被多样性、凋落物基质质量等的关系,旨在明晰凋落物中微生物数量对间伐强度的响应程度。为探讨间伐对凋落物分解的影响机制及制定科学的营林措施提供一定的科学依据。
1 研究地自然概况
研究地位于江苏省南京市溧水县林场的秋湖分场(31°52′~31°61′N,118°99′~119°07′E),地处丘陵岗地,平均海拔约100 m。研究区内岩石由石英粗石岩、粗安岩及砂岩组成,山地土壤多数地带性“黄棕壤”,厚度10~100 cm,微酸性,局部地段岩石裸露较多。气候属亚热带向温带过度的气候带,年平均气温15.5 ℃,年平均日照2 146 h,年平均降水1 005.7 mm,无霜期220 d,四季分明。土壤均为结构疏松的黄棕壤,含石量约为20%,枯枝落叶层厚度11.5~21.5 cm。植被以人工林和通过封山育林形成的次生林为主。林分类型主要有马尾松(PinusmassonianaLamb.)、杉木(Cunninghamialanceolata(Lamb.) Hook.)纯林,麻栎(QuercusacutissimaCarruth.)小叶栎(Q.cheniiNakai.)、杉木(Cunninghamialanceolata(Lamb.) Hook.)麻栎(QuercusacutissimaCarruth.)、马尾松(PinusmassonianaLamb.)麻栎(QuercusacutissimaCarruth.)混交林等。
2 材料与方法
2.1样地设置
试验在25年生的杉木人工林固定样地中进行。课题组于2006年在该林分中设置了12块20 m×20 m的典型样地,样地间有5 m以上的未间伐隔离区。根据树木生长状况和空间分布状态,采取随机区组试验方式对每块样地实施了弱度(约30%株数强度,下同)、中度(约50%)、强度(约70%)3种单株间伐处理和未间伐的对照组。凋落物取样时,统计样地内乔木(直径≥4 cm)的株数,每木检尺,实测胸径、树高。试验样地基本情况如表1所示。
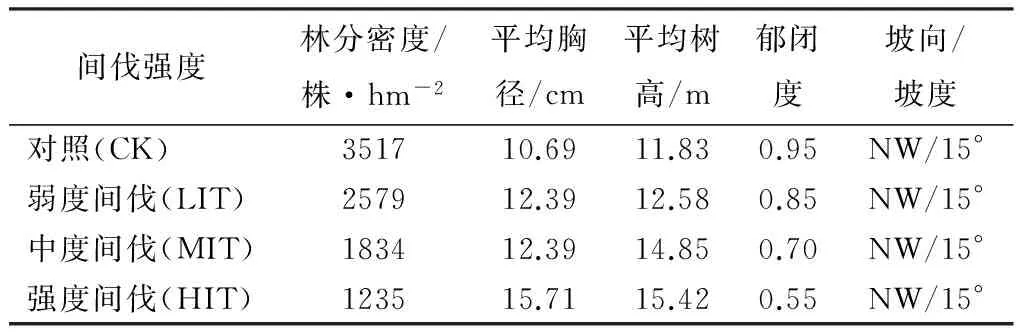
表1 2012年试验林分的基本状况
2.2研究方法
于2012年12月份在4种样地上进行凋落物收集,主要收集地面未分解的干枯枝叶。收集的凋落物称取干质量12 g装入大小为15 cm×25 cm孔径1 mm的凋落物袋中,每块样地布置40个凋落物袋,共480个。于2012年12月份将凋落物袋分别置于4种不同处理杉木人工林下,除去地表层的凋落物,置于土壤表面,让凋落物自然分解。分别于2013年3、6、9、12月份从4种样地中取回3~5个凋落物袋,装入保鲜袋后迅速带回实验室,进行室内分析。同时,于每个样地下凋落物分解袋中随机布置纽扣式温湿度记录仪(Hygrochron DS1923 USA)记录凋落物层温湿度的变化,设定频率为每小时记录1次实时温度和湿度,连续记录1a凋落物层温度和湿度的变化,并统计每月平均温度、最高温、最低温和平均相对湿度。凋落物中全碳、全氮质量分数采用燃烧法,用元素分析仪2004 II仪器测定,木质素和纤维素质量分数采用酸性洗涤法测定。
2.3微生物数量分析
微生物计数采用常规的平板稀释法。每种凋落物样品称取鲜质量5 g,置于45 mL的无菌水中,用摇床在室温下振荡30 min,在无菌环境下,移取10 mL菌液到90 mL无菌水中进行10倍稀释,形成10-1、10-2、10-3、10-44个体积分数梯度。真菌培养取体积分数10-3菌液0.1 mL涂布于马丁氏培养基,培养72 h;细菌培养取体积分数10-4菌液0.1 mL涂布于牛肉膏蛋白胨培养基,培养24 h;放线菌培养取体积分数10-3菌液0.1 mL涂布于改良高氏一号培养基,培养48 h。培养温度为(28±1) ℃,计算菌落数,每个样品每种培养基各3皿,取其平均值。称一定量的凋落物,80 ℃烘干至恒质量,称干质量,计算含水量。
2.4数据处理
通过Microsoft Excel 2007整理数据。通过SPSS16.0软件采用单因子方差分析(ANOVA)的最小显著差异法(LSD)检验不同季节、不同间伐下凋落物微生物数量间的显著性。利用Canoco4.5软件对数据进行冗余分析和作图。
3 结果与分析
3.1杉木人工林下凋落物中微生物数量
由表2可见,与对照样地相比,弱度间伐,春季真菌数量和放线菌数量分别增加了2.11%、14.19%,细菌数量减少了61.05%;而夏季真菌和细菌数量分别减少了16.97%、27.94%,放线菌数量增加了26.32%;秋季弱度间伐下真菌、细菌、放线菌数量分别增加了80.52%、13.58%、75.00%;冬季分别减少了33.85%、42.37%、2.91%。
中度间伐,春季真菌和细菌数量分别减少了25.32%、36.14%,但放线菌数量增加了30.97%;夏季,3种类群微生物数量分别减少了18.18%、50.00%、19.14%;秋季,真菌、细菌数量分别减少了49.35%、9.88%,但放线菌数量增加63.79%;冬季,3种类群分别减少52.31%、53.39%、19.42%。
强度间伐,春季真菌、放线菌数量分别增加了77.64%、59.35%,细菌数量减少了42.11%;夏季,真菌和放线菌数量分别增加了9.70%、16.75%,细菌数量却减少了30.15%;秋季,真菌数量减少了44.16%,细菌和放线菌数量分别增加了44.44%、48.28%;冬季,真菌和细菌数量分别减少了29.23%、13.56%,放线菌数量增加了21.36%。
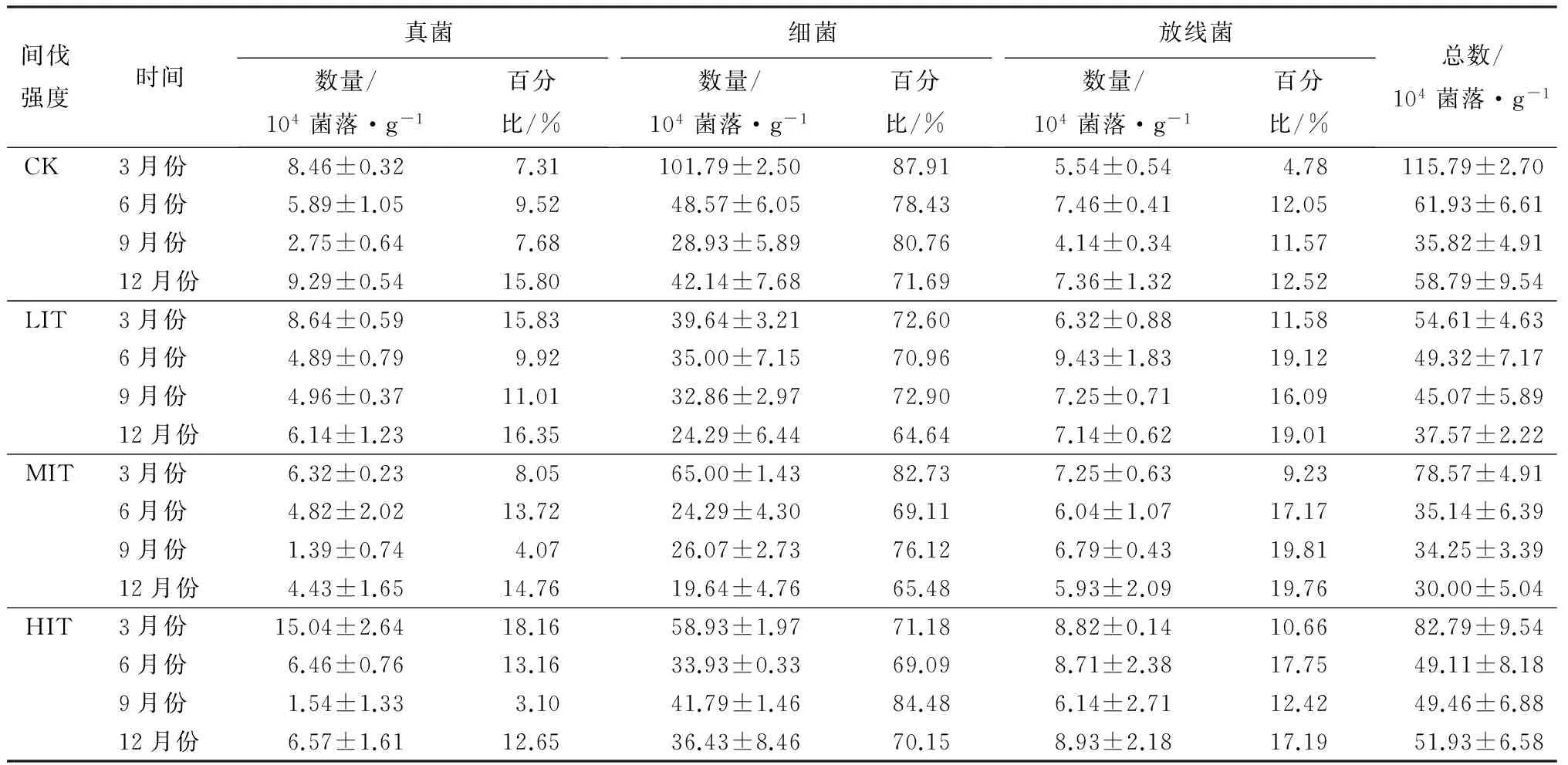
表2 不同间伐强度杉木林下凋落物中微生物数量
注:表中数据为平均值±标准差。
3.2间伐强度对微生物数量的季节性影响
由表3可知,凋落物中的真菌数量在3月份表现为,对照样地与弱度间伐间无明显差异,而与强度间伐及中度间伐之间差异分别达到显著(P<0.05)与极显著(P<0.01);在6月份及9月份真菌数量并不受间伐强度的影响;12月份,弱度间伐、强度间伐与对照样地之间真菌数量有显著差异(P<0.01),中度间伐与对照样地之间真菌数量有极显著差异(P<0.01),表明间伐对凋落物中真菌的数量在冬季有一定影响。
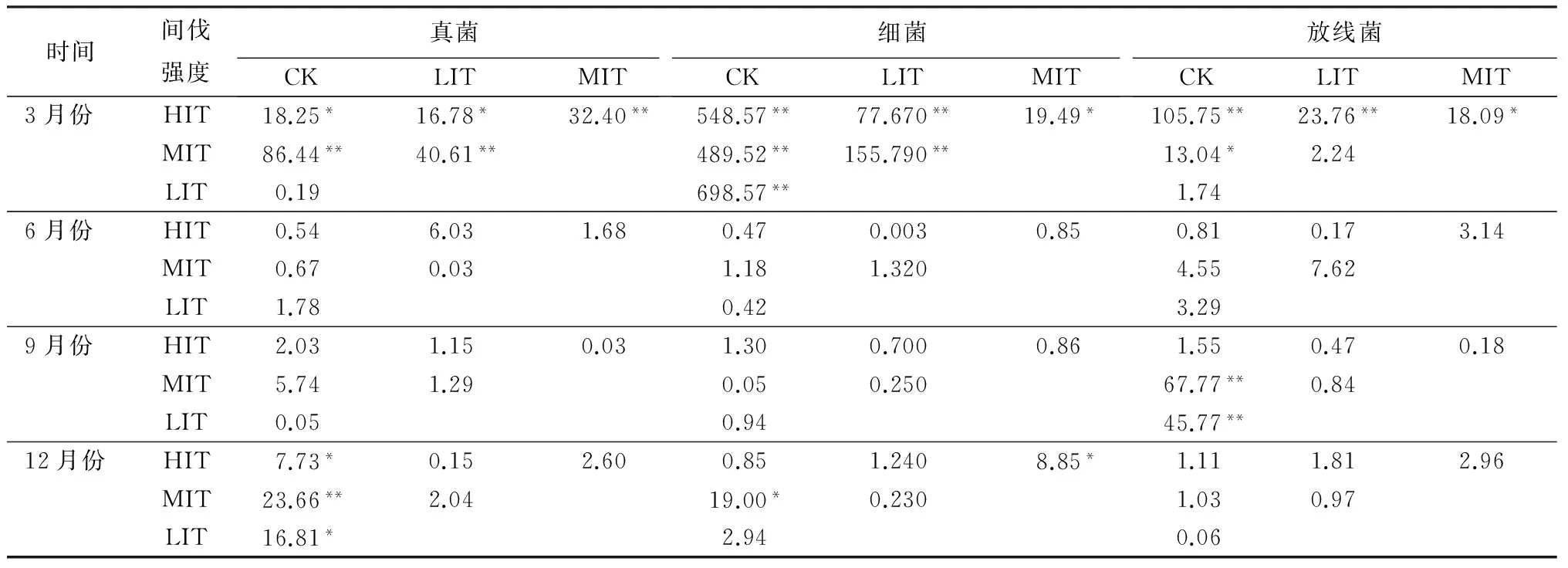
表3 不同时间、不同间伐强度下真菌、细菌、放线菌数量的差异性
注:*表示差异显著(P<0.05);** 表示差异极显著(P<0.01)。
3月份期间,间伐强度对凋落物中细菌数量的影响显著(P<0.05)。间伐强度对6月份和9月份凋落物中细菌数量无明显影响。而12月份的比较结果显示,中度间伐与对照、强度间伐之间均有显著差异(P<0.05),其余处理下差异不显著。
3月份,强度间伐与对照之间放线菌数量有极显著差异(P<0.01),中度间伐与对照结果之间也有显著差异(P<0.05)。6、12月份放线菌数量对间伐的强度响应均无显著性差异。9月份,弱度间伐、中度间伐与对照之间差异极显著(P<0.01),强度间伐对凋落物微生物中放线菌数量并无显著影响。
3.3温、湿度对微生物数量的影响
结合各样地下凋落物层温度变化(表4),对不同间伐强度下凋落物层微生物数量差异比较发现,与对照相比,3种间伐均能提高凋落物层月平均温约0.5 ℃,中度间伐对月最高温度提升作用最为显著。真菌的最适生长温度为22~28 ℃[20],6月份的最高温度均已超过真菌生长的最适温度,对真菌的数量生长有一定的抑制和阻碍作用,而9月份温度变化范围均为真菌适宜生长温度范围以内。中度间伐提升3月份最高温,降低12月份温度,所以显著减少真菌数量(P<0.01);同理,弱度、强度间伐在3、12月份显著影响真菌数量的(P<0.05)。适宜放线菌生长的温度为23~37 ℃[21],3月份中度、强度间伐以及9月份弱度、中度间伐内,凋落物层最高温有明显提升,对放线菌数量的增加影响显著(P<0.05)。

表4 不同间伐强度杉木人工林凋落物层均温及最高温年变化
由表5可知,在观测季节内,凋落物层年均相对湿度由大到小依次为弱度、对照、强度、中度。已有研究表明,相对湿度的增加有利于细菌数量的增长,过高的相对湿度不利于真菌数量的增长[22]。在本试验中,对照样地内细菌数量明显高于间伐处理样地,中度间伐下凋落物层细菌数量最少;真菌数量在强度间伐下最多,在中度间伐样地内最少;放线菌数量则无太大差异。
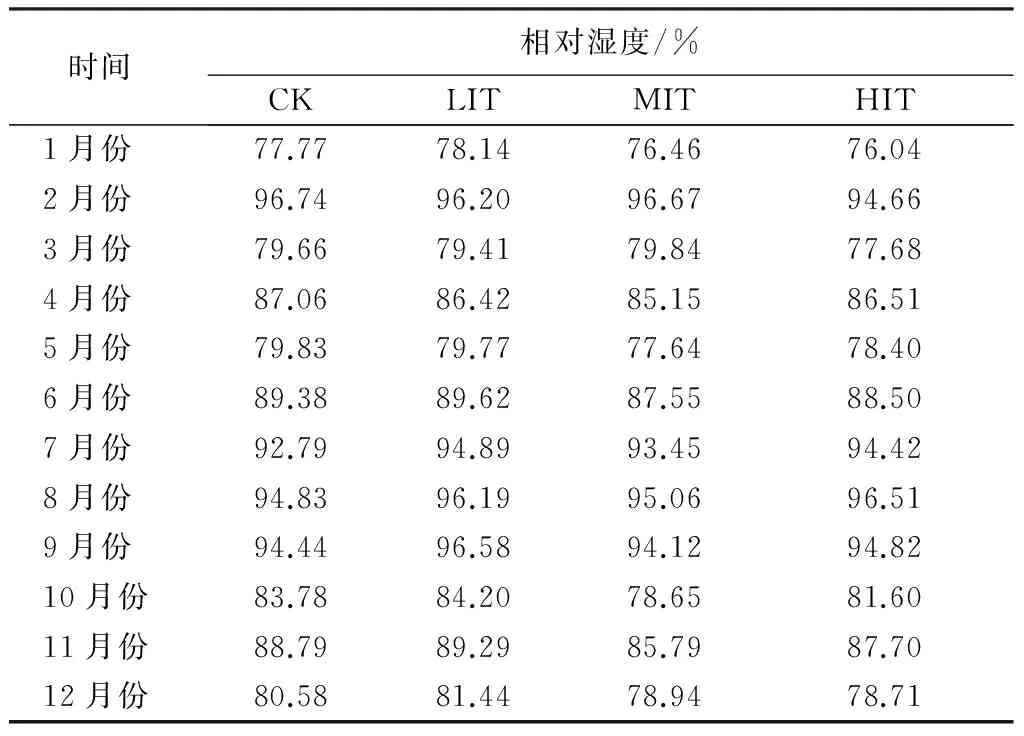
表5 不同间伐强度杉木人工林凋落物层相对湿度年变化
3.4林下植被多样性对微生物数量的影响
林下植被影响微生物数量的原因主要在于,间伐后林分空间及光照的改变会导致林下灌木层的多样性的增加[15],影响植被盖度,同时,在夜间对林地也具有一定的保温作用。从各指数大小来看(表6),强度间伐植被层多样性、丰富度及均匀度指数相对较低,弱度及中度间伐下植被层多样性、丰富度及均匀度指数相对较高。而凋落物层真菌数量在多样性指数相对较低的强度间伐下相对较多,在多样性指数较高的中度间伐下相对较少,表明植被盖度增加会减少真菌数量,林下植被盖度大的杉木林相对于植被盖度低的杉木林,真菌的数量有所下降[23]。放线菌数量也表现为在强度间伐下较多,而指数较高的样地放线菌的数量却相对较低。
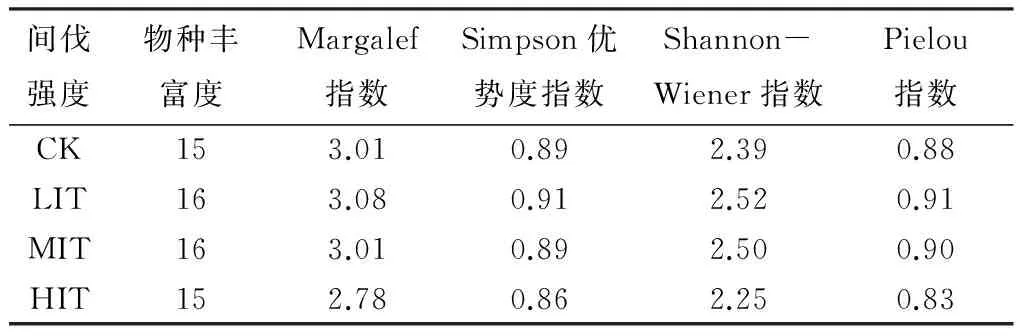
表6 不同间伐强度杉木人工林下植物物种多样性指数统计
3.5初始基质质量分数对微生物数量的影响
底物质量对微生物数量的增长也有一定影响,微生物数量与底物中氮质量分数相关,细菌对氮的需求相对较高。本研究中杉木凋落物中的底物初始值(表7)表示,对照样地中杉木凋落物具有更高的氮质量分数,3种间伐处理后会导致凋落物中的初始氮质量分数降低,间伐通过改变杉木林下的初始凋落物质量,而导致了细菌数量的改变。凋落物层微生物群落主要以细菌为主,放线菌和真菌处于竞争劣势,导致它们数量增加的困难。

表7 不同间伐杉木人工林下凋落物初始基质质量分数
注:表中数据为平均值±标准差;同列不同小写字母表示差异显著(P<0.05)。
3.6细菌、真菌和放线菌数量与环境因子的相关性
冗余分析可用环境因子来预测凋落物层微生物类群中数量变化分布(图1)。第1,2,3,4轴的特征值分别是0.480、0.140、0.069和0.204。第一轴解释了大部分的微生物数量的变异,微生物群落变化和环境的相关性值是0.937。经蒙特卡罗检验,冗余分析结果显示,微生物类群的数量显著受到凋落物层温度、植被层Margalef指数和Simpson优势度指数的影响;植被层Margalef指数、凋落物层温度和植被层Simpson优势度指数分别可以解释24.87%、23.64%、23.48%的微生物数量变化。从图1中还可得知,真菌数量与凋落物层湿度、植被层Simpson优势度指数和Pielou指数呈显著负相关。放线菌数量与植被层Margalef指数呈显著负相关。细菌数量则与凋落物基质初始氮质量分数呈正相关。
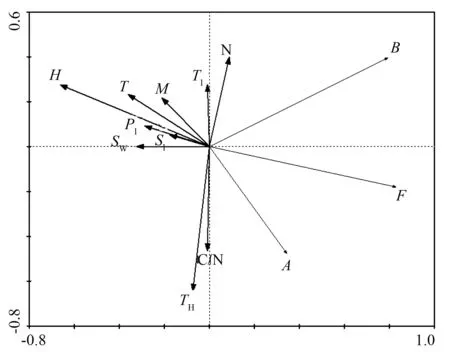
B.细菌数量;F.真菌数量;A.放线菌数量;T.凋落物层均温;T1.凋落物层最高温;H.凋落物层湿度;M.Margalef指数;SI.Simpson优势度指数;SW.Shannon-Wiener指数;PI.Pielou指数;N.凋落物初始氮质量分数;C/N.凋落物初始C质量分数/N质量分数值;TH.间伐强度。
图1凋落物层微生物数量与环境因子的冗余分析
4 结论与讨论
不同间伐强度下杉木凋落物中的微生物数量和主要类群数量随季节变化差异显著,微生物数量的增减并不由单一的原因所决定。不同时间和强度的间伐以及对应的变化会对凋落物产量造成影响[24],凋落物有利于异养微生物的生长、繁殖,使其数量明显增多[25]。本研究各环境因子中,以温度和丰富度指数对微生物数量的直接作用最大,其他因子影响较小,说明植被丰度和平均温度的相互作用对微生物数量的作用较大。微生物数量对间伐强度的响应在3月份最显著,原因在于温度的月动态是调控微生物数量的主要因素,7—9月份温度和湿度较高,利于微生物的繁殖,而3、12月份微生物活性较低,间伐造成温、湿度的差异较大,对微生物数量影响显著。针叶凋落物分解过程中微生物类群和数量相对较小,受环境变化的影响相对较大,3—5月份微生物数量明显下降,生长季节中期由于受针叶凋落物特征的限制,微生物数量相对稳定[12]。而过高湿度不利于真菌生长,极可能导致真菌数量的降低[22],这与本研究中真菌与凋落物层相对湿度呈极显著负相关相一致,由于林地中过高的湿度对微生物的生长及繁殖产生了一定的抑制作用,与水分含量存在显著的相关关系[26]。
微生物主要以植物残体为营养源,植物群落类型与质和量的差异必然会导致微生物在各植物群落中分布的不均一性[9]。细菌的数量和微生物总数与全氮质量分数呈极显著正相关。陈法霖等[27]的研究表明,微生物群落结构受凋落物初始C质量分数/N质量分数和木质素质量分数/N质量分数的调控,这一规律在本研究中并未体现,原因在于本研究仅对单一杉木凋落物进行研究,其不同间伐强度下的凋落物初始C质量分数/N质量分数和木质素质量分数/N质量分数值差异并不显著,若研究针阔混合凋落物分解,能对这一规律有更好的阐述。林下微生物群落活性和多样性特征受到林下小气候、养分和地上植被的直接和间接影响。间伐改变了林下植物群落的结构,为草本和灌木的生长提供了更多空间,改变了林下凋落物的质与量[16,18],对分解者提供更为有利的环境,促进凋落物的分解,使土壤肥力能够迅速恢复。同时,微生物分布和比例,既能反映环境因子的季节变化,也能反映生态系统功能强度的季节动态。
本研究表明,不同强度间伐杉木人工林下凋落物中微生物的数量随季节变化差异显著,微生物数量在春季期间对间伐响应最为显著,其受到凋落物层温度及林下植被丰富度的影响最为显著。
不同间伐强度对3大类群微生物的数量影响会导致凋落物向土壤输送的养分不同,本试验中,春季为中度间伐下微生物生长的旺盛期,应有利于凋落物的分解,对维持土壤肥力提高林分生产力具有重要意义,可为今后进一步深入探讨间伐对凋落物层微生物影响的具体作用机制提供一定的研究思路和科学依据。但凋落物层中微生物数量与土壤动物、分解速率等关系及间伐对微生物功能多样性影响等,有待于进一步深入研究。
[1]郭剑芬,杨玉盛,陈光水.森林凋落物分解研究进展[J].林业科学,2006,42(4):93-100.
[2]王勤,徐小牛,平田永二.不同坡位对琉球松人工林凋落物及其养分归还量的影响[J].应用生态学报,2004,15(7):1121-1125.
[3]TRINDER C J, JOHNSON D, ARTZ R R. Litter type, but not plant cover, regulates initial litter decomposition and fungal community structure in a recolonising cutover peatland[J]. Soil Biology and Biochemistry,2009,41(3):651-655.
[5]SAETRE P, BÅÅTH E. Spatial variation and patterns of soil microbial community structure in a mixed spruce-birch stand[J]. Soil Biology and Biochemistry,2000,32(7):909-917.
[6]周建斌,陈竹君,李生秀.土壤微生物量氮含量、矿化特性及其供氮作用[J].生态学报,2001,21(10):1718-1725.
[7]CHAPMAN S K, NEWMAN G S. Biodiversity at the plant-soil interface: microbial abundance and community structure respond to litter mixing[J]. Oecologia,2010,162(3):763-769.
[8]WANG C, GUO P, HAN G, et al. Effect of simulated acid rain on the litter decomposition ofQuercusacutissimaandPinusmassonianain forest soil microcosms and the relationship with soil enzyme activities[J]. Science of the Total Environment,2010,408(13):2706-2713.
[9]杨凯,朱教君,张金鑫,等.不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化[J].生态学报,2009,2(10):5500-5507.
[10]宋影,辜夕容,严海元,等.中亚热带马尾松林凋落物分解过程中的微生物与酶活性动态[J].环境科学,2014,35(3):1151-1158.
[11]严海元,辜夕容,申鸿.森林凋落物的微生物分解[J].生态学杂志,2010,29(9):1827-1835.
[12]周晓庆,吴福忠,杨万勤,等.高山森林凋落物分解过程中的微生物生物量动态[J].生态学报,2011,31(14):4144-4152.
[13]胡霞,吴宁,尹鹏,等.川西高原季节性雪被覆盖下凋落物输入对土壤微生物数量及生物量的影响[J].生态科学,2013,32(3):359-364.
[14]BARG A K, EDMONDS R L. Influence of partial cutting on site microclimate, soil nitrogen dynamics, and microbial biomass in Douglas-fir stands in western Washington[J]. Canadian Journal of Forest Research,1999,29(6):705-713.
[15]王祖华,李瑞霞,王晓杰,等.间伐对杉木人工林林下植被多样性及生物量的影响[J].生态环境学报,2010,19(12):2778-2782.
[16]BLANCO J A, IMBERT J B, CASTILLO F J. Nutrient return via litterfall in two contrastingPinussylvestrisforests in the Pyrenees under different thinning intensities[J]. Forest Ecology and Management,2008,256(11):1840-1852.
[17]HYNES H M, GERMIDA J J. Relationship between ammonia oxidizing bacteria and bioavailable nitrogen in harvested forest soils of central Alberta[J]. Soil Biology and Biochemistry,2012,46(1):18-25.
[18]SLODICAK M, NOVAK J, SKOVSGAARD J P. Wood production, litter fall and humus accumulation in a Czech thinning experiment in Norway spruce (Piceaabies(L.) Karst.)[J]. Forest Ecology and Management,2005,209(1):157-166.
[19]张鼎华,叶章发,范必有,等.抚育间伐对人工林土壤肥力的影响[J].应用生态学报,2001,12(5):672-676.
[20]毕国昌,郭秀珍,臧穆.在纯培养条件下温度对外生菌根真菌生长的影响[J].林业科学研究,1989,2(3):247-252.
[21]阿不都克里木·热依木,迪力夏提·托呼提.吐鲁番郊区22株放线菌最适生长条件和生长特性的初步研究[J].新疆师范大学学报(自然科学版),2001,20(2):36-38.
[22]贾夏,韩士杰,周玉梅,等.不同二氧化碳浓度条件下红松和长白赤松幼苗根际土壤微生物数量研究[J].应用生态学报,2005,16(7):1295-1298.
[23]焦如珍,杨承栋,孙启武,等.杉木人工林不同发育阶段土壤微生物数量及其生物量的变化[J].林业科学,2005,41(6):163-165.
[24]ROIG S, DEL RO M, CAELLAS I, et al. Litter fall in MediterraneanPinuspinasterAit. stands under different thinning regimes[J]. Forest Ecology and Management,2005,206(1):179-190.
[25]张明明,闫文德,梁小翠,等.不同处理对樟树林土壤微生物数量的影响[J].中南林业科技大学学报,2012,32(3):114-118.
[26]王启兰,曹广民,王长庭.高寒草甸不同植被土壤微生物数量及微生物生物量的特征[J].生态学杂志,2007,26(7):1002-1008.
[27]陈法霖,郑华,欧阳志云,等.土壤微生物群落结构对凋落物组成变化的响应[J].土壤学报,2011,48(3):603-611.
Seasonal Changes of Microbial Number of Forest Litter under Different Thinning Intensity of Chinese Fir//
Xiao Wenya, Fei Fei, Diao Jiaojiao, Guan Qingwei
(Nanjing Forestry University, Nanjing 210037, P. R. China)//Journal of Northeast Forestry University,2016,44(1):31-36.
Thinning intensity; Litter; Fungi; Bacteria; Actinomyces; Seasonal changes
肖文娅,女,1990年11月生,南京林业大学生物与环境学院、南京林业大学南方现代林业协同创新中心,博士研究生。E-mail:zizi0931@163.com。
关庆伟,南京林业大学生物与环境学院、南京林业大学南方现代林业协同创新中心,教授。E-mail:guanjapan999@163.com。
2015年9月19日。
S718.55
1)国家林业公益性行业科研专项 (201104075)、江苏高校协同创新计划资助项目 、江苏高校优势学科建设工程资助项目(PAPD)。
责任编辑:任俐。
猜你喜欢
林业科学研究(2022年2期)2022-03-25
河北果树(2021年4期)2021-12-02
中南林业科技大学学报(2021年3期)2021-03-23
——以杉木抚育间伐为例
绿色科技(2020年21期)2021-01-08
生态学报(2020年12期)2020-07-31
森林与环境学报(2020年4期)2020-07-27
中南林业科技大学学报(2020年2期)2020-02-12
水资源开发与管理(2019年11期)2019-12-02
天然产物研究与开发(2018年10期)2018-11-06
天然产物研究与开发(2014年7期)2014-04-27
