
苦丁冬青苦丁茶咖啡酰奎尼酸类物质与a-淀粉酶的相互作用特性
2016-08-10 07:24徐冬兰王晴川曾晓雄南京农业大学食品科技学院江苏南京210095
食品科学 2016年13期
徐冬兰,王晴川,曾晓雄,孙 怡*(南京农业大学食品科技学院,江苏 南京 210095)
苦丁冬青苦丁茶咖啡酰奎尼酸类物质与a-淀粉酶的相互作用特性
徐冬兰,王晴川,曾晓雄,孙 怡*
(南京农业大学食品科技学院,江苏 南京 210095)
摘 要:探讨苦丁冬青苦丁茶中咖啡酰奎尼酸(caffeoylquinic acids,CQA):3-CQA、4-CQA、5-CQA、3,4-diCQA、3,5-diCQA和4,5-diCQA对α-淀粉酶的体外抑制活性,并采用荧光光谱法和圆二色谱法分析CQA与α-淀粉酶的相互作用特性,使用修正后的Stern-Volmer方程与van't Hoff方程探讨CQA-酶之间的结合常数、结合位点数及热力学参数。结果表明:3-CQA、4-CQA、5-CQA、3,4-diCQA、3,5-diCQA和4,5-diCQA对α-淀粉酶均有较强的抑制作用,半抑制浓度(IC50)分别为1.54、1.05、1.28、0.96、0.33、0.64 mg/mL;CQA与α-淀粉酶发生结合反应,生成稳定的复合物,从而造成酶分子内部荧光发生猝灭;热力学参数分析表明CQA与α-淀粉酶主要靠疏水作用力结合,反应自发进行。此外,圆二色谱结果显示CQA的结合引起α-淀粉酶二级结构的变化,破坏了酶蛋白的天然构象,从而降低了酶的催化活性。
关键词:α-淀粉酶;咖啡酰奎尼酸;荧光光谱;圆二色谱;相互作用
引文格式:
徐冬兰, 王晴川, 曾晓雄, 等.苦丁冬青苦丁茶咖啡酰奎尼酸类物质与α-淀粉酶的相互作用特性[J].食品科学, 2016,37(13): 6-12.DOI:10.7506/spkx1002-6630-201613002. http://www.spkx.net.cn
XU DongLan, WANG Qingchuan, ZENG Xiaoxiong, et al.Interaction properties of caffeoylquinic acid derivatives from Ilex kudingcha C.J.Tseng with α-amylase[J].Food Science, 2016, 37(13): 6-12.(in Chinese with English abstract)
随着现代生活水平的提高和饮食习惯的改变,2型糖尿病和肥胖症的发病率日益增高,其预防和治疗方法成为了人们关注的焦点。已有研究表明餐后血糖浓度过高成为2型糖尿病和肥胖症发生的重要因素[1-2]。胰α-淀粉酶(EC 3.2.1.1)作用于淀粉或糖原分子内部的α-1,4-糖苷键,最终生成麦芽糖、低聚糖、糊精和少量葡萄糖,是消化系统中的一个关键酶[3-4]。抑制α-淀粉酶的活性可以减缓淀粉的水解过程,从而控制餐后血糖水平的升高[5-6]。因此,有效和无毒的α-淀粉酶抑制剂对于2型糖尿病和肥胖的预防和治疗具有重要的意义。
近年来,多酚类物质对α-淀粉酶的抑制活性被广泛探讨。Fei Qunqin等[7]利用酶抑制动力学探讨了乌龙茶多酚及其单体表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)、甲基化EGCG(EGCG3''Me)对胰α-淀粉酶体外活性的影响,表明乌龙茶多酚对胰α-淀粉酶具有显著的抑制效果,抑制类型为竞争性抑制。Miao Ming等[8]的研究表明葡萄皮中的白藜芦醇具有较强的α-淀粉酶抑制活性,同时荧光实验发现白藜芦醇能以静态猝灭的方式与酶发生结合。阮妙芸等[9]采用体外模拟淀粉消化实验表明茶多酚在低浓度下便能对淀粉酶具有明显的抑制作用。
苦丁茶是我国南部和东部地区居民长期饮用的一种代用茶,也是传统的药用植物,其富含多酚类、萜类、黄酮类、多糖等活性物质,具有抗氧化、降血压以及降脂减肥等功效[10-12]。本课题组前期建立了苦丁茶中多酚类物质的分离、纯化以及高效液相色谱分析方法,表明苦丁茶中的主要多酚类物质为咖啡酰奎尼酸(caffeoylquinic acids,CQA)类衍生物,并通过柱层析和半制备色谱制备得到了6 种物质,包括3-CQA、4-CQA、5-CQA、3,4-diCQA、3,5-diCQA和4,5-diCQA[13-15]。CQA类衍生物被证明具有较强的α-淀粉酶抑制活性[16],然而关于其与α-淀粉酶的相互结合作用机理却未见报道。因此,在前期的研究基础上,本实验拟采用荧光光谱法研究CQA与α-淀粉酶的相互作用特性,为阐明CQA对α-淀粉酶的抑制机理提供信息,并为天然α-淀粉酶抑制剂的开发提供理论基础。
1 材料与方法
1.1 材料与试剂
咖啡酰奎尼酸:3-CQA、4-CQA、5-CQA、3,4-diCQA、3,5-diCQA和4,5-diCQA,按照食品与纳米生物技术实验室之前报道的方法自制[13-15],纯度均大于95%。
淀粉天青、猪胰α-淀粉酶、Tris-HCl缓冲溶液(pH 6.9、0.05 mol/L)、醋酸 美国Sigma公司;其他试剂均为分析纯。
1.2 仪器与设备
BL-220H分析天平 日本Shimadzu公司;DELTA 320 pH计 德国Mettler-Toledo公司;Genius 3旋涡混匀器德国IKA公司;TDL-5型台式离心机 上海安亭科学仪器厂;Synergy-2酶标仪 美国Biotek公司;F-7000型荧光分光光度计 日本Hitachi公司;HH-4数显恒温水浴锅江苏国华电器有限公司。
1.3 方法
1.3.1 CQA对α-淀粉酶活性的抑制作用
参照Hansawasdi等[17]的方法并稍作修改:用0.05 mol/L Tris-HCl缓冲溶液配制质量浓度为10 mg/mL的淀粉天青底物溶液,并于沸水浴中助溶5 min,之后置于37 ℃条件下预热备用。将0.2 mL不同质量浓度的样品溶液(0.1、0.2、0.4、0.6、0.8 mg/mL)分别与0.1 mL α-淀粉酶溶液(2.2 U/mL)充分混合,37 ℃预热10 min后加入0.2 mL底物溶液,充分混匀后于37 ℃反应10 min。最后各加入0.5 mL 50%醋酸溶液终止反应。将反应混合液于4 ℃、3 000 r/min离心5 min,取上清液,测定其在595 nm波长处的吸光度。每组实验重复3 次,按下式计算不同样品对α-淀粉酶的抑制率。

式中:Ac+为不加样品但含酶的对照组在595 nm波长处的吸光度;Ac-为不加样品也不含酶的空白组在595 nm波长处的吸光度;As为既加样品也含酶的样品组在595 nm波长处的吸光度;Ab为加入样品但未加酶的样品对照组在595 nm波长处的吸光度。
1.3.2 CQA与α-淀粉酶相互作用特性的荧光光谱法分析
荧光猝灭作用:将3.0 mL 0.05 mg/mL的α-淀粉酶溶液准确移入离心管中,用移液枪分别加入不同的样品溶液(5-CQA、3,4-diCQA、3,5-diCQA或4,5-diCQA),使其终质量浓度为0.005、0.010、0.015、0.020、0.025 mg/mL,在设定温度(293、310 K)条件下于恒温水浴中保温0.5 h。使用1.0 cm的石英比色皿,固定激发波长(λex)为280 nm、发射波长(λem)290~450 nm,激发和发射狭缝宽度均为5.0 nm,扫描速率1 200 nm/min,在荧光分光光度计上恒温扫描α-淀粉酶及反应混合物的荧光光谱。
结合常数及热力学相关参数的计算:固定激发波长为280 nm,发射波长为340 nm,记录α-淀粉酶及反应混合物在293、310 K温度条件下的荧光强度,所得实验数据用于结合常数和相关热力学参数的计算。
3D荧光光谱分析:设定激发波长与发射波长的扫描范围均为200~600 nm,狭缝宽度为5 nm,扫描α-淀粉酶及反应混合物的3D图谱。
1.3.3 圆二色谱(circular dichroism,CD)扫描
α-淀粉酶及其与样品反应混合液的CD光谱在Jasco-810圆二色谱仪上进行。用缓冲液配制α-淀粉酶与样品体系,使体系中蛋白质质量浓度始终固定为0.5 mg/mL,而样品(5-CQA、3,5-diCQA)质量浓度分别为0.01、0.02 mg/mL。在室温条件下反应0.5 h后,分别测定α-淀粉酶原液及反应混合液在200~250 nm波长范围内远紫外CD光谱。每个实验样品重复扫描3 次。采用CDpro软件中Selcon3算法分析取得的CD数据,得到关于α-淀粉酶二级结构的信息。
2 结果与分析
2.1 CQA对α-淀粉酶的抑制作用
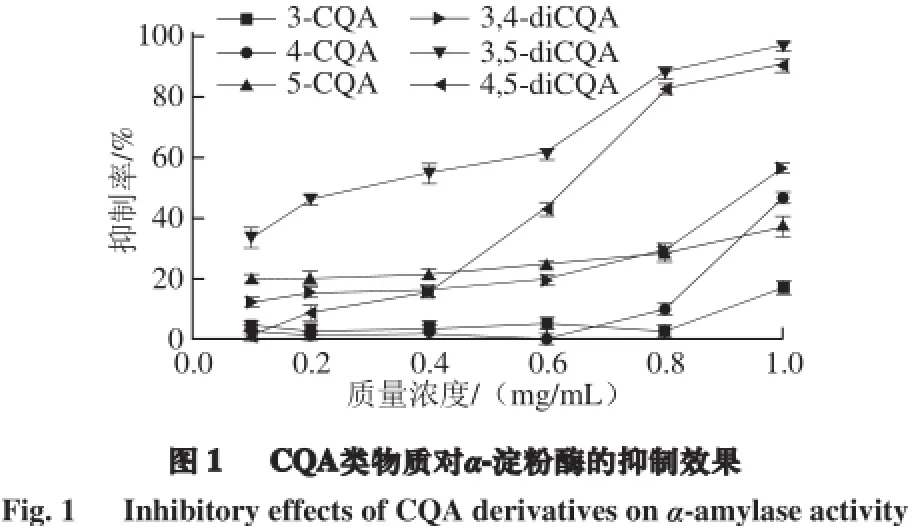
以样品质量浓度为横坐标,抑制率为纵坐标,绘制抑制曲线,结果如图1所示,随着样品质量浓度的增大,抑制率明显升高,当3,5-diCQA的质量浓度为1.0 mg/mL时,抑制率接近100%,表明6 种CQA对α-淀粉酶的活性均有较强的抑制作用。通过线性拟合计算出不同样品对α-淀粉酶的半抑制浓度(IC50),结果显示3-CQA、4-CQA、5-CQA、3,4-diCQA、3,5-diCQA和4,5-diCQA的IC50分别为1.54、1.05、1.28、0.96、0.33、0.64 mg/mL,抑制能力由强到弱的顺序为:3,5-diCQA>4,5-diCQA>3,4-diCQA>4-CQA>5-CQA>3-CQA。双取代咖啡酰奎尼酸的抑制效果明显优于单取代咖啡酰奎尼酸,咖啡酰基增多,抑制活性增强,表明结构中的咖啡酰基是主要的活性基团,对抑制活性贡献最大。
2.2 CQA对α-淀粉酶荧光光谱的猝灭效应
药物小分子与蛋白质的相互结合作用通常会降低蛋白质内部的荧光强度,称为荧光猝灭效应[18]。胰α-淀粉酶由496 个氨基酸残基组成,包括17 个色氨酸,因此在激发波长280 nm处能够检测到内源性荧光[19-20]。实验中固定激发波长为280 nm,扫描α-淀粉酶及CQA与α-淀粉酶反应混合物在300~450 nm波长范围内的发射光谱,结果如图2所示。随着4 种CQA质量浓度不断增大,α-淀粉酶的荧光强度逐渐降低,荧光光谱发生明显猝灭。此外,α-淀粉酶在340 nm波长处具有最大荧光发射峰,随着CQA的加入,最大发射波长发生明显红移(5-CQA,从340 nm移至360 nm;3,4-diCQA,从340 nm移至358 nm;3,5-diCQA,从340 nm移至362 nm; 4,5-diCQA,从340 nm移至365 nm)。这些结果表明CQA小分子与α-淀粉酶之间发生了结合作用,形成了复合物,使得酶蛋白内部结构发生改变,发色团所处微环境的极性增强,疏水性发生变化[21],最终改变了α-淀粉酶与作用底物之间的亲和作用,导致催化活性下降。

2.3 结合常数及热力学相关参数的分析
根据荧光猝灭数据,当荧光供体分子与猝灭剂受体分子发生结合时,荧光猝灭强度和猝灭剂浓度之间的关系符合修正后的Stern-Volmer方程[22]:

式中:F0、F分别为样品加入前后α-淀粉酶的相对荧光强度;Ka为结合常数;n为结合位点数;[Q]为样品质量浓度/(mg/mL)。
以lg[Q]为横坐标,lg[(F0-F)/F]为纵坐标作图,可得一组直线(图3)。由直线的截距和斜率可以求出不同CQA与酶相互作用的结合常数Ka和结合位点数n(表1)。

小分子与蛋白质之间可通过范德华力、氢键、静电引力以及疏水性作用力相互作用。根据CQA和α-淀粉酶结合过程中的反应焓变(ΔH)和熵变(ΔS)可确定两者之间的结合类型。根据van't Hoff方程,可计算出ΔH 和ΔS。


式中:K为相应温度条件下(293、310 K)荧光物质与猝灭剂之间的结合常数;R为气体常数(8.314 J/(mol·K));T为反应温度/K,当温度变化不大时,反应的焓变可以看作常量。
根据公式(3)、(4)计算得到ΔH和ΔS,再由式(5)可计算得到CQA与α-淀粉酶结合过程的吉布斯自由能(ΔG),结果列于表1。

据文献[23]报道,茶多酚与蛋白之间的结合常数通常为1.0×104~1.0×105L/mol。由表1可知,4 种CQA物质与α-淀粉酶的结合常数为1.65×106~2.18×107L/mol,表明两者之间存在较强的结合作用,双取代咖啡酰奎尼酸的结合常数略大于5-CQA,表明咖啡酰基的增多提高了结合能力,与抑制活性实验结果相一致。此外还可发现,在293 K和310 K温度条件下,结合常数随着温度的升高而增大,说明温度升高增加了CQA与α-淀粉酶之间的亲和性,提高了CQA-酶复合物的稳定性,从而说明在人体体温条件下更利于CQA抑酶活性的发挥。表1结果显示n值约为1,表明4 种CQA与α-淀粉酶之间均存在一个结合位点。
Ross等[24]根据大量实验总结了水相中小分子与生物大分子结合的热力学参数与主要作用力之间的关系:1)当ΔH>0、ΔS>0时,两者间为疏水作用力;2)当ΔH<0,ΔS<0时,则为氢键和范德华力;3)当ΔH≈0,ΔS>0,则为静电引力。根据表1结果可知,4 种CQA与α-淀粉酶结合反应的ΔH和ΔS均大于0,表明两者之间主要靠疏水作用力相互结合,同时,CQA与α-淀粉酶分子结构中均含有较多的羟基基团,因此氢键的作用仍然不可忽视。此外,结合过程中的ΔG小于0,表明CQA与α-淀粉酶之间的相互作用是自发进行的。
2.4 3D荧光光谱猝灭分析
3D荧光光谱是近年来发展起来的荧光分析技术,不仅具有灵敏度高和选择性强的特点,还能全面展现蛋白质的荧光特性[25]。α-淀粉酶具有内源荧光发色基团(主要为色氨酸、酪氨酸和脯氨酸),因此可采用3D荧光光谱有效表征CQA与α-淀粉酶相互作用后酶蛋白构象和空间分子结构的变化。

如图4所示,CQA在测定范围内无荧光发射,对结果不会造成干扰。α-淀粉酶在激发波长为200~300 nm,发射波长为300~400 nm范围内具有两个强荧光峰,标为峰1(230 nm/340 nm,λex/λem)和峰2(280 nm/340 nm,λex/λem),其中峰1是与酶蛋白多肽链的骨架结构相关,反映蛋白质二级结构变化,而峰2反映的主要是酶分子中色氨酸残基的荧光行为[26]。从图中可以看出,加入CQA后α-淀粉酶的两个峰明显被猝灭,其中3,4-diCQA和4,5-diCQA的猝灭效果最好,与常规荧光猝灭分析结果一致,再次证明双取代咖啡酰奎尼酸对α-淀粉酶有较强的结合作用。据报道,相对分子质量较大的多酚-蛋白质形成复合物的结合常数比相对分子质量较小的多酚-蛋白质复合物的结合常数要大,这可能是由于相对分子质量较大的多酚中存在更多羟基基团,与蛋白质分子之间形成更多的氢键,从而增加了复合物的稳定性[27]。CQA与α-淀粉酶相结合,由此导致酶蛋白空间构象以及所处微环境疏水性的变化,最终表现为对酶活性的抑制作用。
2.5 CQA对α-淀粉酶圆二色谱的影响
圆二色谱是一种特殊的吸收谱,它利用蛋白质等生物大分子具有的圆二色性得到关于生物大分子的二级结构信息。圆二色谱远紫外区段(178~250 nm)的主要生色团是肽链,这一波长范围的CD谱包含了生物大分子主链构象的信息[28-29]。为了进一步理解CQA与α-淀粉酶之间的结合机制,探讨其对酶蛋白二级结构产生的影响,本实验采用远紫外CD光谱分析5-CQA和3,5-diCQA结合后α-淀粉酶的构象变化。如图5所示,α-淀粉酶在208 nm 和222 nm波长左右有两个负峰,这是由肽链中的α-螺旋结构发生n-π*跃迁而产生的,即负Cotton效应[30]。加入样品(5-CQA或3,5-diCQA)后,α-淀粉酶的CD光谱发生了较大的变化。随着样品质量浓度的增加,α-淀粉酶在208 nm和222 nm波长处的两个负峰信号变弱,并且位置发生明显移动,表明酶蛋白二级结构中的α-螺旋含量降低。并且从图中可以看出,3,5-diCQA对α-淀粉酶二级结构的影响要大于5-CQA,这表明CQA结构中的咖啡酰基在与α-淀粉酶的结合过程中发挥了重要作用。


为了进一步定量分析CD图谱数据,本实验采用CDpro软件中的Selcon3算法对α-淀粉酶分子中α-螺旋、β-折叠、β-转角和无规卷曲结构的相对含量进行预测,结果列于表2。α-淀粉酶二级结构中包含17.6%的α-螺旋,19.2% 的β-折叠,8.2%的β-转角和55%的无规卷曲。5-CQA、3,5-diCQA与酶结合后使其4 种二级结构含量均发生变化,主要表现为α-螺旋含量降低,β-转角含量增多,说明酶蛋白的天然空间构象受到破坏,稳定性下降。

表2a-淀粉酶及a-淀粉酶与CQA结合后的二级结构含量Table2 Secondary structuralanalysisofa- amylaseandtheCQA-enzyme systems%样品 α-螺旋 β-折叠 β-转角 无规卷曲α-淀粉酶 17.6 19.2 8.2 55.0 α-淀粉酶+5-CQA(0.01 mg/mL) 16.7 43.1 9.2 31.0 α-淀粉酶+5-CQA(0.02 mg/mL) 11.9 29.9 21.0 37.2 α-淀粉酶+3,5-diCQA(0.01 mg/mL) 9.3 24.8 32.7 33.2 α-淀粉酶+3,5-diCQA(0.02 mg/mL) 3.4 15.8 32.7 48.1
3 结 论
本研究探讨了苦丁冬青苦丁茶中CQA类物质对α-淀粉酶的抑制作用,并采用荧光光谱法和圆二色谱法研究了两者之间相互作用的机理。结果表明CQA类物质能够抑制α-淀粉酶的活性,抑制能力由大到小为:3,5-diCQA>4,5-diCQA>3,4-diCQA>4-CQA>5-CQA>3-CQA。双取代咖啡酰奎尼酸的抑制效果强于单取代咖啡酰奎尼酸,说明分子中的咖啡酰基发挥了主要作用。CQA与α-淀粉酶的相互作用引起酶蛋白荧光光谱的猝灭,造成酶内部基团所处微环境的疏水性减弱。CQA与酶之间主要依靠疏水性作用力结合,并且只具有一个结合位点,所有反应均自发进行。CD光谱结果表明,CQA与α-淀粉酶形成稳定的复合物后使酶蛋白的二级结构发生变化,破坏了酶分子的空间构象,从而有效抑制了酶的催化活性。因此,本研究为探究苦丁茶中CQA类物质作为天然α-淀粉酶抑制剂的开发提供了理论基础,在糖尿病和肥胖症的预防和治疗中具有一定意义。
参考文献:
[1] HOLMAN R R, SOURIJ H, CALIFF R M.Cardiovascular outcome trials of glucose-lowering drugs or strategies in type 2 diabetes[J].The Lancet, 2014, 383: 2008-2017.DOI:10.1016/S0140-6736(14)60794-7.
[2] CERIELLO A.Postprandial hyperglycemia and diabetes complications[J].Diabetes, 2005, 54(1): 1-7.DOI:10.2337/ diabetes.54.1.1.
[3] CAI Xin, YU Jianan, XU Liman.The mechanism study in the interactions of sorghum procyanidins trimer with porcine pancreatic α-amylase[J].Food Chemistry, 2015, 174: 291-298.DOI:10.1016/ j.foodchem.2014.10.131.
[5] DALAR A, KONCZAK I.Phenolic contents, antioxidant capacities and inhibitory activities against key metabolic syndrome relevant enzymes of herbal teas from Eastern Anatolia[J].Industrial Crops and Products, 2013, 44: 383-390.DOI:10.1016/j.indcrop.2012.11.037.
[6] HE Qiang, LÜ Yuanping, YAO Kai.Effects of tea polyphenols on the activities of α-amylase, pepsin, trypsin and lipase[J].Food Chemistry,2007, 101(3): 1178-1182.DOI:10.1016/j.foodchem.2006.03.020.
[7] FEI Qunqin, GAO Yun, ZHANG Xin, et al.Effects of oolong tea polyphenols, EGCG, and EGCG3” Me on pancreatic α-amylase activity in vitro[J].Journal of Agricultural and Food Chemistry, 2014,62(39): 9507-9514.DOI:10.1021/jf5032907.
[8] MIAO Ming, JIANG Huan, JIANG Bo, et al.Phytonutrients for controlling starch digestion: evaluation of grape skin extract[J].Food Chemistry, 2014, 145: 205-211.DOI:10.1016/ j.foodchem.2013.08.056.
[9] 阮妙芸, 张根义.茶多酚对淀粉酶抑制作用的研究[J].安徽农业科学,2008, 36(11): 4371-4373.DOI:10.3969/j.issn.0517-6611.2008.11.004.
[10] THUONG P T, SU N D, NGOC T M, et al.Antioxidant activity and principles of Vietnam bitter tea Ilex kudingcha[J].Food Chemistry,2009, 113(1): 139-145.DOI:10.1016/j.foodchem.2008.07.041.
[11] SONG Chengwu, XIE Chao, ZHOU Zhiwen, et al.Antidiabetic effect of an active components group from Ilex kudingcha and its chemical composition[J].Evidence-Based Complementary and Alternative Medicine, 2012: 1-12.DOI:10.1155/2012/423690.
[12] 刘瑾, 丁平.冬青属药用植物资源、化学成分及药理作用研究进展[J].广州中医药大学学报, 2008, 25(3): 277-280.DOI:10.3969/ j.issn.1007-3213.2008.03.026.
[13] SUN Yi, XU Wenqing, ZHANG Wenqin, et al.Optimizing the extraction of phenolic antioxidants from kudingcha made frrom Ilex kudingcha C.J.Tseng by using response surface methodology[J].Separation and Purification Technology, 2011, 78(3): 311-320.DOI:10.1016/j.seppur.2011.01.038.
[14] 孙怡, 张鑫, 张文芹, 等.苦丁茶冬青苦丁茶中多酚类物质的分离纯化与结构解析[J].食品科学, 2011, 32(11): 60-63.
[15] 王晴川, 张鑫, 张文芹, 等.苦丁冬青苦丁茶中咖啡酰奎尼酸类物质的分离纯化和高效液相色谱法分析[J].食品科学, 2013, 34(22): 119-122.DOI:10.7506/spkx1002-6630-201322024.
[16] NARITA Y, INOUYE K.Inhibitory effects of chlorogenic acids from green coffee beans and cinnamate derivatives on the activity of porcine pancreas α-amylase isozyme I[J].Food Chemistry, 2011, 127(4): 1532-1539.DOI:10.1016/j.foodchem.2011.02.013.
[17] HANSAWASDI C, KAWABATA J, KASAI T.α-Amylase inhibitors from roselle (Hibiscus sabdariffa Linn.) tea[J].Bioscience,Biotechnology, and Biochemistry, 2000, 64(5): 1041-1043.DOI:10.1271/bbb.64.1041.
[18] SKRT M, BENEDIK E, PODLIPNIK Č, et al.Interactions of different polyphenols with bovine serum albumin using fluorescence quenching and molecular docking[J].Food Chemistry, 2012, 135(4): 2418-2424.DOI:10.1016/j.foodchem.2012.06.114.
[19] BUISSON G, DUEE E, HASER R, et al.Three dimensional structure of porcine pancreatic alpha-amylase at 2.9 A resolution.Role of calcium in structure and activity[J].EMBO Journal, 1987, 6(13): 3909-3916.
[20] RAWEL H M, FREY S K, MEIDTNER K, et al.Determining the binding affinities of phenolic compounds to proteins by quenching of the intrinsic tryptophan fluorescence[J].Molecular Nutrition & Food Research, 2006, 50(8): 705-713.DOI:10.1002/mnfr.200600013.
[21] WU Xuli, WANG Wenpu, ZHU T, et al.Phenylpropanoid glycoside inhibition of pepsin, trypsin and α-chymotrypsin enzyme activity in Kudingcha leaves from Ligustrum purpurascens[J].Food Research International, 2013, 54(2): 1376-1382.DOI:10.1016/ j.foodres.2013.10.020.
[22] LAKOWICZ J R.Principles of fluorescence spectroscopy[M].New York: Springer, 2006: 8-12.
[23] WU Xuli, DEY R, WU Hui, et al.Studies on the interaction of (-)-epigallocatechin-3-gallate from green tea with bovine β-lactoglobulin by spectroscopic methods and docking[J].International Journal of Dairy Technology, 2013, 66(1): 7-13.DOI:10.1111/j.1471-0307.2012.00873.x.
[24] ROSS P D, SUBRAMANIAN S.Thermodynamics of protein association reactions: forces contributing to stability[J].Biochemistry 1981, 20(11): 3096-3102.DOI:10.1021/bi00514a017.
[25] SANDHYA B, HEGDE A H, SEETHARAMAPPA J.Elucidation of binding mechanism and identification of binding site for an anti HIV drug, stavudine on human blood proteins[J].Molecular Biology Reports, 2013, 40(5): 3817-3827.DOI:10.1007/s11033-012-2460-8.
[26] SHEN Haibo, GU Zhengqin, JIAN Kang, et al.In vitro study on the binding of gemcitabine to bovine serum albumin[J].Journal of Pharmaceutical and Biomedical Analysis, 2013, 75: 86-93.DOI:10.1016/j.jpba.2012.11.021.
[27] KANAKIS C D, HASNI I, BOURASSA P, et al.Milk β-lactoglobulin complexes with tea polyphenols[J].Food Chemistry, 2011, 127(3): 1046-1055.DOI:10.1016/j.foodchem.2011.01.079.
[28] LI Daojin, WANG Ye, CHEN Jianjun, et al.Characterization of the interaction between farrerol and bovine serum albumin by fluorescence and circular dichroism[J].Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, 2011, 79(3): 680-686.DOI:10.1016/ j.saa.2011.04.005.
[29] BESLEY N A, HIRST J D.Theoretical studies toward quantitative protein circular dichroism calculations[J].Journal of the American Chemical Society, 1999, 121(41): 9636-9644.DOI:10.1021/ja990627l.
[30] TENG Yue, JI Fanying, LI Chao, et al.Interaction mechanism between 4-aminoantipyrine and the enzyme lysozyme[J].Journal of Luminescence, 2011, 131(12): 2661-2667.DOI:10.1016/ j.jlumin.2011.07.005.
DOI:10.7506/spkx1002-6630-201613002 10.7506/spkx1002-6630-201613002. http://www.spkx.net.cn [4] 康明丽.淀粉酶及其作用方式[J].食品工程, 2008(3): 11-13.10.3969/j.issn.1673-6044.2008.03.004.
中图分类号:TS272;TS201.3
文献标志码:A
文章编号:1002-6630(2016)13-0006-07
收稿日期:2016-01-14
基金项目:国家自然科学基金面上项目(31171666);江苏高校优势学科建设工程资助项目
作者简介:徐冬兰(1990—),女,硕士研究生,研究方向为食品生物技术。E-mail:2013108031@njau.edu.cn
*通信作者:孙怡(1966—),女,高级实验师,博士,研究方向为食品生物技术。E-mail:sunyi01@njau.edu.cn
Interaction Properties of Caffeoylquinic Acid Derivatives from Ilex kudingcha C.J.Tseng with α-Amylase
XU Donglan, WANG Qingchuan, ZENG Xiaoxiong, SUN Yi*
(College of Food Science and Technology, Nanjing Agricultural University, Nanjing 210095, China)
Abstract:The inhibitory effects of six caffeoylquinic acid (CQA) derivatives (3-CQA, 4-CQA, 5-CQA, 3,4-diCQA,3,5-diCQA and 4,5-diCQA) against α-amylase were studied comparatively by inhibitory activity assay.Furthermore, the potential interaction mechanisms between CQA derivative and α-amylase were investigated by fluorescence quenching and circular dichroism (CD) spectroscopy.The binding parameters were calculated according to modified Stern-Volmer equation,and the thermodynamic parameters were determined by the van't Hoff equation.The results showed that all CQA derivatives exhibited inhibitory effects on α-amylase and the half inhibitory concentrations (IC50) were 1.54, 1.05, 1.28, 0.96, 0.33 and 0.64 mg/mL, respectively.CQA derivatives interacted with α-amylase, forming stable complexes and leading to fluorescence quenching.Thermodynamic analysis indicated that the interaction process was spontaneous, and hydrophobic force might be primarily responsible for the interaction.In addition, the CD spectra suggested that the binding of CQA derivatives to the enzyme induced the change of protein structure, thus destabilizing the enzyme and reducing its activity.
Key words:α-amylase; caffeoylquinic acid; fluorescence spectroscopy; circular dichroism spectrum; interaction
猜你喜欢
食品与生物技术学报(2021年5期)2021-01-16
中国粮油学报(2019年4期)2019-07-12
卷宗(2016年10期)2017-01-21
分析化学(2016年7期)2016-12-08
考试周刊(2016年85期)2016-11-11
时代金融(2016年23期)2016-10-31
北方文学·中旬(2016年6期)2016-08-01
今日教育(2016年6期)2016-07-18
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
