
蔗糖和光周期在泸定百合试管鳞茎膨大中的作用机制
2016-08-15 05:25周玲云高素萍陈锋
浙江大学学报(农业与生命科学版) 2016年4期
周玲云, 高素萍, 陈锋
(四川农业大学风景园林学院,成都 611130)

蔗糖和光周期在泸定百合试管鳞茎膨大中的作用机制
周玲云, 高素萍*, 陈锋
(四川农业大学风景园林学院,成都 611130)
为研究蔗糖和光周期对泸定百合试管鳞茎膨大的影响及其交互作用,解释两者在鳞茎膨大过程中的作用机制,以泸定百合种球鳞片为外植体诱导形成的无菌丛芽为材料,采用不同质量浓度(30、60、90 g/L)蔗糖与不同光周期(0、8、12、16、24 h/d)的组合处理,测定鳞茎发生和膨大的形态指标及其内源性糖含量与相关酶活性的变化。结果显示:1)在蔗糖质量浓度为60 g/L、光周期为8 h/d组合处理下,鳞茎膨大效果最好,与全黑暗处理间无显著差异。2)在30 g/L蔗糖和各光照时间处理下,蔗糖含量变化趋势为W型,果糖和葡萄糖含量变化趋势为M型;而在60和90 g/L蔗糖处理下,蔗糖含量呈先下降后升高再下降趋势,果糖和葡萄糖含量变化趋势与蔗糖含量变化趋势相反。3)在不同质量浓度蔗糖处理下,蔗糖磷酸合成酶活性的变化趋势与蔗糖含量变化趋势基本一致,而蔗糖合成酶活性的变化趋势与蔗糖含量变化趋势相反。综上,蔗糖质量浓度和光周期对试管小鳞茎的发生和膨大存在交互作用,但蔗糖对百合鳞茎生长与膨大影响更大,是造成鳞茎诱导及膨大的主要因子;蔗糖合成酶参与蔗糖分解代谢过程,而蔗糖磷酸合成酶参与蔗糖积累过程;蔗糖不仅在膨大过程中扮演了生理角色,还可能扮演了糖信号角色,并且这些过程都受到严格的浓度调控.在生产中,推荐的最适组合为蔗糖质量浓度60 g/L,光周期0 h/d。
泸定百合; 蔗糖; 光周期; 鳞茎; 膨大; 作用机制
Summary Lily has high ornamental and economic value. However, the reproduction rate of traditional propagation method was low, and was easily infected, causing degradation of bulbs and associated problems. Application of tissue culture techniques can greatly improve the reproduction coefficient in the production process, but the weakness of plantlets and low survival rate in the transplanting process have become a prominent problem during the commercial process of lily bulbs. Hence, to cultivate strong seedling and increase survival rate, we could promote the formation and enlargement of bulbs byinvitroculture. This study aimed to explore the influence of sucrose and photoperiod on the enlargement ofinvitrobulblet ofLiliumsargentiaeand their interaction effect, and to explain their action mechanism in the process ofinvitrobulblet enlargement.
Sterile buds induced by bulblet scales ofL.sargentiaeas explants were selected as materials. The experiments were conducted under three different concentrations of sucrose (30, 60 and 90 g/L) and five different photoperiods (0, 8, 12, 16 and 24 h/d). The morphological index, endogenous sugar and related enzymatic activity changes were measured in formation and enlargement ofinvitrobulblet.
The results showed that: 1) The optimal condition for bulblet enlargement was sucrose concentration of 60 g/L and photoperiod of 8 h/d. At a certain concentration of sucrose, there was no significant difference between the photoperiods of 8 h/d and 0 h/d. 2) Under the sucrose concentration of 30 g/L and different lighting conditions, the trend was W-type for sucrose content, and M-type for both fructose and glucose contents; under the sucrose concentrations of 60 and 90 g/L, the trend of sucrose content was fall-rise-fall, while the trend of fructose and glucose contents was rise-fall-rise, opposite to that of sucrose content. 3) Among treatments with different sucrose concentrations, the change trend of sucrose phosphate synthase activity was basically the same with that of the sucrose content, but the change trends of the sucrose synthase activity and sucrose content were opposite.
In conclusion, 60 g/L sucrose in complete darkness can promote bulblet enlargement in production. The sucrose synthase is involved in decomposition process of sucrose, while the sucrose phosphate synthase is involved in the accumulation process of sucrose. Sucrose not only plays a physiological role, but also may play a sugar signal role in the bulblet enlargement process, both of which are strictly regulated by sugar concentration. Sucrose concentration and photoperiod have interaction effect on the induction and enlargement ofinvitrobulblets, but the effect of sucrose is greater. Thus, sucrose is a major factor for the formation and enlargement ofinvitrobulblets.
泸定百合(Liliumsargentiae)为百合科百合属球根类植物[1],拥有极高的园林观赏价值和经济价值。传统的分球繁殖方法存在繁殖系数低、易感染病毒从而造成种球退化等问题。在生产中,采用组织培养技术能极大地提高繁殖系数,但试管苗生长较弱,在移栽过程中成活率低,这已成为我国百合种球在工厂化生产中遇到的突出问题;因此,在试管内结鳞茎并使其膨大,可达到壮苗和提高移栽成活率的目的。而关于百合鳞茎膨大的机制目前尚不清楚。前人已作了关于环境因子及植物生长调节剂等对百合鳞茎膨大的影响研究[2-5];同时,在植物生长发育过程中糖类作为碳源、能源、渗透保护剂和重要的信号分子[6],对植物地下部诱导及鳞茎膨大也具有一定的效果,但关于蔗糖在其中扮演的角色和作用机制尚不明确,有待于进一步探究。此外,有研究表明,光周期有利于马铃薯[7-8]和魔芋[9]等经济作物地下茎的形成与膨大。SLIMMON等[8]对马铃薯试管苗和张延龙等[10]对卷丹百合试管苗鳞茎研究均表明,在8 h光照下诱导比在全黑暗条件下对地下茎形成与膨大的影响更为显著。然而,光周期与蔗糖共同处理对鳞茎诱导及膨大的影响及交互作用尚缺乏系统研究。因此,本研究以泸定百合种球鳞片为外植体诱导形成的无菌丛芽为材料,研究不同质量浓度蔗糖与不同光周期组合处理对百合鳞茎发生和膨大的影响及其内源性糖与相关酶的变化,旨在为生产中人工调控百合种球膨大寻求适宜的环境条件和糖类供给,解决试管苗移栽成活率低的问题,为生产优质泸定百合种球提供技术支撑和实践指导。
1 材料与方法
1.1材料
以泸定百合种球鳞片为外植体,以生长在MS+1.00 mg/L 6-苄氨基嘌呤(6-benzylaminopurine,6-BA)+0.15 mg/LN-乙酰天门冬氨酸(N-aceytl aspartate,NAA)+7 g/L琼脂+30 g/L蔗糖培养基中的健康、正常的无菌丛芽体为试验材料。
1.2组培方法与试验设计
将诱导形成的丛芽体接种于继代培养基上进行扩增,丛芽增殖到所需的数量后,在无菌条件下将丛芽分割成单个芽,剪去上部叶片,接种到MS+1.00 mg/L 6-BA+0.15 mg/L NAA+7 g/L琼脂+不同质量浓度(30、60、90 g/L)蔗糖处理的培养基上,光照时间分别为每天0、8、12、16和24 h。采用2因素完全随机处理(表1),每处理重复5次。以30 g/L蔗糖处理为对照。培养条件为温度(23±2) ℃,光照强度1 500~2 000 lx,相对湿度(55±5)%。

表1 蔗糖和光周期复合处理组合
1.3蔗糖和光周期在诱导膨大过程中形态指标的测定
在未进行培养基更换情况下,培养55 d后,取出诱导的小鳞茎,剪去叶片和根,擦净培养基后称鲜样质量。用游标卡尺测量小鳞茎的直径,统计叶长及根长,每水平统计15株,重复3次,取平均值。生长量用生长指数(表示鲜质量增加的倍数)和结鳞茎率(表示鲜鳞茎增加的倍数)表示。
生长指数=(鳞茎鲜质量-单芽初质量)/
单芽初质量×100%。
结鳞茎率=结鳞茎数/总数×100%。
1.4蔗糖和光周期在诱导膨大过程中内源性糖含量和相关酶活性变化的测定
1.4.1内源性糖含量变化
选择蔗糖、果糖、葡萄糖3种内源性糖含量变化作为测定指标。在初培养第15天时开始取样测定,之后每隔10 d测定1次,到第55天时结束。蔗糖、果糖测定参照张志良等[11]的方法,葡萄糖测定按照葡萄糖测定试剂盒说明书进行(葡萄糖氧化酶-过氧化物酶法)。重复测定3次,取平均值。
1.4.2蔗糖相关酶活性变化
鳞茎蔗糖合成酶(sucrose synthase,SS)和蔗糖磷酸合成酶(sucrose phosphate synthase,SPS)活性测定参考孙红梅等[12]的方法并做适当修订。精密称取0.5 g样品鲜样,加入5 mL提取介质[100 mmol/L Tris-HCl(pH 7.5),10 mmol /L MgCl2,1 mmol/L乙二胺四乙酸二钠,10 mmol/L二硫苏糖醇,2%乙二醇]于研钵中磨成匀浆后,倒入离心管,在低温冷冻超速离心机上以1.2×104r/min离心10 min。以上提取过程均在4 ℃下进行。取上清液进行SS活性测定:加入0.4 mL合成反应介质[100 mmoL/L Tris-HCl(pH 7.5),10 mmoL/L MgCl2,5 mmoL/L 尿苷二磷酸葡萄糖,5 mmoL/LD-果糖],测定时间与1.4.1节相同,对照加0.1 mL 2 moL/L NaOH。SPS活性测定只需将D-果糖换成100 mmol/L 6-磷酸果糖。酶活性单位用μg/(g·h)表示。
1.5数据统计分析
用Excel 2015制作图表。用SPSS 20.0进行数据分析,其中:用2因素方差分析检验处理间的显著性;用最小显著差别法进行处理间多重比较;用皮尔逊法评价相关显著性。
2 结果与分析
2.1蔗糖和光周期对鳞茎诱导和膨大的形态影响
总体上,不同蔗糖质量浓度和光周期组合处理对泸定百合试管丛芽体鳞茎的分化及其生长膨大影响显著(表2)。在同一光周期下,蔗糖质量浓度从30 g/L(CK)提高至60 g/L时,鳞茎的膨大效果持续增强,在60 g/L时达到顶峰;随着蔗糖质量浓度继续提高,鳞茎的膨大效果开始衰退;在蔗糖质量浓度为90 g/L时,膨大效果减弱至30 g/L处理的水平。在相同蔗糖质量浓度下,光周期为8 h/d处理后的鳞茎诱导及膨大效果(直径、鲜质量、生长指数、结鳞茎率)最好,但与0 h/d处理间差异无统计学意义(P>0.05)。通过主效应和交互作用分析可知,蔗糖与光周期存在显著的交互作用,但更多地表现为协同作用,而蔗糖是本试验鳞茎形成及膨大的主要贡献者。在生产上,可将60 g/L蔗糖、0 h/d光周期作为鳞茎诱导及膨大的最佳组合选择。
2.2蔗糖和光周期对鳞茎诱导及膨大过程中内源性糖含量的影响
在形态指标测定基础上,本试验希望从植物生理反应的层面解释泸定百合试管鳞茎膨大的机制。从图1可以看出,在泸定百合鳞茎诱导及膨大过程中,蔗糖和光周期对鳞茎内源性糖类含量具有显著的影响。在15~55 d培养期间内,在30 g/L蔗糖处理下,蔗糖含量随着光周期的延长表现为下降-升高-下降-升高的W型变化,果糖和葡萄糖含量的变化趋势为升高-下降-升高-下降的M型变化;而在60和90 g/L蔗糖处理下,蔗糖含量呈先下降后升高再下降趋势,果糖和葡萄糖含量呈先升高后下降再升高趋势。由此可以看出,果糖和葡萄糖含量的变化趋势与蔗糖正好相反,蔗糖含量降低是由于蔗糖水解形成葡萄糖和果糖,造成葡萄糖和果糖含量上升。这一结果进一步说明了蔗糖与葡萄糖、果糖的上下游关系。
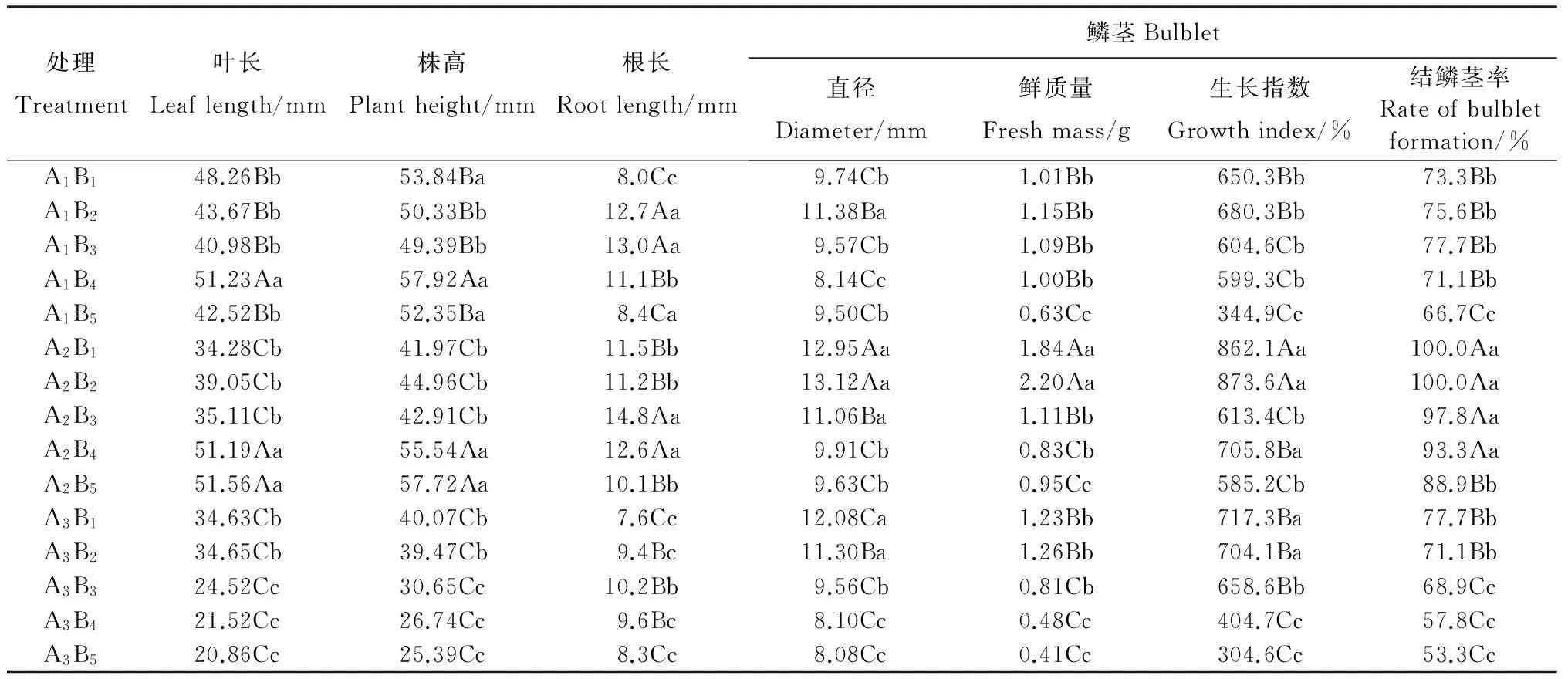
表2 不同组合处理对泸定百合试管鳞茎形成及膨大的影响
各处理表示的含义详见表1。同列数据后的不同大写字母表示不同质量浓度蔗糖处理组间在P<0.05水平差异有统计学意义,不同小写字母表示不同光周期处理组间在P<0.05水平差异有统计学意义。
Please see Table 1 for details of each treatment. The values within a column followed by different capital or lowercase letters show statistically significant differences among the treatments of different sucrose concentrations or photoperiods at the 0.05 probability level, respectively.
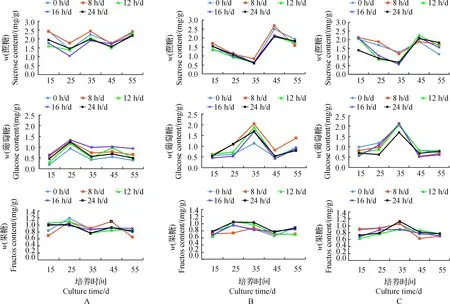
A:30 g/L 蔗糖处理;B:60 g/L蔗糖处理;C:90 g/L蔗糖处理.A: Treatment with 30 g/L sucrose; B: Treatment with 60 g/L sucrose; C: Treatment with 90 g/L sucrose.图1 在蔗糖和光周期处理下鳞茎在膨大过程中内源性糖含量的变化Fig.1 Change of the endogenous sugar content in the process of bulblet enlargement under sucrose and photoperiod treatments
从图1还可以看出:在30 g/L蔗糖处理下,在25 d时蔗糖含量出现最小值,葡萄糖和果糖呈现最大值,最高约有50%的蔗糖分解;与30 g/L蔗糖处理相比,60和90 g/L蔗糖处理在35 d时蔗糖含量出现最小值,这期间造成了60%~70%的蔗糖水解,使得葡萄糖含量迅速增加,高达2.0 mg/g。这一现象说明不同质量浓度蔗糖处理表现为其分解转化时间上的先后;总体上,高质量浓度蔗糖处理的分解转化时间比低质量浓度处理滞后。
2因素方差及主效应分析表明:60与30 g/L蔗糖处理对内源性糖类含量的影响在统计学上有显著差异,蔗糖质量浓度和光周期2因素间具有交互作用,但蔗糖是造成鳞茎诱导及膨大过程中内源性糖类含量变化的主要因子。
2.3蔗糖和光周期对鳞茎诱导及膨大过程中相关酶活性的影响
为了解释在蔗糖和光周期处理过程中内源性糖含量变化的原因,本试验测定了蔗糖磷酸酶(SPS)和蔗糖合成酶(SS)的活性变化,以进一步说明鳞茎诱导膨大的原因。总体上,随着蔗糖质量浓度的增加和光周期的延长,泸定百合鳞茎在诱导及膨大过程中SPS和SS活性变化趋势明显(图2)。在30 g/L蔗糖处理下,SPS活性呈现为下降-升高-下降-升高的W型变化,SS活性表现为升高-下降-升高-下降的M型变化;在60和90 g/L蔗糖处理下,SPS活性表现为先下降后升高的整体变化趋势,而SS活性表现为先升高后下降再略微上升的整体变化趋势。由以上结果可知,SS与SPS的变化趋势相反,且SS活性的变化趋势与蔗糖含量的变化趋势相反,而SPS活性的变化趋势与蔗糖含量的变化趋势基本一致:说明这2个酶分别负责蔗糖的分解与合成。

A:30 g/L蔗糖处理;B:60 g/L蔗糖处理;C:90 g/L蔗糖处理.A: Treatment with 30 g/L sucrose; B: Treatment with 60 g/L sucrose; C: Treatment with 90 g/L sucrose.图2 在蔗糖和光周期处理下鳞茎在膨大过程中蔗糖磷酸酶(SPS)和蔗糖合成酶(SS)活性变化Fig.2 Change of sucrose phosphate synthase (SPS) and sucrose synthase (SS) activities in the process of bulblet enlargement under sucrose and photoperiod treatments
3 讨论与结论
高质量浓度蔗糖具有刺激器官形成的作用。本研究结果表明:在光周期一定的情况下,蔗糖质量浓度为60 g/L时,新增鳞茎率和结鳞茎率最高,生长指数、鲜质量、直径也都优于其他质量浓度蔗糖处理。王爱勤等[13]对新铁炮1号百合的研究以及朱旭东等[14]对东方百合的研究均表明,适当提高蔗糖质量浓度对百合鳞茎诱导及膨大有显著作用,本研究结果与其相同。但对本试验而言,当蔗糖质量浓度达到90 g/L时,鳞茎的生长指数、鲜质量、直径和结鳞茎率都低于60 g/L的处理效果,甚至有些指标低于30 g/L的处理效果。该现象说明,当蔗糖质量浓度高于60 g/L时延迟和制约了鳞茎的诱导及膨大,蔗糖质量浓度越高对鳞茎的限制越大,甚至会演变为抑制。说明蔗糖质量浓度升高对地下茎膨大的有效性是有限制条件的,当高于临界质量浓度时,这种作用表现为抑制效果。
在蔗糖质量浓度一定的情况下,通过5种光照处理对泸定百合试管鳞茎诱导形成的影响进行综合分析可知:在0和8 h/d光照条件下产生的试管鳞茎数量多而大,新增鳞茎率和结鳞茎率高,两者均有利于试管鳞茎的形成;而在8 h/d光照处理下形成的试管鳞茎的平均质量和直径最大,是形成试管鳞茎的最佳光照处理,但与0 h/d光照处理下的效果差异无统计学意义(P>0.05);在16和24 h/d光照处理下形成的试管鳞茎数量少,平均鳞茎质量和直径都较小,对试管鳞茎形成不利。造成这种现象的原因可能是8 h/d处理有一定时间的光照,有较大的“源”,因而形成的试管鳞茎大;16和24 h/d光照处理虽有较大的“源”,但植株在生长中进行了更多的形态建成,包括叶片和根系的生长,从而消耗了鳞茎内的营养元素,最后形成了较小的试管鳞茎,反而不利于鳞茎的膨大。这表明光周期与蔗糖质量浓度间存在显著的交互作用。在外源糖质量浓度一定的情况下,短日照(小于8 h/d)有利于试管鳞茎诱导与膨大,而在生产上全黑暗培养更有利于降低成本。
百合鳞茎的主要营养组成是糖类[15],内源性糖变化对鳞茎形成及膨大具有重要意义。植物产生大量的蔗糖、果糖、葡萄糖等可溶性糖,这些糖类物质的增加不仅可以提供植物生长发育所需的能量物质,而且可以维持细胞渗透压平衡[16-17]。在本研究中蔗糖对鳞茎内源性糖类含量变化具有显著影响的结果也证实了这一结论。低质量浓度与高质量浓度蔗糖处理在各种光照周期条件下内源性糖含量呈现不同的变化趋势,且在相同蔗糖质量浓度、不同光周期处理下表现出相似的内源性糖模式,而高质量浓度蔗糖处理与低质量浓度蔗糖处理所表现出的内源性糖模式不一致。说明在此过程中糖类代谢被严格调控,而添加的外源性蔗糖是影响其变化的主要因子。
植物代谢包括糖类分子的产生和消失。蔗糖分子的产生、分解和消失与蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)息息相关。前人的研究成果已经表明蔗糖代谢改变了植物的生长发育。如:拟南芥SPS的表达导致在“库”中蔗糖水平升高和总干质量增加[18];相反,烟草植物SPS的表达下调导致烟草萎黄,光合作用减弱和生长减缓[19];在马铃薯中,SS在蔗糖代谢过程中具有重要作用,随着SS水平的升高,马铃薯块茎中含有较多的淀粉[20]。因此,为了进一步解释内源性糖代谢的变化,有必要对蔗糖关键酶变化进行深入研究。本试验结果表明,在各质量浓度蔗糖处理下,SPS变化趋势与蔗糖含量的变化趋势基本一致,而SS变化趋势与蔗糖含量的变化趋势相反。说明SS参与蔗糖分解代谢过程,而SPS参与蔗糖积累过程。孟衡玲等[21]研究认为,蔗糖合成酶是调控蔗糖代谢的关键酶之一,该酶的主要功能是在蔗糖的降解方向催化蔗糖进入各种代谢途径,本研究结果与之相一致。蔗糖不但可以作为植物生长发育所必需的碳源、能源或渗透调剂等[22],而且其本身可能还具有信号分子的功能[23-24]。说明蔗糖不仅在膨大过程中扮演了生理角色,还可能扮演了糖信号角色,即通过操纵外源性蔗糖的变化,激发蔗糖感应和信号传导,影响植物体内SS与SPS活性的变化,从而影响植物内源性蔗糖及蔗糖衍生的代谢物水平,从而影响百合鳞茎的生长和膨大。
总之,光周期和蔗糖均对泸定百合鳞茎形成及膨大有显著的影响,且两者存在交互作用,但主要是通过高质量浓度蔗糖引起百合鳞茎内源性糖含量及相关酶活性的变化;SS参与了蔗糖分解代谢过程,而SPS参与了蔗糖积累过程;蔗糖不仅在膨大过程中扮演了生理角色,还可能扮演了糖信号角色,并且这些过程都受到严格的浓度调控。在生产上推荐的最适组合为蔗糖质量浓度60 g/L,光周期0 h/d。
[1]中国科学院中国植物志编辑委员会.中国植物志:14卷.北京:科学出版社,1980:127.
China Flora Editorial Board of CAS.FloraofChina: Vol.14. Beijing: Science Press, 1980:127. (in Chinese)
[2]王晓丽,韩立群,刘杰,等.活性炭和多效唑对垂花百合试管鳞茎膨大的影响.安徽农业科学,2011,39(19):11429-11430.
WANG X L, HAN L Q, LIU J,etal. Effects of AC and PP333on the development of the tube bulbs ofLiliumcernuum.JournalofAnhuiAgriculturalSciences, 2011,39(19):11429-11430. (in Chinese with English abstract)
[3]廉美兰,朴炫春,孙丹.影响百合试管鳞茎诱导及膨大的几种因素.延边大学农学学报,2006,28(3):153-158.
LIAN M L, PIAO X C, SUN D. Several factors affecting bulblet formation and enlargement ofLiliuminvitro.JournalofAgriculturalScienceYanbianUniversity, 2006,28(3):153-158. (in Chinese with English abstract)
[4]郑一强,孙红梅.东方百合试管鳞茎形成条件优化.中国农学通报,2011,27(6):90-94.
ZHENG Y Q, SUN H M. Culture condition optimization of test tube-bulb formation of lily-orientals.ChineseAgriculturalScienceBulletin, 2011,27(6):90-94. (in Chinese with English abstract)
[5]朱志国.影响百合试管鳞茎增殖因素的研究.热带作物学报,2013,34(10):1961-1965.
ZHU Z G. Affecting factors on the proliferation of test-tube lily bulb.ChineseJournalofTropicalCrops, 2013,34(10):1961-1965. (in Chinese with English abstract)
[6]SMEEKENS S, MA J K, HANSON J,etal. Sugar signals and molecular networks controlling plant growth.CurrentOpinioninPlantBiology, 2010,13:273-278.
[7]霍凤兰,栾清业,尹玉花.蔗糖质量浓度和光照对马铃薯试管薯诱导的影响.甘肃农业科技,2009(11):3-5.
HUO F L, LUAN Q Y, YIN Y H. Effect of sucrose concentration and light on microtuber induction ofSolanumtuberousumL.GansuAgriculturalScienceandTechnology, 2009(11):3-5. (in Chinese with English abstract)
[8]SLIMMON T, MACHADO S V, COFFIN R. The effect of light oninvitromicrotuberization of potato cultivars.AmericanJournalofPotatoResearch, 1989,66(12):843-848.
[9]胡建斌.魔芋离体形态发生机制及其繁殖技术.武汉:华中农业大学,2006:66-74.
HU J B. Mechanism ofinvitromorphogenesis of konjac (AmorphophallusBlume) and the techniques for its propagation. Wuhan: Huazhong Agricultural University, 2006:66-74. (in Chinese with English abstract)
[10]张延龙,张启翔,薛晓娜.光周期对野生卷丹试管苗鳞茎形成及糖代谢的影响.园艺学报,2010,37(6):957-962.
ZHANG Y L, ZHANG Q X, XUE X N. The effects of the photoperiods on the bulblet formation and sugar metabolism change of wildLiliumlancifoliuminvitro.ActaHorticulturaeSinica, 2010,37(6):957-962. (in Chinese with English abstract)
[11]张志良,瞿伟菁,李小方.植物生理学实验指导.北京:高等教育出版社,2009:106-108.
ZHANG Z L, QU W J, LI X F.ExperimentGuidanceofPlantPhysiology. Beijing: Higher Education Press, 2009:106-108. (in Chinese)
[12]孙红梅,王微微,何玲,等.百合鳞茎蔗糖合成酶活性检测体系的建立.沈阳农业大学学报,201l,42(3):285-290.
SUN H M, WANG W W, HE L,etal. Establishment of detection system for sucrose synthase activity in lily bulb.JournalofShenyangAgriculturalUniversity, 2011,42(3): 285-290. (in Chinese with English abstract)
[13]王爱勤,周歧伟,何龙飞,等.百合试管结鳞茎的研究.广西农业大学学报,1998,17(1):71-75.
WANG A Q, ZHOU Q W, HE L F,etal. Studies on the bulblet formation in tube ofLiliumlongiflorumL.JournalofGuangxiAgriculturalUniversity, 1998,17(1):71-75. (in Chinese with English abstract)
[14]朱旭东,田松青,储海霞.东方百合试管成球和种球低温处理的研究.江苏农业科学,2005(5):74-76.
ZHU X D, TIAN S Q, CHU H X. Studies on growth and cold treatment of test-tube bulblet ofLilium.JiangsuAgriculturalSciences, 2005(5):74-76. (in Chinese with English abstract)
[15]李玉帆,明军,王良桂,等.百合基本营养成分和活性物质研究进展.中国蔬菜,2012(24):7-13.
LI Y F, MING J, WANG L G,etal. Research progress on basic nutritional and bioactive substances of lily. ChinaVegetables, 2012(24):7-13. (in Chinese with English abstract)
[16]PARTELLI F L, VIEIRA H D, RODRIGUES A P D. Cold induced changes on sugar contents and respiratory enzyme activities in coffee genotypes.CiênciaRural, 2010,40(4):781-786.
[17]TURHAN E, ERGIN S. Soluble sugars and sucrose-metabolizing enzymes related to cold acclimation of sweet cherry cultivars grafted on different rootstocks.TheScientificWorldJournal, 2012:979682.
[18]PARK J Y, CANAM T, KANG K Y,etal. Over-expression of anArabidopsisfamily A sucrose phosphate synthase (SPS) gene alters plant growth and fibre development.TransgenicResearch, 2008,17(2):181-192.
[19]CHEN S, HAJIREZAEI M, PEISKER M,etal. Decreased sucrose-6-phosphate phosphatase level in transgenic tobacco inhibits photosynthesis, alters carbohydrate partitioning, and reduces growth.Planta, 2005,221(4):479-492.
[20]RUAN Y L, LLEWELLYN D J, FURBANK R T. Suppression of sucrose synthase gene expression represses cotton fiber cell initiation, elongation, and seed development.ThePlantCell, 2003,15(4):952-964.
[21]孟衡玲,段承俐,萧凤回,等.铁皮石斛蔗糖合成酶基因的克隆及表达分析.中国中药杂志,2011,36(7):833-837.
MENG H L, DUAN C L, XIAO F H,etal. Molecular cloning and expression analysis of sucrose synthase gene fromDendrobiumofficinale.ChinaJournalofChineseMateriaMedica, 2011,36(7):833-837. (in Chinese with English abstract)
[22]KOCH K E. Carbohydrate-modulated gene expression in plants.AnnualReviewofPlantPhysiologyandPlantMolecularBiology, 1996,47(1):509-540.
[23]CHIOU T J, BUSH D R. Sucrose is a signal molecule in assimilate partitioning.ProceedingsoftheNationalAcademyofScienceoftheUSA, 1998,95(8):4784-4788.
[24]WIND J, SMEEKENS S, HANSON J. Sucrose: Metabolite and signaling molecule.Phytochemistry, 2010,71(14):1610-1614.
Action mechanism of sucrose and photoperiod on bulblet enlargement of Lilium sargentiae.JournalofZhejiangUniversity(Agric. &LifeSci.), 2016,42(4):435-441
ZHOU Lingyun, GAO Suping*, CHEN Feng
(SchoolofLandscapeArchitecture,SichuanAgriculturalUniversity,Chengdu611130,China)
Liliumsargentiae; sucrose; photoperiod; bulblet; enlargement; action mechanism
四川省科技计划(2012FZ0083).
Corresponding author):高素萍(http://orcid.org/0000-0002-4609-9988),E-mail:gsp65@163.com
联系方式:周玲云(http://orcid.org/0000-0002-1460-4943),E-mail:zly1065281208@126.com
2015-09-15;接受日期(Accepted):2016-01-13;网络出版日期(Published online):2016-07-18
S 682.2; Q 813.1
A
URL:http://www.cnki.net/kcms/detail/33.1247.S.20160718.2028.010.html
猜你喜欢
新农村(浙江)(2022年8期)2022-08-01
老年博览·上半月(2021年12期)2021-12-21
花卉(2021年15期)2021-08-17
昆明医科大学学报(2021年8期)2021-08-13
科学(2020年3期)2020-11-26
心理学报(2020年7期)2020-07-13
浙江农业学报(2019年7期)2019-08-14
浙江农业学报(2018年9期)2018-10-11
安徽农业科学(2018年15期)2018-05-14
少儿科学周刊·少年版(2017年7期)2017-09-29
