
浙江沿海地区舟山新木姜子群落及种群结构特征分析
2016-08-12 01:19高浩杰王国明高平仕
植物资源与环境学报 2016年1期
高浩杰 , 王国明, 高平仕
(1. 浙江省舟山市农林科学研究院, 浙江 定海 316000; 2. 浙江省岱山县农林水利围垦局, 浙江 岱山 316200)
浙江沿海地区舟山新木姜子群落及种群结构特征分析
高浩杰1,①, 王国明1, 高平仕2
(1. 浙江省舟山市农林科学研究院, 浙江 定海 316000; 2. 浙江省岱山县农林水利围垦局, 浙江 岱山 316200)
采用立木级结构代替年龄结构的方法,对浙江沿海地区(包括嵊泗县大戢山岛、定海区大猫岛、普陀区朱家尖岛、鄞州区瞻岐镇、北仑区春晓镇和宁海市力洋镇)的舟山新木姜子〔Neolitseasericea(Bl.) Koidz.〕群落的物种组成和垂直结构及其种群的年龄结构、静态生命表和分布格局进行了分析。结果表明:这6个舟山新木姜子天然群落共有维管束植物52科76属92种(含6变种),以木本植物为主,其中常绿木本植物34种,落叶木本植物31种。各群落的物种多样性指数差异较大,其中位于普陀区朱家尖岛的群落乔木层Shannon-Wiener指数(H′)最高,位于定海区大猫岛的群落灌木层和草本层的H′值均最高;且存在灌木层的H′值大于乔木层和草本层的规律。种群个体年龄结构存在差异,但6个种群均为增长型种群,幼苗储备丰富,其他龄级的个体数量总体上随径级增大而减少。6个种群的分布格局均为显著的聚集分布,其中位于北仑区春晓镇的种群聚集强度最高,位于定海区大猫岛的种群聚集强度最低。从静态生命表看,舟山新木姜子岛屿类型种群的个体死亡率在径级Ⅰ(DBH<5 cm)和径级Ⅳ(15 cm≤DBH<20 cm)较高,大陆类型种群的个体死亡率在径级Ⅰ较高。研究结果显示:浙江沿海地区舟山新木姜子群落主要为常绿阔叶林,群落间的物种多样性差异明显源于人类活动的干扰;总体上看舟山新木姜子种群具有很强的更新能力。根据研究结果,对浙江沿海地区舟山新木姜子种群的保护提出了一些建议。
舟山新木姜子; 种群结构; 物种多样性; 静态生命表; 分布格局
舟山新木姜子〔Neolitseasericea(Bl.) Koidz.〕为樟科(Lauraceae)新木姜子属〔Neolitsea(Benth. et J. D. Hook.) Merr.〕常绿乔木,主要分布于中国浙江、上海和台湾等沿海地区,日本和韩国也有分布[1-4]。舟山新木姜子多生于山坡阔叶林中或林缘,为滨海特有植物;由于其种群分布和数量稀少,已被列为稀有濒危植物[5-6],且在《国家重点保护野生植物名录(第一批)》中被列为国家Ⅱ级重点保护野生植物。舟山新木姜子木材有香气、纹理美观、春梢嫩叶金黄色、严冬绿叶丛中红果累累鲜艳夺目,为优良的观叶、观果和珍贵用材树种。目前,有关舟山新木姜子的研究主要涉及繁育技术、资源调查、抗逆性、遗传多样性和群落结构等方面[7-17]。
杨同辉等[17]的研究结果表明,分布于浙江省宁波市天童林场的舟山新木姜子群落起源于舟山市普陀种源,随种子漂流,从宁波市北仑区至奉化市、宁海县、象山县等地区的海岸线低山丘陵区可能还有舟山新木姜子的天然分布。笔者在2013年和2014年对舟山新木姜子资源的调查过程中,已经确认其在北仑区梅山岛和春晓镇、鄞州区瞻岐镇、宁海市力洋镇和象山县檀头山岛有天然分布,且在之前的研究中均未涉及。目前,关于浙江沿海地区舟山新木姜子种群年龄结构、种群分布格局和群落物种多样性方面的研究甚少,而种群与群落生态学研究是拯救与保护该物种的基础。为此,作者拟对舟山新木姜子开展群落和种群结构特征的研究,以期了解这一珍稀濒危物种的生长状况、发展趋势及濒危机制,为舟山新木姜子天然林的有效保护和种质资源保存评价提供理论依据。
1 研究区概况和研究方法
1.1研究区概况
本研究涉及的舟山新木姜子6个样地分别位于浙江省嵊泗县大戢山岛、定海区大猫岛、普陀区朱家尖岛、北仑区春晓镇、鄞州区瞻岐镇和宁海市力洋镇,基本上涵盖了舟山新木姜子在浙江分布的北界、东界、南界、西界及天然分布中心。各样地基本情况见表1。
表1浙江沿海地区舟山新木姜子天然林样地的基本情况
Table 1Basic status of natural forest plots ofNeolitseasericea(Bl.) Koidz. in coastal areas of Zhejiang Province

样地1)Plot1)纬度 Latitude 经度 Longitude 海拔/mAltitude年均温/℃Annualmeantemperature年降水量/mmAnnualprecipitation坡向Aspect坡度/(°)Slope 坡位 Slope position郁闭度/%Canopydensity生境2)Habitat2)N3)Q1N30°48'34″E122°10'20″3316.31152西West32 中Mid-dle75C128Q2N29°57'08″E122°02'41″18016.11274东East28 上Upper80C163Q3N29°54'27″E122°25'14″20616.11200北North31 上Upper95C80Q4N29°46'50″E121°50'15″12716.51317东北North-east26 中Mid-dle95H53Q5N29°45'33″E121°49'55″25316.21539东北North-east23 上Upper90H56Q6N29°23'18″E121°39'02″41216.21300北North30 中Mid-dle85V62
1)Q1: 嵊泗县大戢山岛Dajishan Island of Shengsi County; Q2: 定海区大猫岛Damao Island of Dinghai District; Q3: 普陀区朱家尖岛Zhujiajian Island of Putuo District; Q4: 北仑区春晓镇Chunxiao Town of Beilun District; Q5: 鄞州区瞻岐镇Zhanqi Town of Yinzhou District; Q6: 宁海县力洋镇Liyang Town of Ninghai County.
2)C: 海边山坡Coastal hillside; H: 山坡Hillside; V: 沟谷Valley.
3)N: 舟山新木姜子个体数量 Individual number ofNeolitseasericea.
1.2研究方法
1.2.1样地设置及调查于2014年5月至8月在上述样地,选择保存较完整、集中连片分布的舟山新木姜子天然林划分6个面积30 m×30 m的样地,用DTM-352C全站仪(日本尼康株式会社)将整个样地划分为36个5 m×5 m的样方,对样方内所有舟山新木姜子个体进行定位调查,包括胸径(DBH)、树高、坐标和生长状况等。记录各样方内木本植物(DBH≥5 cm)的种名、树高、DBH、冠幅和坐标等。在每个样地内随机选择5~7个5 m × 5 m的样方和10~14个1 m×1 m的样方,分别进行灌木和草本调查,记录灌木层和草本层中所有物种的多度、高度和盖度[18-19]。
1.2.2种群年龄结构分析采用空间代替时间的方法估算种群年龄结构,用立木级结构代替年龄结构。根据舟山新木姜子习性,以DBH 5 cm为径阶距将个体划分为8个径级:Ⅰ级(DBH<5 cm)、Ⅱ级(5 cm≤ DBH <10 cm)、 Ⅲ级(10 cm≤ DBH <15 cm)、 Ⅳ级(15 cm≤DBH<20 cm)、Ⅴ级(20 cm≤DBH<25 cm)、 Ⅵ 级(25 cm≤DBH<30 cm)、 Ⅶ 级(30 cm≤DBH<35 cm)和Ⅷ级(DBH≥35 cm)。
1.2.3种群静态生命表编制统计各龄级舟山新木姜子的个体数,按照种群静态生命表的编制方法[20-24]计算各参数,包括径级(x)、x径级开始时的存活数(nx)、x径级开始时的标准存活数(lx)、从x径级到x+1径级的死亡数(dx)、x径级的死亡率(qx)、从x径级到x+1径级的平均存活数(Lx)、x径级及以上各径级的存活个体总数(Tx)和进入x径级个体的生命期望(ex)。为了避免生命表中出现死亡率为负值的情况,采用均滑技术对nx进行处理[25]。
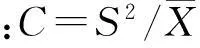
物种多样性测度:乔木植物重要值IVtr=(相对密度+相对频度+相对胸高断面积)/3;灌木和草本植物重要值IVsh=(相对密度+相对盖度+相对频度)/3;Shannon-Wiener指数H′=-∑PilnPi, 式中,Pi为物 种i的重要值[27]。
1.3数据处理和分析
用EXECL 2003软件记录数据,并采用R-3.0.1软件(http:∥www.r-project.org/,主要为Vegan包)对上述各项指标进行分析。
2 结果和分析
2.1群落物种组成和垂直结构分析
调查及统计结果显示:被调查的浙江沿海地区6个舟山新木姜子天然林样地共有维管植物52科76属92种(含6变种),包括蕨类植物4科7属8种(含种下等级,下同)、裸子植物2科2属2种、被子植物46科67属82种,其中,双子叶植物41科60属75种、单子叶植物5科7属7种。各样地均以木本植物为主,共65种,占总数的70.7%;草本植物较少,共27种,占总数的29.3%。在木本植物中,常绿木本植物34种、落叶木本植物31种,分别占木本植物总数的52.3%和47.7%。草本植物则以多年生草本为主,共22种,占草本植物总数的81.5%;一年生和二年生草本仅5种,占草本植物的18.5%。
Q1群落:位于嵊泗县大戢山岛。该样地的乔木层高8~13 m,以普陀樟〔Cinnamomumjaponicumvar.chenii(Nakai) G. F. Tao〕和舟山新木姜子占优势,常见种类有鸡桑(MorusaustralisPoir.)、红楠(MachilusthunbergiiSieb. et Zucc.)和朴树(CeltissinensisPers.)等。灌木层高1.3~3.2 m,以构树〔Broussonetiapapyifera(Linn.) L’Hér. ex Vent.〕占优势,常见种类有天仙果(FicuserectaThunb.)、大叶胡颓子(ElaeagnusmacrophyllaThunb.)和山合欢〔Albiziakalkora(Roxb.) Prain〕等。草本层主要有褐果薹草(CarexbrunneaThunb.)、渐尖毛蕨〔Cyclosorusacuminatus(Houtt.) Nakai〕和黑足鳞毛蕨(DryopterisfuscipesC. Chr.)等种类。该群落乔木层、灌木层和草本层的Shannon-Wiener指数分别为1.198、1.240和1.110。
Q2群落:位于定海区大猫岛。该样地的乔木层高7~12 m,以黄连木(Pistaciachinensis Bunge)占优势,常见种类有舟山新木姜子、黄檀(DalbergiahupeanaHance)和朴树等。灌木层高0.4~2.5 m,以华蔓茶藨子(Ribesfasciculatumvar.chinenseMaxim.)占优势,常见种类有紫金牛〔Ardisiajaponica(Thunb.) Bl.〕、野梧桐〔Mallotusjaponicus(Linn. f.) Müll. Arg.〕、山胡椒〔Linderaglauca(Sieb. et Zucc.) Bl.〕和柞木〔Xylosmaracemosum(Sieb. et Zucc.) Miq.〕等。草本层主要有酸模(RumexacetosaLinn.)、丝穗金粟兰〔Chloranthusfortunei(A. Gray) Solms-Laub.〕和蓟(CirsiumjaponicumFisch. ex DC.)等种类。该群落乔木层、灌木层和草本层的Shannon-Wiener指数分别为0.925、2.920和2.275。
Q3群落:位于普陀区朱家尖岛。该样地的乔木层高6~10 m,以舟山新木姜子占优势,常见种类有普陀樟、紫楠〔Phoebesheareri(Hemsl.) Gamble〕、青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕、日本珊瑚树〔Viburnumodoratissimumvar.awabuki(K. Koch) Zabel ex Rümpl.〕等。灌木层高0.6~2.0 m,以山茶(CamelliajaponicaLinn.)占优势,常见种类有朱砂根(ArdisiacrenataSims)、清风藤(SabiajaponicaMaxim.)和南五味子(KadsuralongipedunculataFinet et Gagnep.)等。草本层主要有阔鳞鳞毛蕨〔Dryopterischampionii(Benth.) C. Chr.〕、褐果薹草和显子草(PhaenospermaglobosaMunro ex Benth.)等种类。该群落乔木层、灌木层和草本层的Shannon-Wiener指数分别为2.048、2.503和1.334。
Q4群落:位于北仑区春晓镇。该样地的乔木层高8~13 m,以舟山新木姜子和紫楠占优势,常见种类有红楠、石栎〔Lithocarpusglaber(Thunb.) Nakai〕和青冈等。灌木层高0.5~3.0 m,以天仙果占优势,常见种类有山鸡椒〔Litseacubeba(Lour.) Pers.〕、茶树〔Camelliasinensis(Linn.) Kuntze〕和紫麻〔Oreocnidefrutescens(Thunb.) Miq.〕等。草本层主要有狗脊〔Woodwardiajaponica(Linn. f.) Sm.〕、蕨〔Pteridiumaquilinumvar.latiusculum(Desv.) Underw. ex Heller〕和玄参(ScrophularianingpoensisHemsl.)等种类。该群落乔木层、灌木层和草本层的Shannon-Wiener指数分别为1.454、1.955和1.010。
Q5群落:位于鄞州区瞻岐镇。该样地的乔木层高6~11 m,以青冈占优势,常见种类有舟山新木姜子、椿叶花椒(ZanthoxylumailanthoidesSieb. et Zucc.)和红淡比(CleyerajaponicaThunb.)等。灌木层高1.2~2.5 m,以毛花连蕊茶(CamelliafraternaHance)占优势,常见种类有矩叶卫矛(EuonymusoblongifoliusLoes. et Rehd.)、胡颓子(ElaeagnuspungensThunb.)和三尖杉(CephalotaxusfortuneiHook.)等。草本层主要有蘘荷〔Zingibermioga(Thunb.) Rosc.〕、盾蕨〔Neolepisorusovatus(Bedd.) Ching〕和蔓赤车(PellioniascabraBenth.)等。该群落乔木层、灌木层和草本层的Shannon-Wiener指数分别为1.175、1.652和1.265。
Q6群落:位于宁海县力洋镇。该样地的乔木层高10~14 m,以红楠、舟山新木姜子占优势,常见种类有青冈和木荷(SchimasuperbaGardn. et Champ.)等。灌木层高1.4~2.8 m,以毛花连蕊茶占优势,常见种类有茶、朱砂根和楤木(AraliachinensisLinn.)等。草本层主要有刺头复叶耳蕨〔Arachniodesexilis(Hance) Ching〕、野青茅〔Deyeuxiapyramidalis(Host) Veldkamp〕和小野芝麻〔Galeobdolonchinense(Benth.) C. Y. Wu〕等。该群落乔木层、灌木层和草本层的Shannon-Wiener指数分别为1.239、1.547和1.199。
2.2种群结构和分布格局分析
2.2.1种群年龄结构分析浙江沿海地区6个舟山新木姜子种群的径级结构见表2。由表2可见,位于嵊泗县大戢山岛Q1样地和定海区大猫岛Q2样地的舟山新木姜子种群中均缺乏Ⅶ级(30 cm≤DBH<35 cm)和Ⅷ级(DBH≥35 cm)成龄树,最大径级只达到Ⅵ级(25 cm≤DBH<30 cm);这2个种群中Ⅰ级(DBH<5 cm)幼龄树所占比例均较高,分别为86.9%和86.1%,且高于其他4个种群,说明这2个种群总体更新状况最好,种群结构呈明显的增长型。
Q3、Q4、Q5和Q6样地的舟山新木姜子种群中均存在植株高大且Ⅵ级以上成龄树占一定比例的现象。其中,位于普陀区朱家尖岛Q3样地的种群以Ⅰ级个体为主,Ⅵ级(25 cm≤DBH<30 cm)个体也占一定比例,除Ⅴ级(20 cm≤DBH<25 cm)个体外,该种群在各径级均有个体分布,种群径级相对完整;位于鄞州区瞻岐镇Q4样地的种群以Ⅰ级个体为主,Ⅱ级(5 cm≤DBH<10 cm)个体也占一定比例,Ⅶ级和Ⅷ级成龄树所占比例达11.9%,在6个种群中最高;位于北仑区春晓镇Q5样地的种群中Ⅰ级幼龄树所占比例较小,为60.5%,低于其他5个种群,但Ⅱ级至Ⅵ级中龄树所占比例较高,为33.8%,高于其他5个种群;位于宁海市力洋镇Q6样地的种群则以Ⅰ级和Ⅱ级个体为主,但Ⅲ级(10 cm≤DBH<15 cm)和Ⅵ级个体缺失。
舟山新木姜子种群各龄级的个体数量总体上随径级增大而减少,符合发育良好的种群径级结构的普遍规律。6个种群中Ⅰ级幼龄树所占比例均超过60%,表明舟山新木姜子小径级植株储备充足,利于其种群的天然更新。北仑、鄞州和宁海3个样地的舟山新木姜子种群中Ⅶ级和Ⅷ级成龄树所占比例均高于嵊泗、定海和普陀样地的舟山新木姜子种群,主要是因为岛屿类型种群频繁受到台风和海雾的袭击,在大风和盐雾胁迫下舟山新木姜子的胸径和树高生长均受到抑制,导致大径级植株数量相对少于大陆类型种群。
表2浙江沿海地区6个舟山新木姜子种群的径级结构
Table 2Diameter class structure of six populations ofNeolitseasericea(Bl.) Koidz. in coastal areas of Zhejiang Province

样地1)Plot1)不同径级个体数所占比例/%2) Percentageofindividualnumberindifferentdiameterclasses2)ⅠⅡⅢⅣⅤⅥⅦⅧQ186.91.44.84.81.40.70.00.0Q286.15.22.63.12.11.00.00.0Q384.31.41.41.40.06.33.31.9Q473.89.52.40.02.40.04.87.1Q560.516.311.64.61.30.00.05.6Q670.814.60.02.12.10.04.26.3
1)Q1: 嵊泗县大戢山岛Dajishan Island of Shengsi County; Q2: 定海区大猫岛Damao Island of Dinghai District; Q3: 普陀区朱家尖岛Zhujiajian Island of Putuo District; Q4: 北仑区春晓镇Chunxiao Town of Beilun District; Q5: 鄞州区瞻岐镇Zhanqi Town of Yinzhou District; Q6: 宁海县力洋镇Liyang Town of Ninghai County.
2)Ⅰ: DBH<5 cm; Ⅱ: 5 cm≤DBH<10 cm; Ⅲ: 10 cm≤DBH<15 cm; Ⅳ: 15 cm≤DBH<20 cm; Ⅴ: 20 cm≤DBH<25 cm; Ⅵ: 25 cm≤DBH<30 cm; Ⅶ: 30 cm≤DBH<35 cm; Ⅷ: DBH≥35 cm.
2.2.2种群静态生命表分析由于舟山新木姜子岛屿类型种群和大陆类型种群地理隔离较远,生境差异较大,位于6个样地的种群年龄结构存在一定差异,且均存在个别径级的缺失,故以岛屿类型种群(Q1、Q2、Q3)和大陆类型种群(Q4、Q5、Q6)为对象分别编制种群静态生命表,结果见表3。
表3浙江沿海地区舟山新木姜子岛屿类型和大陆类型种群的静态生命表1)
Table 3Static life table of island type and mainland type populations ofNeolitseasericea(Bl.) Koidz. in coastal areas of Zhejiang Province1)
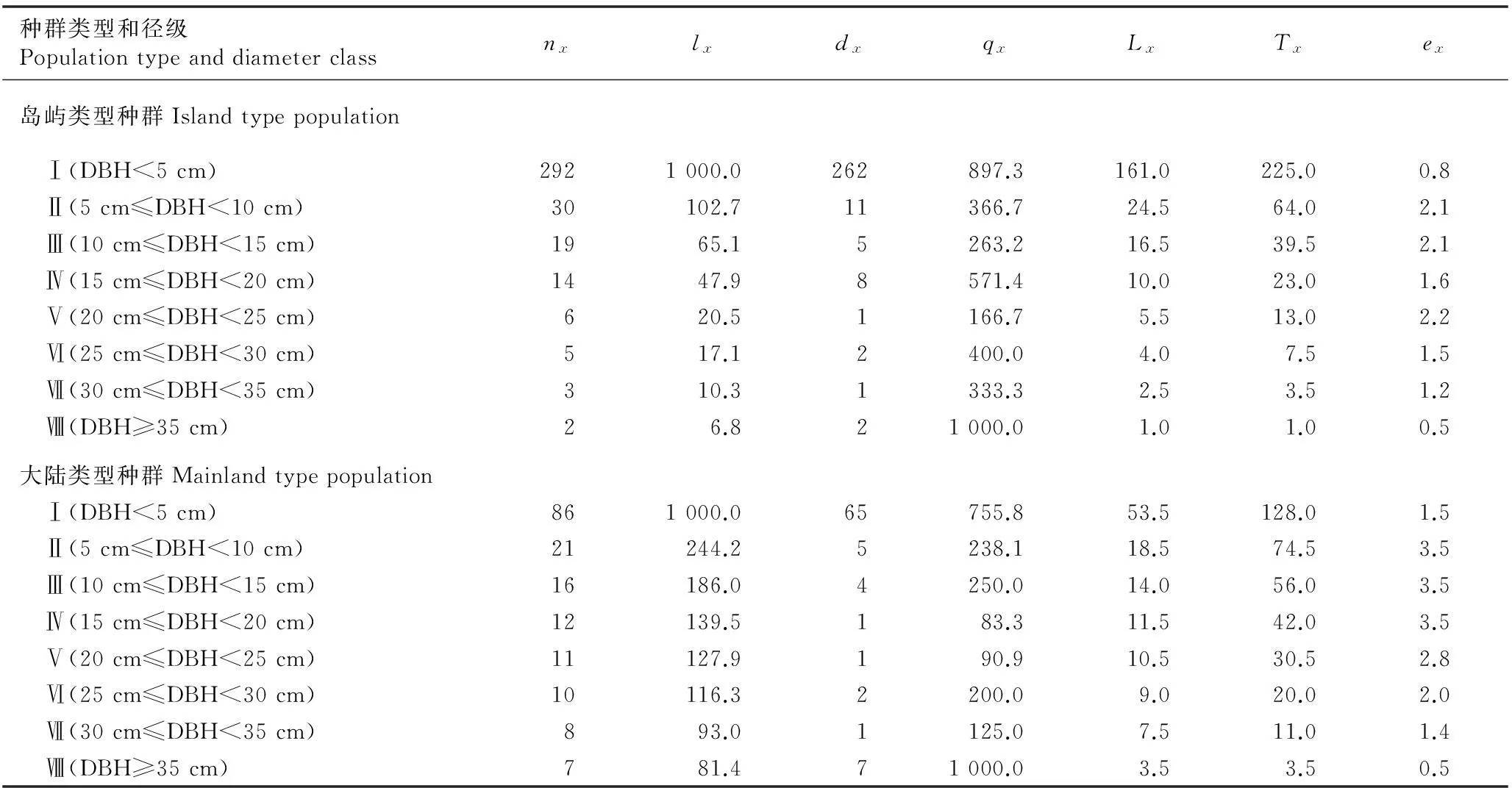
种群类型和径级PopulationtypeanddiameterclassnxlxdxqxLxTxex岛屿类型种群Islandtypepopulation Ⅰ(DBH<5cm)2921000.0262897.3161.0225.00.8 Ⅱ(5cm≤DBH<10cm)30102.711366.724.564.02.1 Ⅲ(10cm≤DBH<15cm)1965.15263.216.539.52.1 Ⅳ(15cm≤DBH<20cm)1447.98571.410.023.01.6 Ⅴ(20cm≤DBH<25cm)620.51166.75.513.02.2 Ⅵ(25cm≤DBH<30cm)517.12400.04.07.51.5 Ⅶ(30cm≤DBH<35cm)310.31333.32.53.51.2 Ⅷ(DBH≥35cm)26.821000.01.01.00.5大陆类型种群Mainlandtypepopulation Ⅰ(DBH<5cm)861000.065755.853.5128.01.5 Ⅱ(5cm≤DBH<10cm)21244.25238.118.574.53.5 Ⅲ(10cm≤DBH<15cm)16186.04250.014.056.03.5 Ⅳ(15cm≤DBH<20cm)12139.5183.311.542.03.5 Ⅴ(20cm≤DBH<25cm)11127.9190.910.530.52.8 Ⅵ(25cm≤DBH<30cm)10116.32200.09.020.02.0 Ⅶ(30cm≤DBH<35cm)893.01125.07.511.01.4 Ⅷ(DBH≥35cm)781.471000.03.53.50.5
1)nx:x径级开始时的存活数Survival number at the beginning ofxdiameter class;lx:x径级开始时的标准存活数Standard survival number at the beginning ofxdiameter class;dx: 从x径级到x+1径级的死亡数Death number fromxdiameter class tox+1 diameter class;qx:x径级的死亡率Mortality rate atxdiameter class;Lx: 从x径级到x+1径级的平均存活数Average survival number fromxdiameter class tox+1 diameter class;Tx:x径级及以上各径级的存活个体总数Total survival individual number fromxdiameter class to higher diameter class;ex: 进入x径级个体的生命期望Life expectancy of individual number intoxdiameter class.
由表3可知:舟山新木姜子岛屿类型种群生长过程中出现2次死亡高峰。第1次死亡高峰出现在径级Ⅰ到径级Ⅱ,死亡率为89.7%,生命期望(ex)也相对较低,为0.8;第2次死亡高峰出现在径级Ⅳ到径级Ⅴ,死亡率为57.1%。舟山新木姜子大陆类型种群生长过程中仅在径级Ⅰ至径级Ⅱ时出现明显的死亡高峰,死亡率为75.6%,其他径级的死亡率均不高。2种类型种群在幼树期间均有50%以上的个体死亡,这一现象与其年龄结构分布一致。主要原因是随着舟山新木姜子幼树个体的生长,其所需的环境资源也更多,争光、争水、争肥和争空间现象明显加剧,导致自疏或它疏作用强烈,大量植株被淘汰。此外,岛屿类型种群在径级Ⅳ死亡率相对较高,可能与特殊的岛屿生境(如风大、土壤瘠薄和水热条件差)有关,较差的立地条件对其生长产生制约作用。
2.2.3种群分布格局分析浙江沿海地区6个舟山新木姜子种群的空间分布格局见表4。由表4可知:舟山新木姜子6个种群的扩散系数(C)均大于1、负二项参数(k)均大于0、聚块性指数(PAI)均大于1、Morisita指数(Iδ)均大于1,表明各种群分布格局均为聚集分布。对C值的t检验和Iδ值的F检验结果则显示,供试的6个舟山新木姜子种群的分布格局均为极显著的聚集分布。
从平均拥挤度指数(m*)判断,6个舟山新木姜子种群的拥挤强度由高至低依次排序为Q3种群、Q1种群、Q2种群、Q4种群、Q5种群、Q6种群。从PAI值判断,6个舟山新木姜子种群的聚集强度由高至低依次排序为Q4种群、Q5种群、Q3种群、Q1种群、Q6种群、Q2种群。分析结果表明:岛屿类型种群的拥挤度高于大陆类型种群,这可能与岛屿类型种群中舟山新木姜子的个体数相对较多有关。Q4种群的聚集强度最高,说明位于北仑区春晓镇的这个舟山新木姜子种群的分布范围最窄。
6个舟山新木姜子种群的分布格局均为聚集分布,主要原因是舟山新木姜子为典型的雌雄异株植物,以有性繁殖为主,种子扩散以雌株为中心,但其果实大而重,传播距离较近,导致雌株周围的种子量大于其他区域,实生苗往往聚集在雌株周围生长,因此容易形成聚集分布。此外,北仑、鄞州和普陀3个样地的舟山新木姜子种群的聚集强度高于嵊泗、宁海和定海3个样地,很可能是因为前者林分郁闭度高,而舟山新木姜子种子萌发对光照有一定的依赖性,为满足光照条件幼苗大都聚集在光线较充足的林窗内或林缘生长,导致种群的聚集强度较高;而后者郁闭度较低,光照条件相对一致,种群分布的聚集强度也相应较低。
表4浙江沿海地区6个舟山新木姜子种群的空间分布格局
Table 4Spatial distribution pattern of six populations ofNeolitseasericea(Bl.) Koidz. in coastal areas of Zhejiang Province
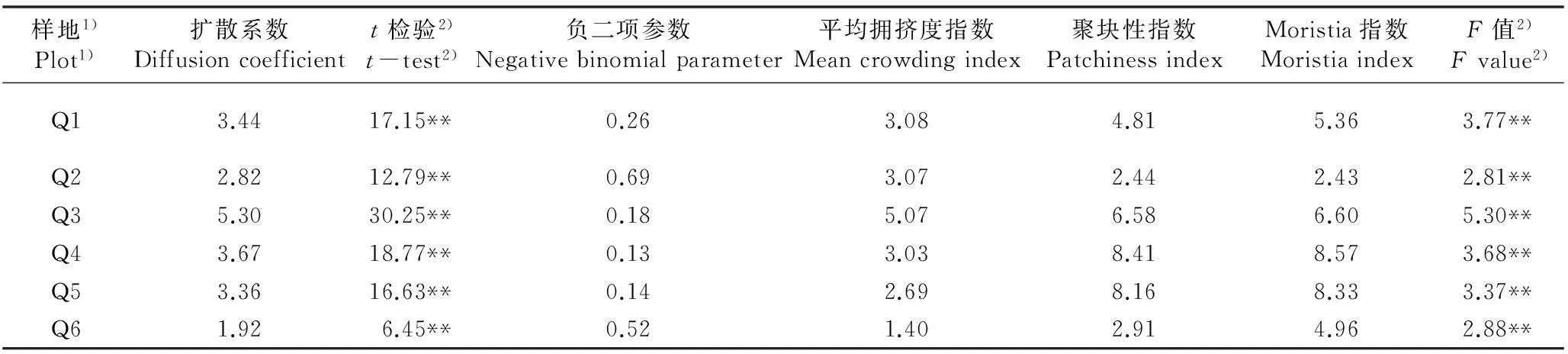
样地1)Plot1)扩散系数Diffusioncoefficientt检验2)t-test2)负二项参数Negativebinomialparameter平均拥挤度指数Meancrowdingindex聚块性指数PatchinessindexMoristia指数MoristiaindexF值2)Fvalue2)Q13.4417.15**0.263.084.815.363.77**Q22.8212.79**0.693.072.442.432.81**Q35.3030.25**0.185.076.586.605.30**Q43.6718.77**0.133.038.418.573.68**Q53.3616.63**0.142.698.168.333.37**Q61.926.45**0.521.402.914.962.88**
1)Q1: 嵊泗县大戢山岛Dajishan Island of Shengsi County; Q2: 定海区大猫岛Damao Island of Dinghai District; Q3: 普陀区朱家尖岛Zhujiajian Island of Putuo District; Q4: 北仑区春晓镇Chunxiao Town of Beilun District; Q5: 鄞州区瞻岐镇Zhanqi Town of Yinzhou District; Q6: 宁海县力洋镇Liyang Town of Ninghai County.
2)**: 差异极显著Extremely significant difference.
3 讨论和结论
调查结果表明:浙江沿海舟山新木姜子群落共有维管束植物52科76属92种,常绿木本、落叶木本、多年生草本以及一年生和二年生草本分别占37.0%、33.7%、23.9%和5.4%。王凤英等[28]认为,舟山新木姜子群落具有亚热带常绿阔叶林的特点;本研究中,位于定海区大猫岛的舟山新木姜子群落是以落叶树种占优势的常绿落叶阔叶混交林,这与位于浙江天童的舟山新木姜子群落特征[17]相似,其他5个群落均属于常绿阔叶林。从物种多样性水平来看,6个舟山新木姜子群落中乔木层Shannon-Wiener指数最高的是位于普陀区朱家尖岛的群落,最低的是位于定海区大猫岛的群落;灌木层Shannon-Wiener指数最高的定海区大猫岛群落,最低的是嵊泗县大戢山岛群落;草本层Shannon-Wiener指数最高的也是定海区大猫岛群落,最低的是北仑区春晓镇群落。Shannon-Wiener指数在各群落中均表现出灌木层大于乔木层和草本层的规律,与典型常绿阔叶林通常具有的“灌木层多样性指数大于乔木层,乔木层多样性指数又大于草本层”的格局基本一致[29]。
舟山新木姜子各群落的物种多样性差异明显主要源于人类活动的干扰。如位于普陀区朱家尖岛的群落面海孤山且交通闭塞,干扰程度较低;而位于定海区大猫岛的群落则处于村庄附近,周围是反复开垦的旱地,干扰程度较高,因而,前者的物种多样性较高(乔木层Shannon-Wiener指数2.048),而后者的物种多样性则较低(乔木层Shannon-Wiener指数0.925)。这也从侧面反映了舟山新木姜子对环境条件要求较高,一旦出现人为干扰,生境遭受一定程度的破坏,其群落物种多样性也随之降低。因此,应针对不同的群落现状,采取适当的技术措施,拯救和保护现有的舟山新木姜子天然林资源。例如,对位于普陀区朱家尖岛的舟山新木姜子群落可以通过封山育林维持天然林规模稳定,而对定海区大猫岛的群落可以通过定株抚育促进林分质量提高。
舟山新木姜子各种群的年龄结构存在差异,但总体上均表现为增长型种群,表明舟山新木姜子具有很强的更新能力。随着群落演替的推进,在没有人为干扰和特殊自然灾害的情况下,舟山新木姜子将维持其在群落中优势种和主要组成树种的地位,这一结果与“浙江天童舟山新木姜子胸径级频率分布呈逆J字型”的研究结论一致[17]。从本研究结果还可以看出,舟山新木姜子种群虽有丰富的幼苗储备,但在成长过程中幼苗大量死亡,中龄树的存活率很低。因此在生产实践中,针对这种情况应对幼树进行适当的人工移植,既有助于舟山新木姜子天然种群的更新,又可为其回归自然提供种苗。整体上看,舟山新木姜子岛屿类型种群中小径级个体的数量相对较多,但较差的立地条件制约了个体的生长,导致大径级个体数量较少;而大陆类型种群中虽然小径级个体数量相对较少,但大径级个体数量却较多。6个舟山新木姜子种群均呈极显著的聚集分布,这与其种子散布的规律和生物学特性有关。舟山新木姜子为典型的雌雄异株植物,因重力作用造成其种子多散布在雌株周围,进而表现出由种子萌发产生的幼树聚集分布。
舟山新木姜子6个种群的年龄结构和生长繁殖特性均存在差异,针对这一现象,建议开展舟山新木姜子不同地理种群生态适应性研究,并从中选育出适宜浙江沿海推广的优良品种。舟山新木姜子6个种群地理跨度较远且部分种群处于岛屿隔离状态,由于舟山新木姜子具有中等偏低水平的遗传多样性[9],因此建议将大陆类型种群和岛屿类型种群的个体进行混合繁殖,不仅能够促进种群间基因交流,还有利于维持舟山新木姜子的遗传多样性。
[1]中国科学院中国植物志编辑委员会. 中国植物志: 第三十一卷[M]. 北京: 科学出版社, 1982: 348-349.
[2]WU Z Y, RAVEN P H, HONG D Y. Flora of China: Vol. 7[M]. Beijing: Science Press, 2008: 109.
[3]MAKINOT.NewIllustratedFloraofJapan[M].Tokyo: Hokuryukan, 1979: 194.
[4]林弥栄. 日本の樹木[M]. 増補改訂新版. 東京: 山と溪谷社, 2011: 215.
[5]傅立国. 中国植物红皮书——稀有濒危植物: 第一册[M]. 北京: 科学出版社, 1991: 356.
[6]国家林业局野生动植物保护和自然保护区管理司, 中国科学院植物研究所. 中国珍稀濒危植物图鉴[M]. 北京: 中国林业出版社, 2013: 199.
[7]孙敏琴. 舟山新木姜子扦插繁殖试验初报[J]. 浙江林业科技, 1999, 19(4): 62-63, 71.
[8]丁方明, 张成标, 卢小根. 舟山新木姜子资源调查报告[J]. 浙江林业科技, 2001, 21(4): 52-54.
[9]王中生, 安树青, 冷欣, 等. 岛屿植物舟山新木姜子居群遗传多样性的RAPD分析[J]. 生态学报, 2004, 24(3): 414-422.
[10]王慰, 黄胜利, 丁国剑, 等. 盐胁迫下舟山新木姜子1年生苗形态变化及生理反应[J]. 浙江林学院学报, 2007, 24(2): 168-172.
[11]俞慈英, 陈叶平, 袁燕飞, 等. 舟山海岛普陀樟等3种特有树种种质资源清查[J]. 浙江林学院学报, 2007, 24(4): 413-418.
[12]徐斌芬, 王国明, 陈斌, 等. 舟山新木姜子和普陀樟容器育苗试验[J]. 林业科技开发, 2007, 21(5): 71-73.
[13]李影丽, 汪奎宏, 许利群, 等. 舟山新木姜子盐胁迫下生长变化及生理反应[J]. 浙江林业科技, 2008, 28(2): 48-51.
[14]李修鹏, 赵慈良, 俞慈英, 等. 舟山新木姜子保存技术研究[J]. 浙江海洋学院学报: 自然科学版, 2009, 28(1): 81-85.
[15]应松康, 赵颖, 陈斌, 等. 舟山新木姜子苗木分级及不同密度试验[J]. 浙江林业科技, 2009, 29(3): 80-83.
[16]杜佩剑, 徐迎春, 李永荣. 舟山新木姜子容器育苗基质配方研究[J]. 江苏林业科技, 2010, 37(1): 1-4, 8.
[17]杨同辉, 王良衍. 浙江天童舟山新木姜子群落结构及其更新类型[J]. 浙江林学院学报, 2010, 27(6): 877-883.
[18]胡尔查, 王晓江, 刘永宏, 等. 乌拉山自然保护区油松种群结构与分布格局研究[J]. 林业科学研究, 2011, 24(3): 363-369.
[19]宋萍, 洪伟, 吴承祯, 等. 珍稀濒危植物桫椤种群结构与动态研究[J]. 应用生态学报, 2005, 16(3): 413-418.
[20]康华靖, 陈子林, 刘鹏, 等. 大盘山自然保护区香果树种群结构与分布格局[J]. 生态学报, 2007, 27(1): 389-396.
[21]何东, 魏新增, 李连发, 等. 神农架山地河岸带连香树的种群结构与动态[J]. 植物生态学报, 2009, 33(3): 469-481.
[22]KOSTRAKIEWICZ K. Population structure of a clonal endangered plant speciesIrissibiricaL. in different habitat conditions[J]. Polish Journal of Ecology, 2008, 56: 581-592.
[23]WU J X, ZHANG X M, DENG C Z, et al. Structure and dynamic ofPopuluseuphraticapopulation along Tarim River[J]. Pakistan Journal of Botany, 2012, 44: 1651-1656.
[24]LI W, ZHANG G F. Population structure and spatial pattern of the endemic and endangered subtropical treeParrotiasubaequalis(Hamamelidaceae)[J]. Flora, 2015, 212: 10-18.
[25]何东进, 肖石红, 李晓景, 等. 中国红树林北缘区秋茄树种群空间分布格局和结构特征分析[J]. 植物资源与环境学报, 2014, 23(2): 41-47.
[26]李吉玫, 徐海量, 张青青, 等. 塔里木河下游荒漠河岸林不同退化区胡杨种群结构和空间分布格局研究[J]. 中国沙漠, 2009, 29(5): 897-904.
[27]王德国, 邢韶华, 崔国发, 等. 甘肃连城自然保护区8种主要森林类型的植物物种多样性研究[J]. 西部林业科学, 2008, 37(3): 51-55.
[28]王凤英, 田旗, 彭红玲, 等. 舟山群岛2种濒危植物生境特征与迁地保护研究[J]. 浙江农林大学学报, 2014, 31(3): 417-423.
[29]贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J]. 植物生态学报, 1998, 22(4): 303-311.
(责任编辑: 张明霞)
Characteristic analysis on community and population structure ofNeolitseasericeain coastal areas of Zhejiang
GAO Haojie1,①, WANG Guoming1, GAO Pingshi2
(1. Zhoushan Academy of Agriculture and Forestry Sciences of Zhejiang, Dinghai 316000, China; 2. Daishan Agri-forest and Water Conservancy Bureau of Zhejiang, Daishan 316200, China),J.PlantResour. &Environ., 2016, 25(1): 94-101
Taking method of tree class structure replacing age structure, species composition and vertical structure of community, and age structure, static life table and distribution pattern of populations ofNeolitseasericea(Bl.) Koidz. in coastal areas of Zhejiang (including Dajishan Island of Shengsi County, Damao Island of Dinghai District, Zhujiajian Island of Putuo District, Zhanqi Town of Yinzhou District, Chunxiao Town of Beilun District and Liyang Town of Ninghai County) were analyzed. The results show that there are vascular plants of 92 species (including 6 varieties) in 76 genera belonging to 52 families in these sixN.sericeanatural communities. Woody plants are the main plants, in which there are 34 species of evergreen woody trees and 31 species of deciduous woody trees. There are great differences in species diversity indexes, in which Shannon-Wiener index (H′) of arbor layer in community located Zhujiajian Island of Putuo District is the highest,H′ values of shrub and herb layers in community located Damao Island of Dinghai District both are the highest, andH′ value of shrub layer is higher than that of arbor and herb layers. There are differences in age structure of individuals of populations, but six populations ofN.sericeaall are growing type, seedlings are abundant, and number of individuals in other age classes decreases with increasing of diameter class. Distribution patterns of six populations all are significant aggregation distribution, in which, aggregation intensity of population located Chunxiao Town of Beilun District is the highest, and that of population located Damao Island of Dinghai District is the lowest. According to static life table, individual mortality rates at Ⅰ diameter class (DBH<5 cm) and Ⅳ diameter class (15 cm≤DBH<20 cm) of island type population ofN.sericeaare high, and that at Ⅰ diameter class of mainland type population is also high. It is suggested thatN.sericeacommunity in coastal areas of Zhejiang is main evergreen broad-leaved forest, the difference in species diversity among communities is obviously due to interference of human activities; on a whole,N.sericeapopulation has a high renewal ability. Based on research result, some suggestions for protectingN.sericeapopulation in coastal areas of Zhejiang are proposed.
Neolitseasericea(Bl.) Koidz.; population structure; species diversity; static life table; distribution pattern
10.3969/j.issn.1674-7895.2016.01.12
2015-04-17
浙江省舟山市科技计划项目(2013C31074)
高浩杰(1985—),男,浙江岱山人,本科,工程师,主要从事林业科技推广方面的工作。
Q948.15; S718.54
A
1674-7895(2016)01-0094-08
猜你喜欢
广西林业科学(2022年4期)2022-11-07
贵州畜牧兽医(2022年3期)2022-06-28
林业调查规划(2022年3期)2022-06-13
现代园艺(2021年23期)2021-12-01
青年文学家(2021年16期)2021-08-05
园艺与种苗(2021年3期)2021-05-26
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
少儿美术(2020年8期)2020-11-06
中国民族医药杂志(2016年1期)2016-05-09
