
亚热带不同植被恢复林地凋落物层碳、氮、磷化学计量特征
2020-02-12 09:03:52杜雨潭陈金磊李雷达刘兆丹苏元博
中南林业科技大学学报 2020年2期
杜雨潭,陈金磊,李雷达,辜 翔,刘兆丹,苏元博,方 晰,2
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004; 2.南方林业生态应用技术国家工程实验室,湖南 长沙 410004)
凋落物(也称枯落物)的分解过程是森林生态系统养分循环、能量流动的基本过程[1],是地上植被与土壤之间物质交换的“枢纽”,也是森林土壤肥力的自然来源之一[2],对森林生态系统的有机质储存和养分循环起着重要的作用[3]。因此,森林凋落物及其在地表形成的凋落物层是生态学、土壤学、生物地球化学的重要研究内容之一。凋落物层是地上植物生长发育过程中产生并归还到地表形成的死有机物质,由未分解、半分解及已分解的凋落物组成,是森林生态系统的重要组成部分,其种类、贮量和数量上的消长反映着群落间的差别和动态特征[4]。
1876年,德国学者E.Ebermayer 阐述了森林凋落物在养分循环中的重要性。随后,许多学者对森林凋落物进行了大量的研究[5-6]。20 世纪60年代初,国内开始开展凋落物的研究,20 世纪80年代有较大进展,凋落物的积累和分解被认为是控制生态系统能量流动与功能结构的重要因素,研究主要集中在凋落物的年凋落量、组成及其动态变化[7],凋落物的分解速率及其养分释放规律[8-9]等方面。然而,有关林地地表凋落物层的研究,特别是凋落物层现存量大多是因为其是生态系统生物量的组成部分,才会在研究生态系统生物量时被提及,而且大多研究却忽视了不同植物群落由于组成树种的差异,凋落物产量及其形成的凋落物层现存量可能会有较大的差异[10-11]。近年来,随着人们对森林生态系统凋落物研究的深入,也逐渐关注凋落物层现存量及其与其他生态因子(如土壤[12]、气象因子[13]、动物[14]等)的关系,但也主要集中在亚热带的针叶林,而有关阔叶林、针阔混交林的研究报道仍较少[15]。此外,近年来的C、N、P化学计量特征研究也主要集中在植物的鲜叶[16-17], 不同演替阶段植物群落土壤[18-20]两方面,有关地表凋落物层C、N、P 化学计量特征的研究报道仍很少[21-22]。
中国亚热带地区分布着大面积的常绿阔叶林,是全球同纬度地带上的“绿洲”。但由于长期受到人类干扰,常绿阔叶林遭到了严重破坏,生物多样性下降,生态系统功能衰退。为了保护生物多样性和恢复常绿阔叶林的生态服务功能,近20多年来,中国政府实施了天然林保护、退耕还林和防护林体系等建设一系列林业生态工程,该地区植被恢复迅速,但由于人为干扰强度不同,形成了处于不同恢复阶段的多种次生植物群落。在亚热带特定的气候条件和土壤类型下,森林皆伐后,从次生裸地开始,植被恢复的通常模式是:经2 ~3 a 的自然恢复,形成以草本植物为主,伴随少量灌木的灌草丛;约再经10 a 恢复为以檵木Loropetalum chinense、南烛Vaccinium bracteatum、杜鹃Rhododendron mariesii、 白 栎Quercus fabri、茅栗Castanea seguinii等灌木为主的灌木林;随后约30年出现一些阳性先锋乔木树种,如马尾松Pinus massoniana针叶树种和枫香Liquidambar formosana、 麻 栎Quercus acutissima、 南 酸 枣Choerospondias axillaries、 拟 赤 杨Alniphyllum fortunei等落叶阔叶树种,形成马尾松针叶林,马尾松针阔混交林或落叶常绿阔叶林;随后恢复演替为以壳斗科Fagaceae(如柯Lithocarpus glaber、青冈Cyclobalanopsis glauca)、山茶科Theaceae(如木荷Schima superba、红淡比Cleyera japonica)和樟科Lauraceae(如樟树Cinnamomum camphora、椤木石楠Photinia davidsoniae、毛豹皮樟Litsea coreana)等耐荫性常绿阔叶树种为优势树种的常绿阔叶林[23-25]。因此沿着亚热带森林恢复演替梯度,根据植被恢复程度及其树种组成和结构,这些次生植物群落可以划分为灌草丛、灌木林、针阔混交林、落叶常绿阔叶混交林和常绿阔叶林[26]。森林类型更替是影响生态系统有机质循环的重要因素,它对森林生态系统的生产力、碳吸存和养分保持功能有影响。然而有关中亚热带凋落物层现存量及其C、N、P 随着植被恢复的动态研究鲜有报道,对正确了解与认识随着植被恢复,凋落物层及各分解层现存量如何演变,土壤N、P 含量增加,凋落物层及其各分解层凋落物C/N、C/P、N/P 比值是否会相应的变化,凋落物层在生态系统物质循环过程中的作用是否增强等问题仍不十分清楚。因此,本研究采用空间差异代替时间序列的方法,在湘中丘陵区选取处于不同植被恢复阶段的4种林地(灌草丛、灌木丛、针阔混交次生林和常绿阔叶林)为作为一个恢复系列,比较研究不同恢复阶段林地凋落物层现存量及其C、N、P 化学计量的变化特征,为亚热带地区退化林地的植被恢复和管理提供科学依据。
1 研究地概况
研究地设置在湖南省中东部的长沙县(113°17′ ~113°27′E,28°23′ ~28°24′N), 属于典型低山丘陵区,地形起伏较大,坡度多在20°~30°,海拔约为55 ~350 m;为亚热带东南季风湿润气候,年平均气温17.0 ℃,极端高温为40 ℃(7—8月份),极端低温为-11 ℃(1月份),平均无霜期216~269 d,雨量充沛,相对湿度较大,年降水量1 412 ~1 559 mm,主要集中于4—7月份,年均日照1 440 h。土壤以板岩和页岩发育而成的酸性红壤为主,地带性植被为亚热带常绿阔叶林,由于人为干扰(采伐、火烧、放牧)频繁,强度各异,原生地带性森林植被破坏比较严重。20 世纪90年代初开始实施封山育林,形成了多个处于不同恢复演替阶段的植物群落(灌草丛、灌木林、针阔混交林、常绿阔叶林),以及以杉木Cunninghamia lanceolata人工林、马尾松人工林、湿地松Pinus elliottii人工林为主的人工林群落,为开展中亚热带森林植被恢复演替研究提供了良好的场所。
2 研究方法
2.1 样地设置和群落调查
2015年10月,采用空间代替时间的研究方法,按照亚热带森林群落自然演替进程,根据人为干扰程度、植被恢复程度及其群落树种组成,选取地域毗邻、环境条件(立地、土壤、气候)基本一致,处于不同植被恢复阶段的4种植物群落:恢复早期阶段的檵木Loropetalum chinensis-南烛Vaccinium bracteatum-杜鹃Rhododendron mariesii灌草丛(LVR),恢复中期I 阶段的檵木-杉木Cunninghamia lanceolata-白栎Quercus fabri灌木林(LCQ),恢复中期II 阶段的马尾松-柯(又名石栎,Lithocarpus glaber)-檵木针阔混交林(PLL),恢复后期(亚顶级)阶段的柯-红淡比Adinandra millettii- 青 冈Cyclobalanopsis glauca常绿阔叶林(LAG)。4 个不同恢复演替阶段植物群落的基本概况如下:
1)LVR:天然常绿阔叶林采伐后,1966年春炼山、人工整地,营造马尾松人工林,无施肥历史,1990年皆伐马尾松人工林。1991年以来,不断砍杂、放牧,2012年停止人为干扰,当前自然恢复为檵木+南烛(Vaccinium bracteatum)+杜鹃灌草丛,草本植物多且生长较好,伴随许多幼小灌木。
2)LCQ:天然常绿阔叶林采伐后,1966年春炼山、人工整地,营造杉木人工林,无施肥历史,1989-1990年皆伐杉木人工林。之后每隔3 ~5 a砍伐一次,2004年停止砍伐,当前自然恢复为檵木+杉木+白栎灌木林,灌木发育良好,但没有形成明显的乔木层,草本植物稀少。
3)PLL:20 世纪60年代末,天然常绿阔叶林采伐后自然恢复为马尾松+柯+檵木针阔混交林(PLL),林龄约45 ~50 a,林下幼苗、幼树丰富,植株密度较大,但大径级植株比例不高[27]。
4)LAG:长期以来无明显人为干扰,保存比较完好的柯+红淡比+青冈常绿阔叶林(LAG),群落结构相对稳定,林龄约80 ~90 a。
根据不同恢复阶段植物群落物种组成及其结构的复杂程度,分别设置3 ~4 个长期观测固定样地。LVR 群落设置4 个20 m×20 m 样地,LCQ群落设置3 个20 m×20 m 样地,PLL、LAG 群落均设置3 个30 m×30 m 样地。
采用样方调查法,2016年10—11月落叶树种落叶前完成群落调查,所有植物均鉴定到种。4种群落的基本特征及其主要树种组成如表1 所示。
2.2 凋落物层现存量的测定和分析样品采集、 处理
于2015年12月,2016年4月、6月、10月每一块固定样地分别沿对角线均匀设置3 个1 m× 1 m 样方,根据凋落物层的分解层分层标准[15],测定样方内未分解层、半分解层、已分解层凋落物的湿重后,分别采集各分解层凋落物的分析样品(约1.0 kg),将分析样品置于80 ℃恒温烘干至恒质量,测定分析样品的含水率。根据分析样品的含水率,计算各样方不同分解层凋落物的干质量,估算不同植被恢复阶段各分解层凋落物及整个凋落物层的现存量。取4 次采样的平均值作为最终测定结果。
凋落物层分析样品烘干称干质量后,经植物粉碎机磨碎,过60 目筛孔保存于样品瓶中,用于测定有机碳(C)、全氮(N)、全磷(P)含量。
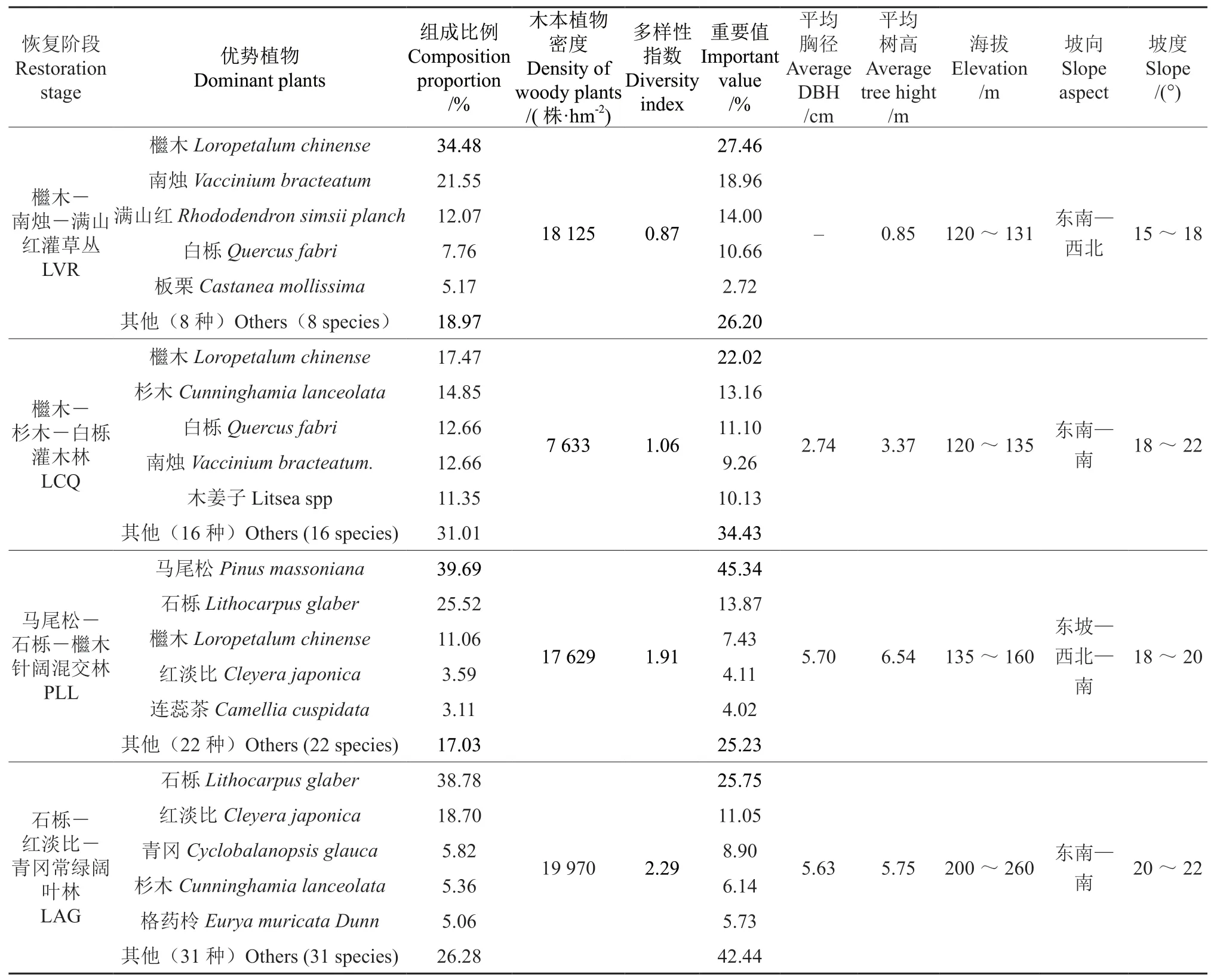
表1 不同植被恢复阶段样地的基本特征†Table 1 Stand characteristics at different vegetation restoration stages
2.3 凋落物层样品中有机碳、全氮、全磷含量的分析方法
凋落物层分析样品中C 含量用重铬酸钾-浓硫酸容量法测定,N 含量用K9840 自动凯氏定氮仪(山东海能科学仪器有限公司,山东济南)测定,P 含量用碱熔-钼锑抗比色法测定。每个分析样品平行测定2 次,取平均值作为该样品的测定结果。取4 次采样的平均值作为最终测定结果。
2.4 数据统计与处理
凋落物层及各分解层C、N、P 储量用以下公式估算:
DL=WL×CL
式中:DL为凋落物层或各分解层凋落物的C(或N、P)储量(kg/hm2),WL为凋落物层或各分解层凋落物的现存量(kg/hm2),CL为凋落物层或各分解层凋落物的C(或N、P)含量(g/kg)。
用Excel 2016 统计各项指标的平均值和标准偏差。用SPSS16.0 软件包中的单因素方差分析(One-way ANOVA)的的最小显著差数法(LSD,P<0.05)分析不同恢复阶段林地凋落物层现存量,凋落物层C、N、P 含量、储量及其化学计量比各项指标的差异显著性。图、表中的数据用“平均值±标准偏差”表示。运用SigmaPlot 12.5 软件制图。
3 结果与分析
3.1 不同植被恢复阶段林地凋落物层的现存量
如表2 所示,凋落物层现存量随着植被恢复呈增加趋势,最高是最低的4.97 倍,且LVR 与LCQ、PLL、LAG 差 异 显 著(P<0.05), 而LCQ、PLL、LAG 两两之间差异不显著(P>0.05)。同一分解层凋落物现存量总体上也随着植被恢复而增加。在未分解层,PLL 最高,LVR 最低,且LVR 与LCQ、PLL、LAG 之 间,LCQ 与PLL 之间差异显著(P<0.05),而LCQ、PLL 与LAG差异不显著(P>0.05);在半分解层、已分解层,LAG 最高,LVR 最低,且LVR 与LCQ、PLL、LAG 差异显著(P<0.05),LCQ、PLL、LAG两两之间差异不显著(P>0.05)。同一林地,不同分解层凋落物现存量均表现为:已分解层>半分解层>未分解层,不同分解层之间的差异随着植被恢复而增大。其中,LVR 不同分解层之间差异不显著(P>0.05),LCQ 未分解层、半分解层与已分解层差异显著(P<0.05),但未分解层与半分解层差异不显著(P>0.05),PLL 未分解层与已分解层差异显著(P<0.05),半分解层与未分解层、已分解层差异不显著(P>0.05);LAG 不同分解层两两之间差异显著(P<0.05)。未分解层凋落物现存量占凋落物层现存量的百分比最低,其次为半分解层,而已分解层最高,表明未分解层凋落物已被分解为半分解和已分解的凋落物。

表2 不同恢复阶段林地凋落物层现存量† Table 2 Floor litter mass in different restoration stages kg·hm-2
3.2 凋落物层C、N、P 含量和储量
3.2.1 C、N、P 的含量
如图1 所示,凋落物层以及未分解层、半分解层、已分解层平均C 含量分别为:277.20 ~ 424.10、371.71 ~502.73、288.52 ~453.19、171.36 ~316.38 g/kg,均以PLL 为最高,LCQ 为最低,且LVR 与PLL 差异显著(P<0.05),但与LCQ、LAG 差异不显著(P>0.05),LCQ、PLL、LAG 两两之间差异显著(P<0.05)。同一林地,C 含量均随着凋落物的分解而下降,其中LVR、LCQ、PLL 不同分解层两两之间差异显著 (P<0.05),LAG 未分解层与半分解层、已分解层差异显著(P<0.05),但半分解层与已分解层差异不显著(P>0.05)。
凋落物层及各分解层N 平均含量总体上随着植被恢复而增加,且LAG 与LVR、LCQ、PLL(除已分解层外)差异显著(P<0.05),而LVR、LCQ、PLL 两两之间差异不显著(P>0.05)。同一林地N 含量随着凋落物的分解而下降,除PLL外,其他3 种林地不同分解层两两之间差异显著 (P<0.05)(图1)。
从图1 可以看出,凋落物层及各分解层P 平均含量总体上随着植被恢复而增加,且LVR 与LAG 差异显著(P<0.05),与LCQ(除未分解层外)、PLL(除已分解层外)差异不显著(P>0.05),LCQ(除已分解层外)、PLL(除未分解层外)与LAG 差异不显著(P>0.05)。LVR、LCQ 各分解层凋落物P 含量随着凋落物的分解而下降,而PLL 半分解层P 含量最高,未分解层最低,LAG 依次为:半分解层>未分解层>已分解 层,LVR、PLL、LAG 不同分解层之间差异不显著 (P>0.05),LCQ 未分解层与已分解层差异显著(P<0.05),但半分解层与未分解层、已分解层差异不显著(P>0.05)。
3.2.2 C、N、P 的储量
从表3 可以看出,PLL 凋落物层及其各分解层C 储量最高,其次是LAG,LVR 最低,且PLL与LVR、LCQ、LAG(除半分解层、未分解层外)差异显著(P<0.05),LAG 与LVR、LCQ(除未分解层、已分解层外)差异显著(P<0.05),LCQ 与LVR 差异显著(P<0.05)。同一林地不同分解层凋落物C 储量两两之间的差异均不显著(P>0.05)。
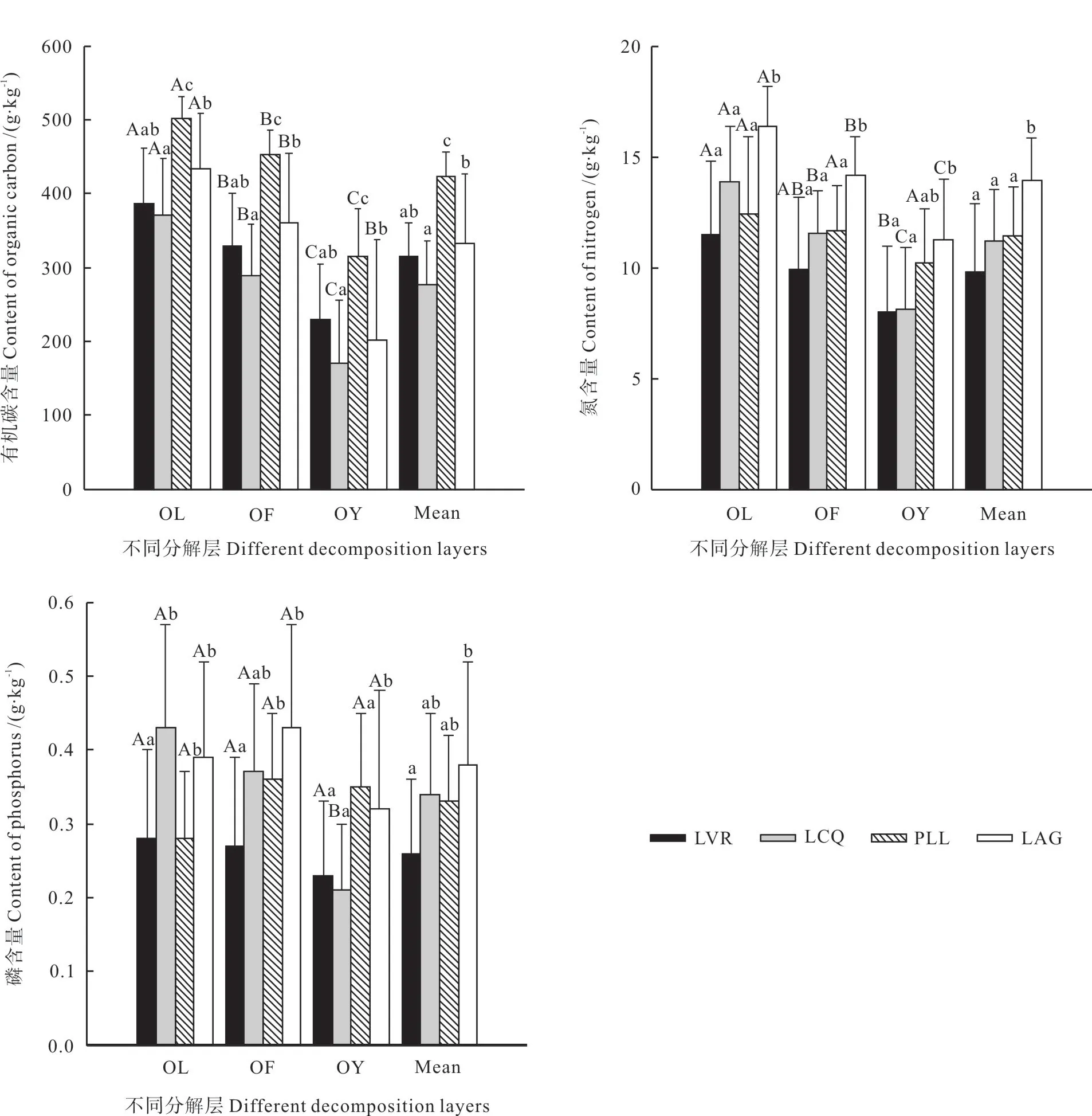
图1 不同恢复阶段林地凋落物层碳、氮、磷含量Fig.1 Concentrations of carbon, nitrogen and phosphorus in litter layer at different restoration stages
凋落物层及各分解层N 储量均随着植被恢复而增加,且LVR 与LCQ、PLL、LAG 差异显著 (P<0.05),LCQ、PLL(除未分解层、半分解层外)与LAG 差异显著(P<0.05),LCQ 与PLL 差异不显著(P>0.05)。同一林地,N 储量均表现为:已分解层>半分解层>未分解层,LAG 未分解层与半分解层、已分解层差异显著(P<0.05),但半分解层与已分解层差异不显著(P>0.05),其余3 种林地不同分解层两两之间差异不显著 (P>0.05)(表3)。
如表3 所示,凋落物层及各分解层P 储量总体上随着植被恢复而增加,且LVR 与LCQ、PLL、LAG 差异显著(P<0.05),LCQ 与PLL(除未分解层、半分解层外)、LAG差异显著(P<0.05),但PLL 与LAG 差异不显著(P>0.05)。同一林地,P 储量也表现为:已分解层>半分解层>未分解层,LVR、LCQ 不同分解层两两之间差异不显著(P>0.05),PLL、LAG 未分解层与半分解层、已分解层差异显著(P<0.05),半分解层与已分解层差异不显著(P>0.05)。
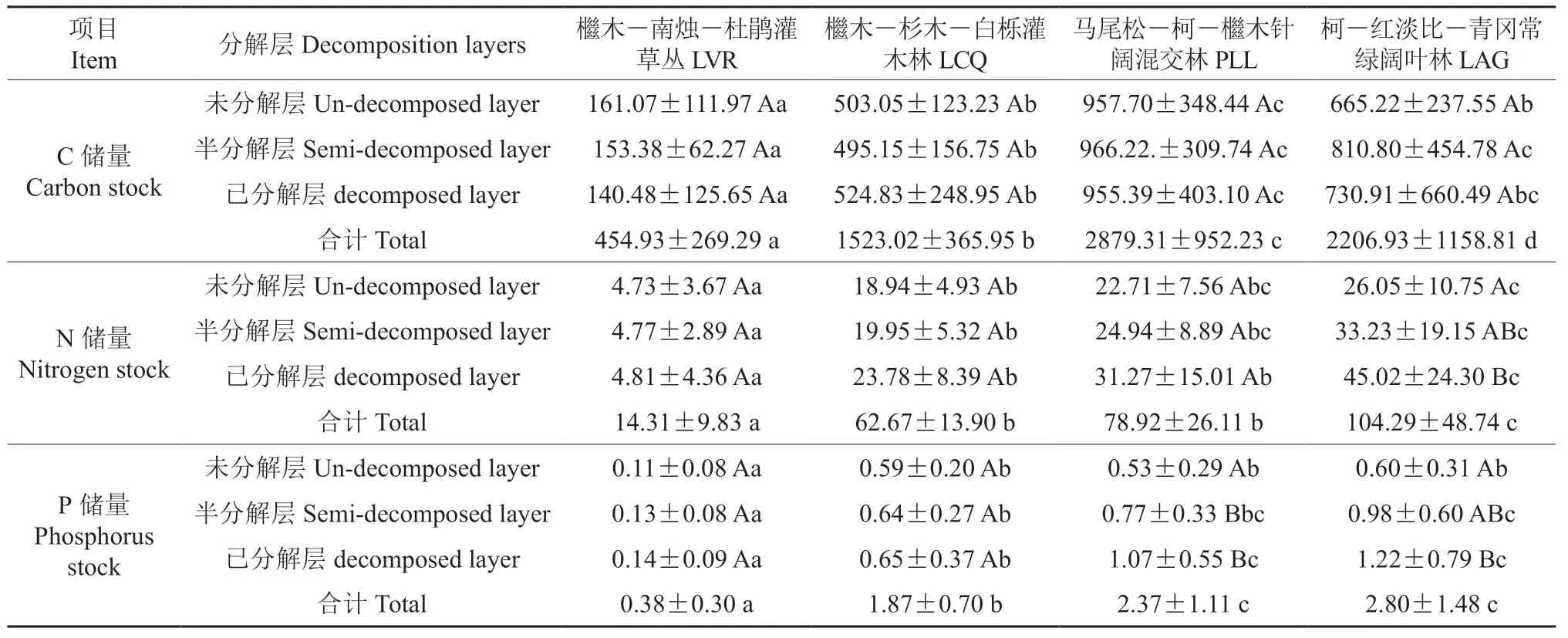
表3 不同恢复阶段林地凋落物层C,N,P 储量 Table 3 Carbon, nitrogen and phosphorus storage of litter layers at different restoration stages kg·hm-2
3.3 凋落物层C、N、P 化学计量比
从图2 可以看出,凋落物层及各分解层C/N比值均表现为:PLL >LVR >LCQ >LAG,且LVR、PLL 与LCQ、LAG 差异显著(P<0.05),但LVR 与PLL 之间,LCQ 与LAG 之间差异不显著(P>0.05)。同一林地,C/N 比值总体上随着凋落物的分解而下降,与C 含量的变化趋势基本一致,LVR、LCQ 不同分解层两两之间差异不显著(P>0.05),PLL、LAG 已分解层与未分解层、半分解层差异显著(P<0.05),而未分解层与半分解层差异不显著(P>0.05)。
如图2 所示,凋落物层、半分解层、已分解层C/P 比值均以LVR 最大,其次为PLL,LCQ 或LAG 最小,且LVR 与LCQ(除已分解层外)、LAG 差异显著(P<0.05),与PLL 差异不显著 (P>0.05),LCQ、PLL、LAG 两两之间差异不显著(P>0.05)。未分解层C/P 比值为:PLL >LVR >LAG >LCQ,且PLL 与LCQ、LAG 差异显著(P<0.05),与LVR 差异不显著(P>0.05),LVR 与LCQ 差异显著(P<0.05),而与LAG差异不显著(P>0.05),LCQ 与LAG 差异不显著(P>0.05)。同一林地,C/P 比值随着凋落物的分解而下降,LVR、LCQ 不同分解层两两之间差异不显著(P>0.05),PLL未分解层与半分解层、已分解层差异显著(P<0.05),而半分解层与已分解层差异不显著(P>0.05);LAG 未分解层与已分解层差异显著(P<0.05),而半分解层与未分解层、已分解层差异不显著(P>0.05)。
凋落物层、半分解层N/P 比值为:LVR > LAG >PLL >LCQ,未分解层为:PLL >LVR > LAG >LCQ,已分解层为:LCQ >LAG >LVR > PLL,但凋落物层及各分解层不同林地两两之间差异均不显著(P>0.05)。同一林地,LVR、PLL的N/P 比值随着凋落物的分解而下降,LCQ、LAG 呈先下降再升高,但4种林地不同分解层之间(除PLL 外)差异不显著(P>0.05)(图2)。
4 讨 论
4.1 植物恢复对凋落物层现存量的影响
研究表明,群落凋落物层现存量主要取决于两个因素,即与凋落物量呈正相关,与分解速率呈负相关[28]。因而凋落物层现存量受到生物和非生物多个因素的共同影响[29]。研究表明,同一气候条件下,不同植物群落由于组成树种不同,凋落物产量及其分解速率的差异,使得不同植物群落凋落物层及其各分解层凋落物现存量不同[10-11,30]。本研究中,凋落物层以及各分解层凋落物现存量总体上随着植被恢复而逐渐增加,与张庆费等、郑路等的研究结果基本一致[32,15]。进一步分析表明,凋落物层现存量与群落植物多样性指数(表1)呈极显著正相关关系(r=0.720,P<0.05,n=13),表明随着植被恢复,树种组成增多,植物多样性提高是凋落物层现存量增加的主要影响因子之一。
本研究中,不同恢复阶段林地同一分解层现存量占凋落物层现存量的百分比不同,未分解层为20.08%~27.30%,半分解层为27.67%~31.11%,已分解层为41.59%~50.72%,表明不同林地凋落物层的分解强度不同。究其原因可能是:一方面,随着植被恢复,物种种类增加且物种组成不断改变,大型树木比例增加(表1),新陈代谢加快,凋落物量增加[31];另一方面,不同恢复阶段群落内微环境(光照、温度、湿度等)不同,凋落物的分解速率不同,地表凋落物层现存量也不同。
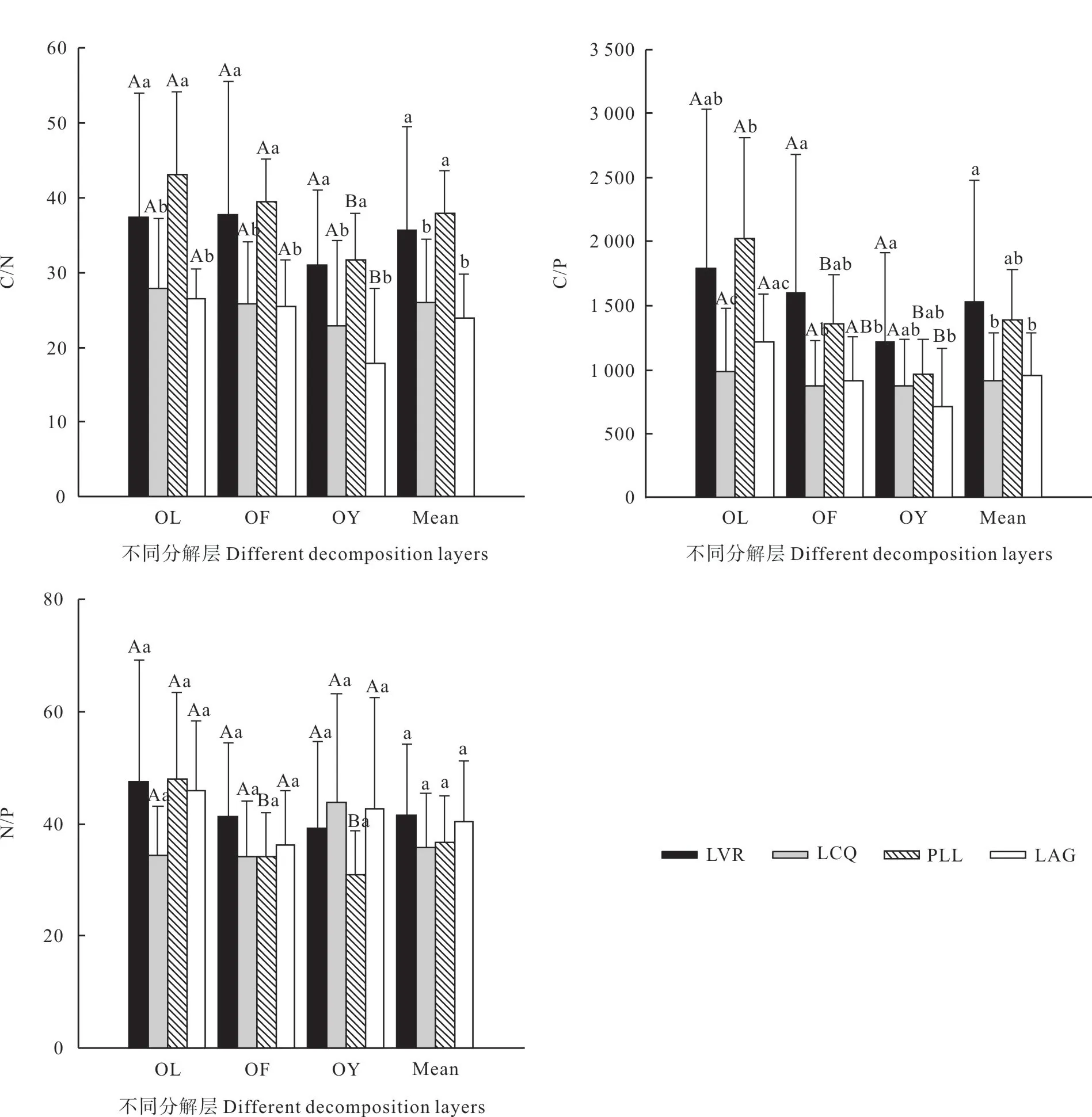
图2 不同恢复阶段林地凋落物碳、氮、磷化学计量比Fig.2 Carbon, nitrogen, phosphorus stoichiometry ratios of litter layer at different different restoration stages
研究表明,未分解层凋落物现存量除了与群落的凋落物量有关外,还与外界环境条件、凋落物特性有关[32]。本研究中,由于PLL 以马尾松为优势树种(表1),凋落物以马尾松针叶为主,凋落物层C/N 比值保持较高(图2),分解速率较低,未分解层凋落物积累多,而LAG 凋落物以阔叶树叶为主(表1),C/N 比值较低(图2),分解速率较高,未分解层凋落物被分解为半分解层和已分解层凋落物而积累较少,导致PLL 未分解层凋落物现存量高于LAG,而半分解层和已分解层凋落物现存量低于LAG,甚至已分解层凋落物现存量低于LCQ,表明随着植被恢复,LAG 凋落物能迅速分解归还养分,更有利于土壤养分的保持与积累。
凋落物分解是一个复杂而又漫长的过程,包括物理、化学和微生物3 种分解方式[33]。分解初期,由于水溶性物质和碳水化合物的快速淋失与降解,凋落物分解速率较高,随着凋落物的分解,纤维素、木质素等难以降解的物质不断积累,使得分解中期或后期凋落物分解速率下降[34],导致半分解或已分解凋落物的积累。本研究中,同一林地,不同分解层凋落物现存量随着凋落物的分解而增加,占凋落物层现存量的百分比增大,可能与不同分解阶段凋落物的分解速率有较大关系[35]。表明未分解层凋落物向半分解层和已分解层迁移,有机质向土壤归还,有利于土壤肥力的增加[15],且随着植被恢复,这种迁移能力逐渐增强,说明随着植被恢复,凋落物分解速率会提高[21],土壤微生物活动增强,被分解的凋落物越来越多,未分解层凋落物现存量越来越少。
4.2 植被恢复对凋落物层C、N、P 含量及其化学计量比的影响
由于针叶树具有特殊的养分获取方式,其针叶C 含量较阔叶树高1.6%~3.4%[36];相反,针叶树针叶N,P 含量普遍低于阔叶树[37]。本研究中,PLL 以马尾松为优势树种,凋落物以马尾松针叶为主,而LVR 以小灌木和草本植物为主,LCQ 以落叶灌木为主,LAG 以阔叶树为主,因而PLL 未分解层凋落物C 含量显著高于LCQ、LAG;相反,PLL 未分解层凋落物N,P 含量低于LCQ、LAG,这也是PLL 未分解层凋落物C/N、C/P 比值显著高于LCQ、LAG 的原因。研究表明,凋落物C 含量随着凋落物的分解呈显著单调下降[38]。本研究中,4种不同恢复阶段林地凋落物层C 含量随着凋落物的分解而下降。但与多种林分类型相比,马尾松林凋落物层C 含量随着凋落物分解下降速率最小[39-40]。因此,PLL 半分解层、已分解层凋落物C 含量仍显著高于LCQ、LAG,而PLL半分解层、已分解层N、P 含量仅略高于LCQ,仍低于LAG,使得PLL 半分解层、已分解层凋落物C/N、C/P 比值仍高于LCQ、LAG,特别是C/N比值显著高于LCQ、LAG。
本研究中,C/N 比值基本上随着植被恢复呈下降趋势,与马文济等的研究结果一致[21]。也由于C、N 含量随着凋落物分解而下降,但C 含量下降幅度比N 含量更为明显,因此,C/N 比值随着凋落物的分解而下降,与C 含量的变化趋势一致。由于未分解层凋落物C/N、C/P 比值较高时,凋落物分解过程中出现N、P 富集现象或慢缓释放,从而导致半分解层、已分解层凋落物N、P 含量仍维持较高的水平,C/P 比值大体上随着凋落物的分解而下降。但由于不同恢复阶段林地凋落物N、P 含量及其随着凋落物分解的变化程度不同,因此,N/P比值随着凋落物分解没有呈现一致的变化趋势。
本研究中,4种不同植被恢复阶段林地未分解层凋落物N、P 含量分别为11.53 ~16.39、0.28 ~0.43 g/kg,与全球衰老叶片的N、P 含量[41]相比,P 含量较低,而N 含量较高,与全球木本植物凋落叶N、P 含量(10.9、0.85 g/kg)[42]相比,也呈现高N 含量,低P 含量。此外,4种林地未分解层凋落物的C/N 比值(26.57 ~43.18)与长白山次生针阔混交林凋落物初始的C/N 比值(31 ~70)接近,而C/P(984.03 ~2 026.60)、N/P(34.38 ~47.88)比值显著高于长白山次生针阔混交林凋落物初始的C/P(381 ~876)和N/P(8.6 ~20.0)比值[38],表明研究区植物生长发育明显受到P 的限制,也可能是由于高纬度和低温的影响,北方森林土壤养分(N,P)相对丰富,森林凋落物养分(N,P)含量高于低纬度地区[43];此外,不同地区植物群落类型及其组成树种的差异,可能也是不同地区凋落物层C,N,P 含量及化学计量比不同的原因。
4.3 植被恢复对凋落物层C、N、P 储量的影响
凋落物层及各分解层C、N、P 储量主要取决于凋落物层及各分解层的现存量和C、N、P的含量。由于凋落物层及各分解层凋落物现存量和N、P 含量基本上随着植被恢复而增加,而PLL 凋落物层C 含量显著高于LAG,因此,C 储量以PLL 为最高,其次是LAG,LVR 最低,而N、P 储量随着植被恢复而增加。也由于凋落物层N、P 含量随着植被恢复的变化幅度比凋落物层现存量小,凋落物层N、P 储量主要取决于凋落物层现存量,因而N、P 储量的变化趋势与凋落物层现存量基本一致。不管是凋落物层现存量还是C、N、P 储量随着植被恢复呈增加趋势,且养分储量向腐殖质层转移增强,因此促进植被恢复可明显提高林地养分的蓄存量。
在植被恢复过程中,地上植物组成不断改变,凋落物组成和数量也随之发生变化,因此本研究的数据和结论只能说明当前的研究结果,要准确地反映植被恢复对凋落物层现存量及其C、N、P化学计量的影响还需要长期定位研究,深入揭示植物组成的变化对凋落物层现存量的影响;此外,凋落物分解和养分释放受到多种因素(如土壤微生物、动物以及林下微环境的改变等)的综合影响,因此不同植被恢复阶段凋落物分解过程中,各因素及其综合的驱动机制还有待进一步研究。
5 结 论
随着植被恢复,凋落物层及其各分解层现存量增加,未分解层凋落物向半分解层和已分解层的迁移能力逐渐增强;PLL 凋落物层及其各分解层C 含量、储量最高,其次是LAG;随着植被恢复,凋落物层及各分解层N、P 含量、储量总体上呈增加趋势,而C/N、C/P、N/P 比值总体上呈下降趋势,有利于凋落物的分解及其养分释放。表明植被恢复促进了生态系统养分生物循环和自我维持机制的形成,有利于土壤养分的保持与积累,体现了生态系统物质循环随着植被恢复逐渐优化。
猜你喜欢
河北地质(2023年1期)2023-06-15 02:45:12
戏曲研究(2022年2期)2022-10-24 01:54:12
矿产勘查(2020年4期)2020-12-28 00:30:08
水资源开发与管理(2019年11期)2019-12-02 09:09:16
河北林业科技(2016年5期)2016-11-08 03:12:58
河北林业科技(2016年3期)2016-10-27 08:02:44
河北林业科技(2015年3期)2015-12-17 06:31:42
断块油气田(2014年5期)2014-03-11 15:33:45
中国质量与标准导报(2014年6期)2014-02-28 22:24:03
河南科技(2014年16期)2014-02-27 14:13:25
