
饲料中不同含量豆粕对乌苏里拟鲿生长、体成分及表观消化率的影响
2016-08-10 06:15:49王裕玉周歧存卜宪勇苏宝辉杨雨虹
水生生物学报 2016年4期
王裕玉周歧存卜宪勇苏宝辉杨雨虹
(1. 东北农业大学动物科学技术学院, 哈尔滨 150030; 2. 宁波大学海洋学院, 宁波 315211)
饲料中不同含量豆粕对乌苏里拟鲿生长、体成分及表观消化率的影响
王裕玉1周歧存2卜宪勇1苏宝辉1杨雨虹1
(1. 东北农业大学动物科学技术学院, 哈尔滨 150030; 2. 宁波大学海洋学院, 宁波 315211)
以初始体重(0.50±0.00) g的乌苏里拟鲿(Pseudobagrus ussuriensis)稚鱼为试验对象, 在室内进行为期8周的摄食生长实验, 研究饲料中豆粕替代鱼粉蛋白对乌苏里拟鲿生长、体成分及消化率的影响。以豆粕分别替代对照饲料(S0)中10%(S10)、20%(S20)、30%(S30)、40%(S40)、50%(S50)和60%(S60)的鱼粉蛋白, 另外,60%组添加0.3%蛋氨酸(SM60), 共配制8种等氮等能试验饲料。结果表明, 随着饲料中豆粕替代比例的增大,试验鱼的增重率、特定生长率和蛋白质效率均逐渐降低, 但10%—40%替代组与对照组之间差异不显著(P>0.05), S50和S60显著低于对照组(P<0.05); 与S60组相比, SM60组增重率和特定生长率有所提高(P>0.05),但显著低于0—40%替代组(P<0.05); 饲料系数的变化规律与增重率的趋势正好相反; 当饲料中豆粕替代水平由0增加到20%时, 乌苏里拟鲿的摄食率由3.22%提高到3.55%, 而后随着替代比例进一步增加, 摄食率逐渐降低, 但方差分析表明, 差异不显著。饲料中豆粕替代鱼粉对乌苏里拟鲿全鱼水分和灰分含量的影响不显著, 肌肉水分随着替代比例的升高而逐渐升高; 全鱼和肌肉粗蛋白含量随着替代比例的升高而逐渐降低, 其中,S60和SM60组显著低于其他组(P<0.05); 随着替代比例的升高, 全鱼和肌肉粗脂肪呈现升高的趋势, S60和SM60组显著高于其他替代组(P<0.05); 肥满度在各组间差异不显著(P>0.05); 当豆粕替代鱼粉蛋白比例为60%时, 肝体比、脏体比指数显著高于其他组(P<0.05), 而其他组间差异不显著(P>0.05)。50%和60%替代组的蛋白质和干物质的表观消化率显著低于0—40%替代组(P<0.05), 而0—40%替代组之间差异不显著(P>0.05);随着豆粕替代比例由0升高到60%, 磷表观消化率由36.07% 升高到66.50%。在实验条件下, 豆粕替代乌苏里拟鲿饲料中40%鱼粉蛋白, 不会影响乌苏里拟鲿的生长和饲料利用, 而高比例替代对生长和鱼的健康有负面影响。
乌苏里拟鲿; 鱼粉; 豆粕; 生长; 表观消化率
鱼粉因其具有高蛋白质、富含必需氨基酸、脂肪酸和未知生长因子、适口性好以及较好的消化率等优点, 一直以来被认为是水产饲料中优质的蛋白源[1]。然而, 近年来, 由于过度捕捞和环境污染等影响, 引起海洋渔业资源衰减, 鱼粉的总产量逐年下降, 而当今世界水产养殖业却以年均11%的速度增长[2]。New[3]预言, 到2020年全球水产饲料中对鱼粉的需求量将超过总的鱼粉供应量, 供给与需求关系的不平衡导致鱼粉价格飙升, 使饲料成本升高, 最终会导致养殖效益下降。另外, 鱼粉中的磷利用率相对较低, 未被吸收的磷会排入水环境中,导致水体富营养化。基于经济和生态效益, 人们急切寻找能够部分或完全替代鱼粉的质优廉价的蛋白源, 这既有利于保护渔业资源, 又有利于节约饲料成本, 提高养殖效益。
大豆蛋白具有氨基酸组成较好、消化吸收率高、资源丰富和价格低廉等优点, 因而常常被用作水产饲料中鱼粉的替代物。研究表明, 当饲料中适量添加豆粕时, 对鱼类的生长性能、饲料利用以及鱼体健康等方面无显著影响, 但当添加量超出鱼类的耐受极限时, 鱼类的生长降低, 死亡率上升, 尤其是作为单一蛋白源时, 会导致生长急剧下降, 且长期使用会影响鱼类的肉质[4—7], 这主要是由于添加量较高时, 饲料中的必需氨基酸不平衡, 尤其是蛋氨酸的缺乏, 会严重抑制鱼类的生长; 饲料中的抗营养因子例如大豆抗原、大豆凝集素的含量会随之升高, 饲料的适口性会降低, 并会导致肠道和肝脏组织损伤, 发生病变。
乌苏里拟鲿(Pseudobagrus ussuriensis), 地方名牛尾巴, 隶属于鲶形目、鲿科、拟鲿属鱼类, 是黑龙江流域特有的鱼类[8]。其肉质细嫩, 无肋间刺, 味道特别鲜美, 营养价值高, 是饭店的上乘佳肴。目前, 有关于乌苏里拟鲿的研究主要集中在繁殖生物学[9]、人工繁育技术[10]和生化遗传学[11]领域, 而关于其营养生理及配合饲料的研发的研究较少。为进一步推广这一养殖品种的规模化及集约化养殖,本课题组对乌苏里拟鲿蛋白、脂肪、赖氨酸、蛋氨酸的营养需求及其营养价值进行了系统的研究。本实验以(0.50±0.00) g的乌苏里拟鲿稚鱼为试验对象, 探讨豆粕替代鱼粉蛋白对乌苏里拟鲿生长性能、饲料利用、体组成和营养物质消化率等的影响, 为乌苏里拟鲿配合饲料中豆粕的应用提供技术支持。
1 材料与方法
1.1 试验饲料
以鱼粉为主要蛋白源, 鱼油为脂肪源, 面粉作为碳水化合物源, 配制基础饲料(S0)。以豆粕分别替代基础饲料中10% (S10)、20% (S20)、30% (S30)、40% (S40)、50% (S50)和60% (S60)的鱼粉蛋白, 配制7种等氮等能试验饲料。另外, 为了验证高比例豆粕替代后, 添加晶体氨基酸的作用效果,向60%组添加0.3%蛋氨酸(SM60)。饲料配方及营养成分分析见表 1。饲料氨基酸组成见表 2。
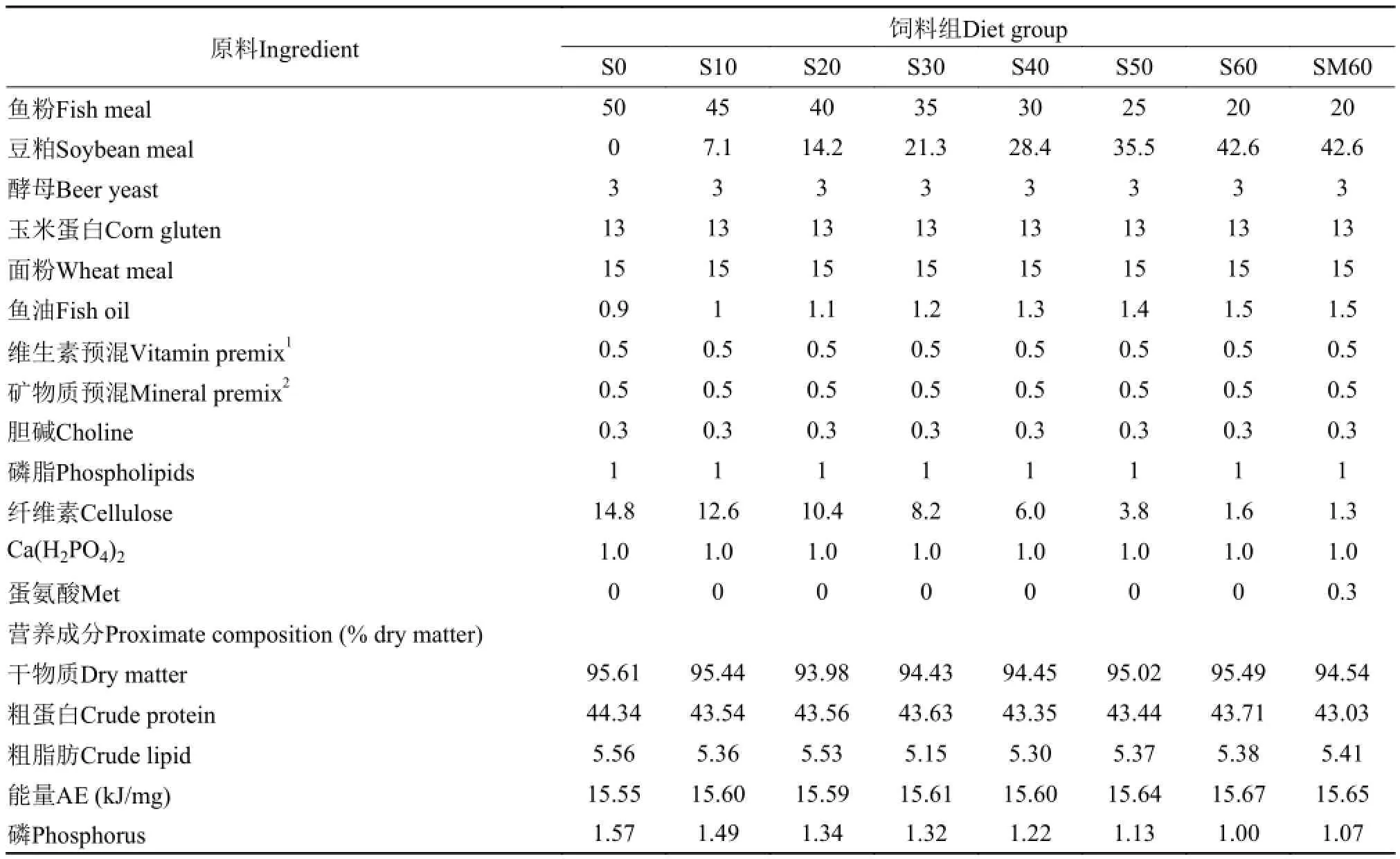
表 1 试验饲料组成及营养成分(%干物质)Tab. 1 Ingredients and nutrients of the experimental diets (% dry matter)
配制饲料前, 所有原料经过粉碎, 过60目筛。准确称量饲料原料, 先将固体原料混合均匀, 然后再加入鱼油和磷脂, 手工将油脂颗粒搓细, 混匀, 最后添加30%的水, 再次混匀, 用强力牌双螺杆制粒机(河南偃师)挤压成条状(直径约1.0 mm), 60℃烘干, 封口袋包装, -20℃保存备用。
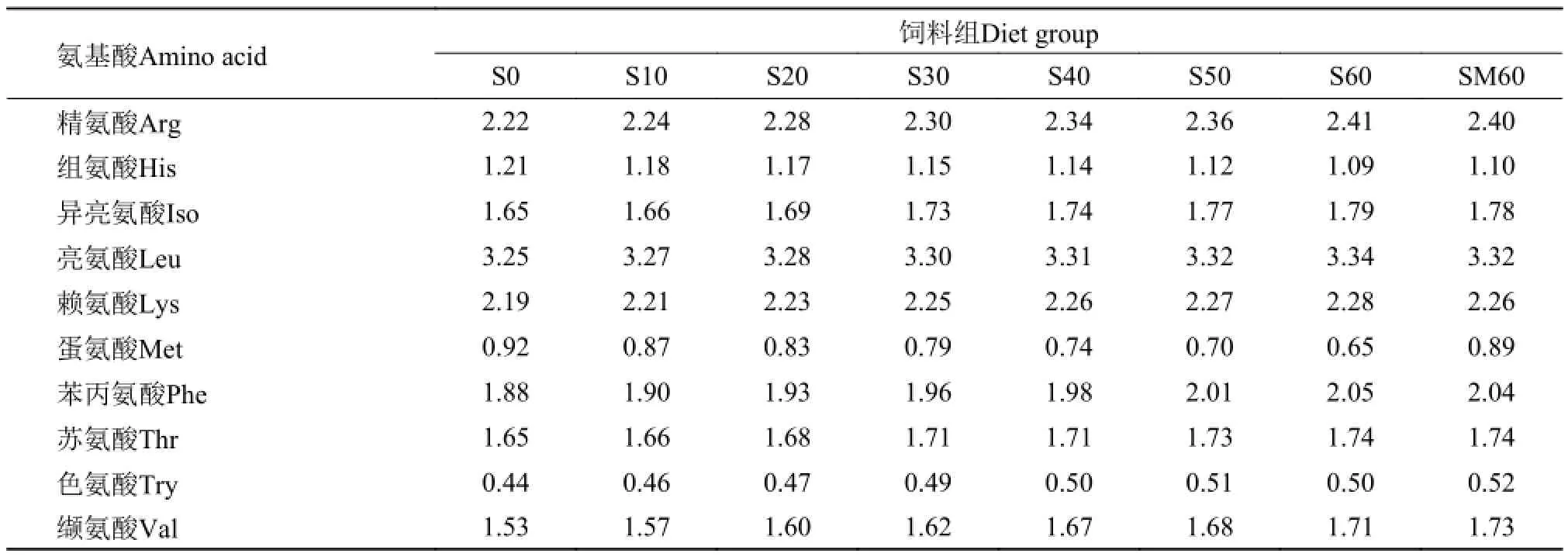
表 2 实验饲料的必需氨基酸组成(%干物质)Tab. 2 The composition of essential amino acids of diets (% dry weight)
1.2 试验鱼
本试验在东北农业大学水产养殖实验中心进行。乌苏里拟鲿稚鱼为当年人工孵化的同一批鱼苗。试验开始前, 用基础饲料对乌苏里拟鲿进行为期2周的驯养, 使之适应试验饲料和养殖环境。
在试验开始前, 试验鱼饥饿24h。挑选出大小一致、健康无病的稚鱼(0.50±0.00) g, 进行分组。实验在玻璃缸(1 m×0.5 m×0.5 m)中进行, 设8个处理, 每个处理设3个重复, 每个重复放50尾鱼。每天饱饲投喂2次(8:00和16:00), 定期清洗养殖缸, 随时检查鱼的生长情况。在整个养殖期间, 水的流量控制在0.3 L/min, 24h连续充氧, 保证溶氧≥6.0 mg/L,水温度为(23±1)℃, 氨氮<0.18 mg/L, pH为7.6—8.0, 光照周期为12h L/12h D。养殖试验持续8周。
1.3 样品收集
在试验开始前, 随机取50尾鱼, -20℃保存, 用于体组成分析。在试验结束后, 试验鱼饥饿24h。准确称每缸鱼总重, 记录鱼体总数; 每缸随机取5尾鱼, -20℃保存, 用于体组成分析; 每缸取5尾鱼, 测量体长和体重, 取内脏团和肝脏并称重; 取鱼体肌肉, -20℃保存, 用于肌肉组成分析。
在养殖试验结束后, 选择健康、规格一致的(3 g左右)稚鱼, 重新称重并分组。实验设计8个处理, 每个处理2个重复, 每重复放养30尾鱼, 并投喂含0.5%Cr2O3的饲料, 每天2次, 驯化1周后, 开始收集粪便。粪便的收集参照Lee[40]的方法。将收集的粪便-20℃保存。
1.4 分析方法
采用AOAC(1995)国际标准方法[12]分析饲料、全鱼和肌肉营养组成, 包括水分、粗蛋白、粗脂肪和灰分。饲料和粪便中Cr2O3和磷含量的测定采用容量分析法。
1.5 计算公式
实验指标的计算公式如下:
成活率(%)=100×(实验结束时尾数)/(实验开始时尾数)
增重率(WGR, %)= 100×(鱼终末体重-鱼初始体重)/鱼初始体重
特定生长率(SGR, %/d)=(Ln鱼终末体重-Ln鱼初始体重)×100/养殖周期
饲料系数(FCR)=实验期间投料量(g)/鱼净增重(g)
蛋白质效率(PER)=鱼净增重(g)/蛋白质摄取量(g)
肥满度(CF) =100×(体重, g)/(体长, cm)3
肝体比(HSI)=100×(肝重)/(体重)
内脏比(VSI)=100×(内脏重)/(体重)
表观消化率的计算公式如下:

式中: F是粪便中营养素含量, D是饲料中营养素含量, DCr是饲料中Cr2O3含量, FCr是粪便中Cr2O3含量。
1.6 统计分析
采用SAS 9.12统计软件对数据进行统计学分析。先作单因素方差分析(ANOVA), 若各处理之间差异显著, 再作Duncan's (1995)多重比较, P<0.05表示差异显著。试验结果采用“平均值±标准差”表示。
2 结果
2.1 豆粕替代鱼粉对生长和饲料效率的影响
试验结果表明, 乌苏里拟鲿的成活率较高(78.67%—86.67%), 各处理组之间差异不显著(P>0.05); 随着饲料中豆粕替代比例的增大, 鱼的末均重、增重率和特定生长率逐渐降低, 但10%—40%替代组与对照组之间差异不显著(P>0.05),50%和60%替代组显著低于对照组(P<0.05); 与S60相比, S60组添加晶体蛋氨酸后, 其增重率和特定生长率有所提高, 但差异不显著(P>0.05), 还是显著低于0—40%替代组(P<0.05); 当饲料中豆粕替代水平由0增加到20%时, 鱼的摄食率由3.22%提高到3.55%, 而后随着替代比例增加, 摄食率逐渐降低,但单因素方差分析表明差异不显著(表 3)。饲料系数随着豆粕替代比例的增大而逐渐增加(表 4), 其中, 60%替代组组显著高于对照组、10%和20%替代组(P<0.05), 与其他组之间差异不显著(P>0.05);随着豆粕替代比例的增大, 蛋白质效率表现出下降趋势, 其中, 60%替代组显著低于对照组、10%和20%替代组(P<0.05), 而与30%、40%和50%替代组差异不显著(P>0.05); 添加蛋氨酸组, 其蛋白质效率显著高于60%替代组(P<0.05), 而与其他组之间无显著差异(P>0.05)。这一结果表明, 豆粕可以替代乌苏里拟鲿饲料中40%鱼粉蛋白, 并没有对乌苏里拟鲿的生长和饲料利用产生不利影响。根据乌苏里拟鲿增重率和豆粕替代水平的关系, 经折线模型拟合得:y=-0.424x+427.3(R2=0.768), y=-2.560x+ 514.3(R2=0.975), 经计算得到当增重率达到最大时,豆粕替代水平为40.72%(图 1)。

表 3 不同豆粕替代量对乌苏里拟鲿生长的影响Tab. 3 The effect of different levels of SBM on growth of juvenile Pseudobagrus ussuriensis
2.2 豆粕替代鱼粉对体组成的影响
由表 5可知, 豆粕替代鱼粉对乌苏里拟鲿全鱼水分和灰分含量的影响不显著, 肌肉水分随着替代比例的升高而逐渐升高; 全鱼和肌肉粗蛋白含量随着替代比例的升高而逐渐降低, 其中, S60和SM60组显著低于其他组(P<0.05); 随着替代比例的升高, 全鱼和肌肉粗脂肪呈现升高的趋势, S60和SM60组显著高于其他替代组(P<0.05); 肥满度在各组间差异不显著; 当豆粕替代鱼粉蛋白比例为60% 时, 肝体比和脏体比指数显著高于其他组(P<0.05), 而其他组间差异不显著(表 6)。与SM60组间差异不显著(P>0.05)。
2.3 豆粕替代鱼粉对营养物质表观消化率的影响
由表 7可知, 当替代水平由0升高到40%时, 各实验组粗蛋白和干物质消化率差异不显著(P>0.05),50%和60%替代组的粗蛋白和干物质表观消化率显著降低(P<0.05); 随着替代水平的升高, 磷的表观消化率显著升高。豆粕替代比例对粗脂肪的表观消化率影响不显著(P>0.05); 添加晶体蛋氨酸能显著提高60%替代组粗蛋白消化率(P<0.05), 而对干物质、粗脂肪和磷的消化率无显著影响(P>0.05)。
3 讨论
3.1 豆粕替代鱼粉对生长和饲料效率的影响
在本实验中, 随着豆粕替代比例的增大, 试验鱼的生长和饲料效率逐渐降低, 其中, 10%—40%替代组与对照组之间差异不显著, 50%和60%组显著低于对照组; 这一结果表明, 豆粕替代乌苏里拟鲿饲料中40%鱼粉蛋白, 不会影响试验鱼的生长和饲料利用, 此替代比例下, 饲料中鱼粉和豆粕的含量分别为30%和28.4%。经折线模型拟合, 当增重率达到最大时, 豆粕替代水平为40.72%。本实验中, 豆粕替代鱼粉蛋白的比例同研究者对南方鲇(Silurusm eridionalis)(39%)[4]和军曹鱼(Rachycentron canadum)(40%)[6, 13]的研究结果相似, 却高于尖吻鲈(Lates calcarifer)(10%)[5]、鮸状黄姑鱼(Nibea miichthioides)(20%)[14]、金头鲷(Sparus aurata)(10%)[15]和许氏平鲉(Sebastes schlegeli)(20%—30%)[16]的研究结果, 低于尖头鲷(Diplodus puntazzo)(60%)[17]、南亚野鲮(Labeo rohita)(100%)[18]和蓝鲶鱼(Ictalurus furcatus)(70%—100%)[19]的研究结果。以上结果的差异可能是由于鱼的种类和大小、生理状态、饲料组成以及养殖条件差异。
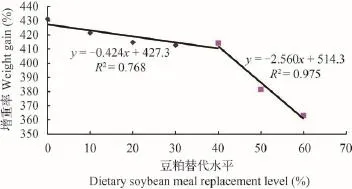
图 1 豆粕替代鱼粉与乌苏里拟鲿增重率的关系Fig. 1 The effects of different dietary soybean meal replacement level on weight gain of Pseudobagrus ussuriensis

表 4 不同豆粕替代量对乌苏里拟鲿饲料系数和蛋白质效率的影响Tab. 4 The effect of different levels of soybean meal on FCR and PER of juvenile Pseudobagrus ussuriensis
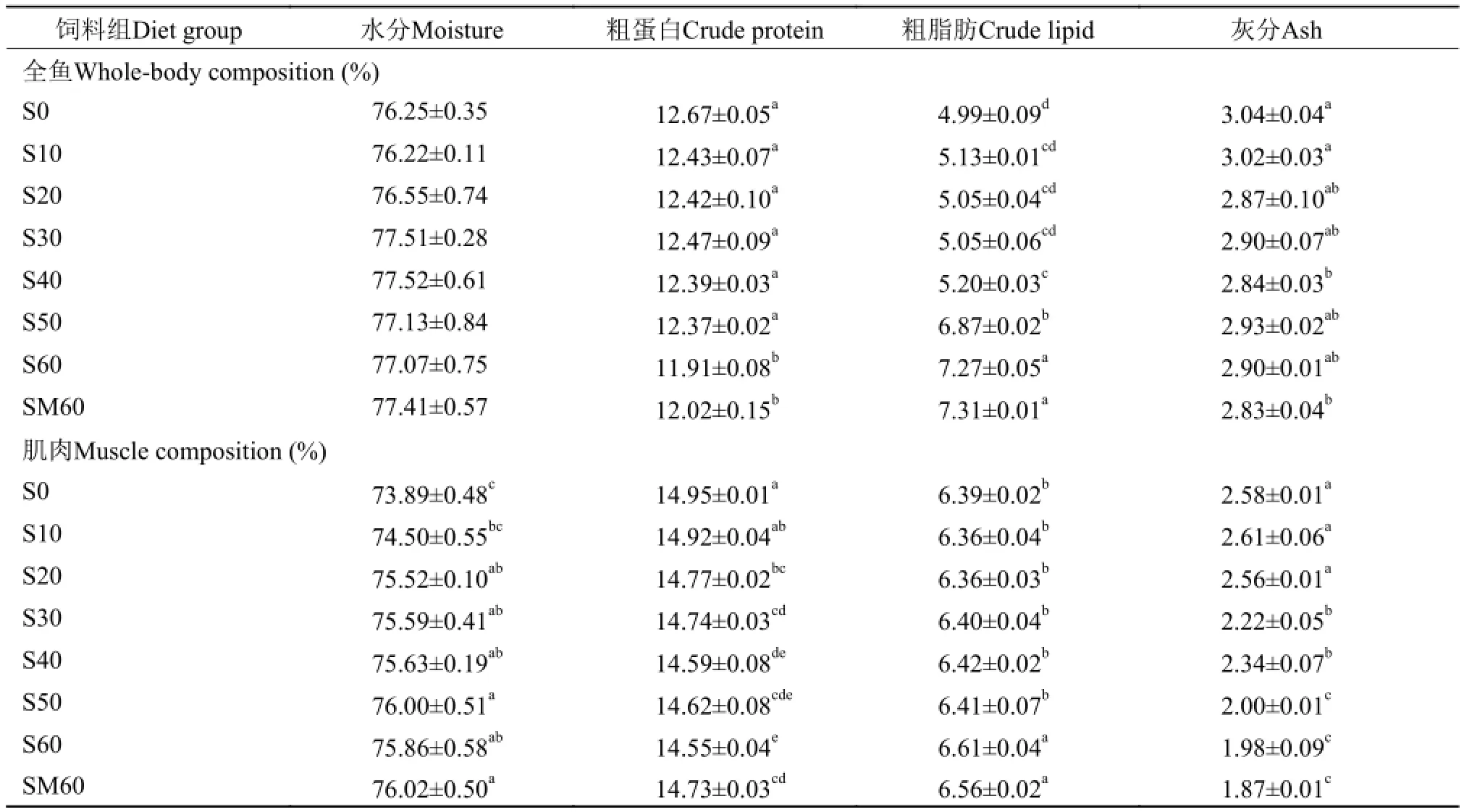
表 5 饲料中豆粕替代鱼粉对乌苏里拟鲿体成分的影响Tab. 5 Effects of dietary fish meal replacement by soybean meal on body composition of Pseudobagrus ussuriensis
在南方鲇[4]、鲈(Lates calcarifer)[5]、军曹鱼[13]、斑点叉尾鲖(Ictalurus punctatus)[20]和大西洋鲑(Salmo salar)[21]的实验中均发现, 饲料中豆粕含量较高时, 鱼类的生长和饲料利用显著降低, 这与本实验的研究结果趋势相一致。适口性差、抗营养因子的存在及必需氨基酸不平衡是导致高含量豆粕引起生长性能和饲料效率降低的原因。在本实验中, 当豆粕替代比例由0增加到20%时, 鱼的摄食率由3.22%提高到3.55%, 而后随着替代比例进一步增加, 摄食率逐渐降低, 但方差分析表明, 差异不显著, 这表明高比例替代组乌苏里拟鲿的生长降低与饲料的适口性无关。一些学者在对南方鲇[4]、虹鳟(Oncorhynchus mykiss)[7]、军曹鱼[13]、金头鲷[15]和南亚野鲮[18]的研究中均发现, 饲料中添加适量大豆蛋白, 并没有引起适口性的变化, 这与本研究结果类似。然而, 对南方鲇[4]、尖吻鲈[5]和军曹鱼[6]研究结果表明, 饲料中高含量的豆粕会引起摄食率下降, 这可能是因为大豆蛋白中存在胰蛋白酶抑制因子(TIA)等抗营养因子, 会抑制鱼类的食欲。有关豆粕中抗营养因子对乌苏里拟鲿生长和饲料利用的影响还有待于进一步研究。
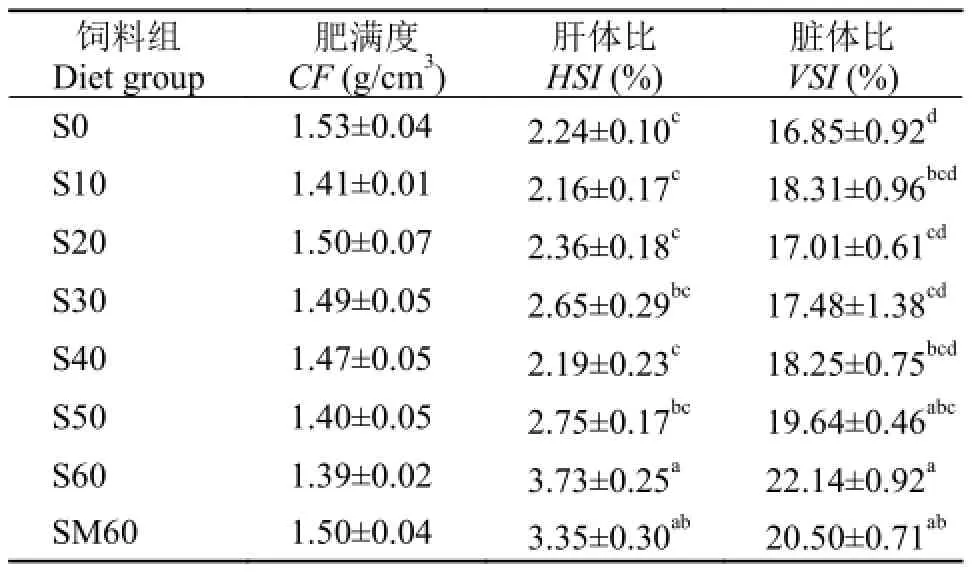
表 6 饲料中豆粕替代鱼粉对乌苏里拟鲿肝体指数的影响Tab. 6 Effect of dietary fish meal replacement by soybean meal on Pseudobagrus ussuriensis somatic parameters (dry weight basis)

表 7 豆粕替代鱼粉蛋白对乌苏里拟鲿干物质、粗蛋白、粗脂肪和磷表观消化率的影响Tab. 7 Apparent digestibility coefficients for dry matter, crude protein, crude lipid and phosphorus of the Pseudobagrus ussuriensis
蛋氨酸是豆粕型饲料的第一限制性氨基酸。当大豆蛋白的添加量过高时, 饲料氨基酸的不平衡性表现的越明显, 对生长的抑制作用也越强烈[4, 13]。过40%时, 乌苏里拟鲿的生长和饲料效率显著降低,经过分析得出, 随着豆粕替代比例由0升高到60%,饲料中蛋氨酸含量由0.92%减少至0.65%, 蛋氨酸含量降低29.35%, 蛋氨酸含量不足导致的必需氨基酸模式不平衡可能是乌苏里拟鲿生长和饲料利用率降低的原因。研究者对军曹鱼[6, 13]、黄姑鱼[14]和虹鳟[22]研究中也得出了类似的结论。
饲料中含量不足的必需氨基酸的添加对提高植物蛋白源的营养价值、促进鱼类生长是必要的[19, 23]。研究者在对许氏平鲉的试验中发现, 不添加蛋氨酸时, 豆粕可替代20%的鱼粉蛋白, 而添加蛋氨酸后,可替代30%的鱼粉蛋白而对生长无不良影响[16];Médale等[22]的研究结果表明: 蛋氨酸缺乏导致虹鳟摄食量和生长降低, 补充晶体蛋氨酸则提高摄食率和生长。本实验中, 与60%替代相比较, 添加0.3%蛋氨酸组鱼的生长性能和饲料效率有所提高,但是促生长效果不明显, 研究者对虹鳟[24]的研究中也得到了类似的结果。这可能是由于存在晶体氨基酸溶失及其在鱼类肠道中与结合氨基酸吸收不同步等问题, 可以通过增加投喂次数和包被氨基酸来解决这些问题[25]。
3.2 豆粕替代鱼粉对体组成和形态指数的影响
不同的研究者对豆粕替代鱼粉后对鱼体成分的影响的结论不尽相同, 可能与鱼的种类、饲喂方法、蛋白原料特性和饲料组成有关[41]。本研究的结果表明, 豆粕替代鱼粉对乌苏里拟鲿全鱼水分和灰分含量的影响不显著, 而随着替代比例的升高,肌肉水分逐渐升高; 全鱼和肌肉粗蛋白含量随着替代比例的升高而逐渐降低, 其中, S60和SM60组显著低于其他组, 这与研究者对军曹鱼[13]和大西洋鲑[21]的研究结果一致, 但与一些研究结果趋势相反[5, 16]。高比例的豆粕引起鱼体蛋白含量降低, 可能是因为饲料中豆粕含量太高时, 蛋氨酸含量降低, 使得饲料中必需氨基酸比例失衡, 导致蛋白质合成受阻[26]。随着豆粕替代鱼粉比例的升高, 全鱼和肌肉粗脂肪含量呈现升高的趋势, 其中, S60组显著高于其他替代组, 这与许多替代性研究结果趋势相一致[13, 27],这可能是由于蛋氨酸含量降低, 减缓了肉碱的合成,进而抑制了脂肪酸的β-氧化, 使鱼体组织脂肪含量上升。
在本试验中, 在0—50%替代组, 肝体比和脏体比指数差异不显著, 但是当豆粕替代鱼粉比例为60%时, 肝体比和脏体比指数显著升高, 这表明高比例豆粕替代鱼粉会影响肝脏的形态。研究者在斑点叉尾鲖[28]、虹鳟[29]和笛鲷(Lutjanus argentiventris)[30]等鱼类的研究中也发现, 饲喂过量大豆蛋白会导致肝胰脏指数升高。据报道, 饲喂生大豆组小鼠的肝胰脏重量显著高于去除TIA组, 且胰脏的重量随着TIA含量的增加而增加。TIA会诱导肝胰脏分泌更多的胰蛋白酶补充至肠道中, 达到消化蛋白质的目的, 结果造成胰蛋白酶原的大量消耗, 引起含硫氨基酸内源流失, 同时引起胰腺的肥大[31]。 研究发现, 向虹鳟[32]和罗非鱼(O. niloticus×O. Aureus)[33]饲料中添加一定量的蛋氨酸可以明显降低肝胰脏的脂质积累, 在本实验中, 向60%组添加0.3%蛋氨酸可以降低肝体比和脏体比指数, 但是降低效果不明显, 这可能与添加量以及对晶体氨基酸的利用率有关。
3.3 豆粕替代鱼粉对乌苏里拟鲿营养物质表观消化率的影响
本研究结果表明, 当替代水平由0升高到40%时, 各实验组干物质和蛋白质消化率差异不显著,50%和60%组的消化率显著降低, 这与大多数有关大豆蛋白源替代鱼粉对营养物质消化率的影响规律一致[4, 17, 23, 34]。饲料中过量添加大豆蛋白导致消化率降低, 这主要与抗营养因子和氨基酸不平衡有关[4, 21, 23, 36]。大豆蛋白中抗营养因子改变了消化道的黏膜结构, 引起非感染性亚急性肠炎, 从而导致营养物质消化率的降低[21, 35, 36]。本研究关于肠道的变化需通过组织学方法进一步研究。还有研究认为, 大豆蛋白产品中可溶性的非淀粉多糖对消化道有损伤作用, 导致营养物质在肠道中的转运能力降低, 甚至在消化道中形成胶团物质, 从而阻止了养分与消化酶的进一步接触[21, 37]。鱼类饲料原料的磷表观消化率变化较大。在本实验中, 随着豆粕替代比例由0升高到60%, 磷表观消化率由36.07%升高到66.50%, 高于一些实验结果[38, 39]。有关豆粕替代鱼粉对鱼类营养物质的消化率的影响结果差异较大, 这可能与鱼的种类、生理状态、饲料组成、大豆产品种类、投喂方法、粪便收集方法和指示剂种类等有关。
4 结论
当饲料中豆粕替代鱼粉蛋白的比例低于40%时, 乌苏里拟鲿的生长、饲料转化率、鱼体组成和消化率均不受到显著影响。而高比例豆粕替代会影响鱼类的生长和肌体健康。
[1]Zhou Q C, Mai K S, Liu Y J, et al. Advances in animal and plant protein sources in place of fish meal [J]. Journal of Fisheries of China, 2005, 29(3): 404—410 [周歧存, 麦康森, 刘永坚, 等. 动植物蛋白源替代鱼粉研究进展. 水产学报, 2005, 29(3): 404—410]
[2]Tacon A G J, Metian M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects [J]. Aquaculture,2008, 285(1—4):146—158
[3]New M B. Use of fishmeal and fish oil in aquafeeds: further thoughts on the fishmeal trap. Food and Agriculture Organization of the United Nations. Rome. 2002
[4]Ai Q H, Xie X J. Effects of replacement of fish meal by soybean meal and supplementation of methionine in fish meal/soybean meal-based diets on growth performance of the Southern catfish Silurusm eridionalis [J]. Journal of the World Aquaculture Society, 2005, 36(4): 498—507
[5]Tantikitti C, Sangpong W, Chiavareesajja S. Effects of defatted soybean protein levels on growth performance and nitrogen and phosphorus excretion in Asian seabass (Lates calcarifer) [J]. Aquaculture, 2005, 248(1—4):41—50
[6]Zhou Q C, Mai K S, Tan B P, et al. Partial replacement of fishmeal by soybean meal in diets for juvenile cobia (Rachycentron canadum) [J]. Aquaculture Nutrition,2005, 11(3): 175—182
[7]Ostaszewska T, Dabrowski K, Palacios M E, et al. Growth and morphological changes in the digestive tract of rainbow trout (Oncorhynchus mykiss) and pacu (Piaractus mesopotamicus) due to casein replacement with soybean proteins [J]. Aquaculture, 2005, 245(1—4):273—286
[8]Zhang J M. The Fishes of Heilongjiang [M]. Harbin: Heilongjiang Science and Technology Press. 1995, 205—207[张觉民. 黑龙江省鱼类志. 哈尔滨: 黑龙江科学技术出版社. 1995, 205—207]
[9]Yin H B, Sun Z W, Yu B. Ultrastructure of Spermatozoon of Pseudobagrus ussuriensis [J]. Journal of Guangdong Ocean University, 2009, 29(6): 28—31 [尹洪滨, 孙中武, 于波. 乌苏里拟鲿(Pseudobagrus ussuriensis)精子的超微结构. 广东海洋大学学报, 2009, 29(6): 28—31]
[10]Pan W Z, Wang P, Zhao C G, et al. Preliminary study on propagationbiology and artificial breeding of Pseudobagrus ussuriensis [J]. Chinese Journal Fisheries, 2001,14(2): 1—3 [潘伟志, 王鹏, 赵春刚, 等. 乌苏里拟鲿人工繁育技术及开发利用—乌苏里拟鲿繁殖生物学及人工催产初步研究. 水产学杂志, 2001, 14(2): 1—3]
[11]Pan W Z, Yin H B, Sun Z W, et al. Isozyme of Pseudobagrus ussuriensis in Heilongjiang water system [J]. Journal of Northeast Forestry University, 2006, 34(6): 66—69[潘伟志, 尹洪滨, 孙中武, 等. 乌苏里拟鲿(Pseudoba-grus ussuriensis)同工酶分析. 东北林业大学学报, 2006,34(6): 66—69]
[12]AOAC. Official Methods of Analysis of Official Analytical Chemists International [M]. 16th edn. Association of Official Analytical Chemists, Arlington, VA. 1995
[13]Chou R L, Her B Y, Su M S, et al. Substituting fish meal with soybean meal in diets of juvenile cobia Rachycentron canadum [J]. Aquaculture, 2004, 229(1—4):325—333
[14]Wang Y, Kong L J, Li C, et al. Effect of replacing fish meal with soybean meal on growth, feed utilization and carcass composition of cuneate drum (Nibea miichthioides) [J]. Aquaculture, 2006, 261(4): 1307—1313
[15]Venou B, Alexis M N, Fountoulaki E, et al. Effects of extrusion and inclusion level of soybean meal on diet digestibility, performance and nutrient utilization of gilthead sea bream (Sparus aurata) [J]. Aquaculture, 2006, 261(1):343—356
[16]Lim S R, Choi S M, Wang X J, et al. Effects of dehulled soybean meal as a fish meal replacer in diets for fingerling and growing Korean rockfish Sebastes schlegeli [J]. Aquaculture, 2004, 231(1—4): 457—468
[17]Hernández M D, Martínez F J, Jover M, et al. Effects of partial replacement of fish meal by soybean meal in sharpsnout seabream (Diplodus puntazzo) diet [J]. Aquaculture, 2007, 263(1—4): 159—167
[18]Khan M A, Jafri A K, Chadha N K, et al. Growth and body composition of rohu (Labeo rohita) fed diets containing oilseed meals: partial or total replacement of fish meal with soybean meal [J]. Aquaculture Nutrition, 2003,9(6): 391—396
[19]Webster C D, Goodgame-Tiu L S, Tidwell J H. Total replacement fish meal by soybean meal, with various percentages of supplemental L-methionine, in fish diets for blue catfish, Ictalurus furcatus (Leseur) [J]. Aquaculture Research, 1995, 26(5): 299—306
[20]Mohsen A A, Lovell R T. Partial substitution of soybean meal with animal protein sources in diets for channel catfish [J]. Aquaculture, 1990, 90(3—4): 303—311
[21]Refstie S, Storebakken T, Baeverfjord G, et al. Long-term protein and lipid growth of Atlantic salmon (Salmo salar)fed diets with partial replacement of fish meal by soy protein products at medium or high lipid level [J]. Aquaculture, 2001, 193(1—2): 91—106
[22]Medale F, Boujard T, Vallee F, et al. Voluntary feed intake, nitrogen and phosphorus losses in rainbow trout (Oncorhynchus mykiss) fed increasing dietary levels of soy protein concentrate [J]. Aquatic Living Resource,1998, 11(4): 239—246
[23]Deng J M, Mai K S, Ai Q H, et al. Effects of replacing fish meal with soy protein concentrate on feed intake and growth of juvenile Japanese flounder, Paralichthys olivaceus [J]. Aquaculture, 2006, 258(1—4): 503—513
[24]Schuhmacher A, Wax C, Gropp J. Plasma amino acids in rainbow trout (Oncorhynchus mykiss) fed intact protein or a crystalline amino acid diet [J]. Aquaculture, 1997,151(1—4): 15—28
[25]Segovia-Quintero M A, Reigh R C. Coating crystalline methionine with tripalmitin-polyvinyl alcohol slows its absorption in the intestine of Nile tilapia, Oreochromis niloticus [J]. Aquaculture, 2004, 238(1—4): 355—367
[26]Langar H, Guillaume J, Métailler R, et al. Augmentation of protein synthesis and degradation by poor dietary amino acid balance in European sea bass (Dicentrarchus labrax) [J]. Journal of Nutrition, 1993, 123(10):1754—1761
[27]Refstie S, Sahlstrom S, Brathen E, et al. Lactic acid fermentation eliminates indigestible carbohydrates and antinutritional factors in soybean meal for Atlantic salmon (Salmo salar) [J]. Aquaculture, 2005, 246(1—4):331—345
[28]Peres H, Lim C, Klesius P H. Nutritional value of heattreated soybean meal for channel catfish (Ictalurus punctatus) [J]. Aquaculture, 2003, 225(1—4): 67—82
[29]Oliva-Teles A, Gouveia A, Gomes E, et al. The effect of different processing treatment on soybean meal utilization by rainbow trout [J]. Aquaculture, 1994, 124(1—4):343—349
[30]Alarcon F L, Garcia-Carreno, Navarrete D A. Effects of plant protease inhibitors on digestive proteases in two fish species, Lutjanus argentiventris and L. novemfasciatus [J]. Fish Physiology and Biochemistry, 2001, 24(3):179—189
[31]Struthers B J, Macdonald J R, Dahlgren R R, et al. Effects on the monkey, pig, and rat pancreas of soy products with varying levels of trypsin inhibitor and comparison with the administration of cholecystokinin [J]. Journal of Nutrition, 1983, 113(1): 86—97
[32]Yokoyama M, Nakazoe J. Utilization of methionine supplemented to diet in rainbow trout [J]. Nippon Suisan Gakkaishi, 1992, 58(7): 1347—1349
[33]Lin S M, Mai K S, Tan B P. Influence of practical diet supplementation with freed-methionine on the growth and body composition in tilapia Oreochromis niloticus×O. aureus [J]. Acta Hydrobiologica Sinica, 2008, 32(5):741—749 [林仕梅, 麦康森, 谭北平. 实用饲料中添加结晶蛋氨酸对罗非鱼生长、体组成的影响. 水生生物学报, 2008, 32(5): 741—749]
[34]Koprucu K, Ozdemir Y. Apparent digestibility of selected feed ingredients Nile tilapia (Oreochromis niloticus)[J]. Aquaculture, 2005, 250(1—2): 308—316
[35]Opstvedt J, Aksnes A, Hope B, et al. Efficiency of feed utilization in Atlantic salmon (Salmo salar L.) fed diets with increasing substitution of fish meal with vegetable proteins [J]. Aquaculture, 2003, 221(1—4): 365—379
[36]Mundheim H, Aksnes A, Hope B. Growth, feed effi-ciency and digestibility in salmon (Salmo salar L.) fed different dietary proportions of vegetable protein sources in combination with two fish meal qualities [J]. Aquaculture, 2004, 237(1—4): 315—331
[37]Refstie S, Storebakken T, Roem A J. Feed consumption and conversion in Atlantic salmon (Salmo salar) fed diets with fish meal, extracted soybean meal or soybean meal with reduced content of oligosaccharides, trypsin inhibitors, lectins and soy antigens [J]. Aquaculture, 1998,162(3—4): 301—312
[38]Vielma J, Makinenj T, Ekholm P, et al. Influence of dietary soy and phytase levels on performance and body composition of large rainbow trout Oncorhynchus mykiss and algal availability of phosphorus load [J]. Aquaculture, 2000, 183(3—4): 349—362
[39]Sajjadi M, Carter C G. Dietary phytase supplementation and the utilization of phosphorus by Atlantic salmon (Salmo salar L.) fed a canola-meal-based diet [J]. Aquaculture Nutrition, 2004, 240(1—4): 417-431
[40]Lee S M. Apparent digestibility coefficients of various feeding redients for juvenile and grower rockfish (Sebastes schlegeli) [J]. Aquaculture, 2002, 207(1—2):79—95
[41]Zeitler M H, Kirchgessner M, Schwarz F J. Effects of different protein and energy supplies on carcass composition of carp (Cyprinus carpio L.) [J]. Aquaculture, 1984,36(1): 37—48
EFFECTS OF REPLACING FISH MEAL WITH SOYBEAN MEAL ON GROWTH PERFORMANCE, BODY COMPOSITION AND APPARENT DIGESTIBILITY COEFFICIENTS OF PSEUDOBAGRUS USSURIENSIS
WANG Yu-Yu1, ZHOU Qi-Cun2, BU Xian-Yong1, SU Bao-Hui1and YANG Yu-Hong1
(1. College of Animal Science and Technology, Northeast Agricultural University, Harbin 150030, China; 2. Laboratory of Fish Nutrition, School of Marine Sciences, Ningbo University, Ningbo 315211, China)
An 8-week feeding trial was conducted in indoor culture system to evaluate effects of replacing fish meal with soybean meal (SBM) on growth performance, body composition and apparent digestibility coefficients of juvenile Pseudobagrus ussuriensis [initial body weight of (0.50±0.00) g]. Seven isonitrogenous diets groups were formulated to replace fish meal protein by SBM at 0 (S0), 10% (S10), 20% (S20), 30% (S30), 40% (S40), 50% (S50) and 60% (S60). To investigate the effect of supplementation with methionine, S60 added 0.3% methionine to formulated the 8thgroup (SM60). Weight gain rate (WGR), specific growth rate (SGR) and protein efficiency ratio (PER) linearly decreased with increasing SBM replacement level from 0 to 60%. There were no significant differences in WGR, SGR and PER among groups of S0, S10, S20, S30, and S40 (P>0.05), but S50 and S60 groups had lower WGR, SGR and PER than S0 group (P<0.05). No significant differences were observed in growth between S60 group and SM60 (P>0.05) group, but both of them had significantly lower growth than S0 group. Feed conversion ratio (FCR) had an opposite trend with WGR and SGR. S0 group had significantly lower muscle moisture content than other groups (P<0.05). Whole body and muscle protein contents decreased with increasing dietary SBM levels, and S60 and SM60 groups had significantly lower protein content than other groups (P<0.05). Whole body and muscle lipid contents increased with increasing dietary SBM levels, and S60 and SM60 groups had significantly higher lipid contents than other groups (P<0.05). S60 and SM60 groups had higher VSI and HSI than other groups (P<0.05). Apparent digestibility coefficients (ADC) of protein and dry matter of S50 and S60 groups were significantly lower than other groups (P<0.05). The ADC of phosphorus significantly increased with increasing SBM levels in the diet (P<0.05). These results suggest that SBM at not more than 40% of fish meal protein replacement have no significantly negative effect on growth or feed efficiency, but higher dietary SBM levels reduce fish performance.
Pseudobagrus ussuriensis; Fish meal; Soybean meal; Growth; Apparent digestibility coefficients
S965.1
A
1000-3207(2016)04-0681-09
10.7541/2016.91
2015-09-18;
2016-01-15
宁波大学浙江省重中之重学科开放基金(xkzsc1510)资助 [Supported by the Open Fund of Zhejiang Provincial Top Key Discipline of Aquaculture in Ningbo University (xkzsc1510)]
王裕玉(1983—), 男, 山东日照人; 博士研究生; 研究方向为水产动物营养与饲料学。E-mail: wangyuyu1983010@163. com
杨雨虹, E-mail: yuhongyang68@163. com
猜你喜欢
当代水产(2022年8期)2022-09-20 06:46:06
当代水产(2022年6期)2022-06-29 01:13:12
延边大学学报(社会科学版)(2022年2期)2022-04-26 14:12:04
——乌苏里白鲑
中国水产(2022年3期)2022-03-23 04:40:36
当代水产(2019年6期)2019-07-25 07:52:08
当代水产(2019年1期)2019-05-16 02:41:50
民族音乐(2018年5期)2018-01-24 11:26:33
广东饲料(2016年3期)2016-12-01 03:43:11
现代农业(2015年5期)2015-02-28 18:41:02
中国畜牧业(2014年4期)2014-10-16 03:50:40
