
深绿木霉1-氨基环丙烷-1-羧酸脱氨酶基因的克隆及表达1)
2016-08-08 00:56刁桂萍李松阳王志英
东北林业大学学报 2016年8期
关键词:脱氨酶
刁桂萍 李松阳 王志英
(东北林业大学,哈尔滨,150040)
深绿木霉1-氨基环丙烷-1-羧酸脱氨酶基因的克隆及表达1)
刁桂萍李松阳王志英
(东北林业大学,哈尔滨,150040)
摘要以深绿木霉(Trichoderma atroviride)ACCC30153为试材,应用聚合酶链式反应(PCR)技术克隆获得1-氨基环丙烷-1-羧酸脱氨酶ACCD基因的cDNA全长。结果表明:此基因的cDNA编码区序列全长1 083 bp,编码区可编码360个氨基酸。经BlastP相似性分析,其属于Try-synth-beta-Ⅱ家族,与里氏木霉(T. reesei)氨基酸序列XP-006967764的同源性最高,达到91%。在5种不同诱导条件下,ACCD基因的表达量均有明显变化。在C、N饥饿培养基中分别培养8 h与16 h时,ACCD基因表达量达到最大值,为未诱导时的22.7倍与22.4倍;在山新杨叶粉与茎粉的诱导下,ACCD的表达量分别在培养的24 h与2 h达到最大,为未诱导的22.6倍与27.3倍。
关键词深绿木霉;脱氨酶;诱导表达
1-氨基环丙烷-1-羧酸脱氨酶(ACC脱氨酶)广泛存在于多种微生物中,它能催化乙烯前体分解来降低植物体内乙烯的合成,从而减少过量乙烯对植物自身生长造成的不良影响[1]。大量研究发现,土壤中微生物产生的ACC脱氨酶能促进植物根部的生长发育[2-4];而近年来研究也发现,ACC脱氨酶能减轻非生物因素(如干旱、重金属、盐碱等)对植物的危害[5-7]。此外,部分研究证明,含有ACC脱氨酶的细菌能减轻植物病害[8-9],但ACC脱氨酶与病原菌的作用机制还不清楚。
木霉菌(Trichodermaspp.)是一种极具应用价值的生防菌,不仅能抑制多种病原真菌,还能促进植物生长、诱导其免疫能力,更是具有降解土壤中盐类化合物、农药残留物及重金属离子等的功能[10]。然而,对于木霉菌中ACC脱氨酶生防功能的研究尚不多见。仅有的研究表明,深绿木霉(T. atroviride)通过产生ACC脱氨酶降解植物根部的乙烯前体,减少植物根部乙烯的合成来促进番茄幼苗的生长[11];棘孢木霉(T. asperellum)T203中的ACC脱氨酶也被证明在促进植物根部生长中起重要作用[12]。本研究通过聚合酶链式反应(PCR)技术获得了深绿木霉ACCC30153中ACC脱氨酶基因的cDNA全长,并研究了不同诱导条件下ACC脱氨酶的表达,为今后进一步研究深绿木霉ACC脱氨酶的生防功能提供参考。
1材料与方法
深绿木霉ACCC30153菌株,购于中国农业微生物菌种保藏管理中心。
根据转录组测序得到的ACCD基因部分cDNA序列设计引物,以深绿木霉ACCC30153菌株菌丝体总mRNA和基因组DNA为模板进行PCR扩增,克隆获得ACCD基因的全长cDNA和DNA序列。
使用ORF查找器寻找ACCD基因的开放读码框;用ExPASy-ProtParam软件分析蛋白质的等电点和相对分子质量等信息;通过BlastP预测蛋白保守区,寻找相似性序列;依据ClustalW软件进行多序列比对。
诱导培养基:MM基础培养基为15g/LNaH2PO4、453mg/LCaCl2、600mg/LMgSO4·7H2O、9.0mg/LFeSO4·7H2O、3.7mg/LCoCl2·7H2O、1.8mg/LMnSO4·H2O、2.5mg/LZnSO4·7H2O,并且含有5g/L(NH4)2SO4和质量分数为0.5%的葡萄糖;C饥饿诱导培养基为MM基础培养基(不含葡萄糖);N饥饿诱导培养基为MM基础培养基(不含硫酸铵);山新杨叶粉培养基为MM基础培养基(不含葡萄糖)+1%山新杨叶粉;山新杨茎粉培养基为MM基础培养基(不含葡萄糖)+1%山新杨茎粉。
菌丝体的培养与收集:将106个/mL木霉孢子接种于180mL1/4PD培养基中,于28 ℃、200r/min培养36h。然后,将菌丝体收集转接到上述5种诱导培养基中,分别于0、0.5、1.0、2.0、4.0、8.0、16.0、24.0h收集菌丝体,保存到冰箱中(-80 ℃)备用(重复3次)。
采用Trizol方法提取木霉菌丝体RNA,用DNA酶消化RNA中的DNA。参照TaKaRaPrimeScriptTMRT试剂盒(大连宝生物公司Code:DRR037S),以消化后的RNA为模板,反转录合成cDNA。将稀释10倍后的cDNA用作RT-qPCR模板。
RT-qPCR反应试剂盒为SYBRGreenRealtimePCRMastermix(ToyoboCo.,Ltd,Osaka,Japan)。反应体系为2×SYBRGreenRealtimePCRMastermix10μL、引物(10μmol/L)各0.5μL、去离子水6μL、cDNA模板3μL。RT-qPCR反应条件:95 ℃预变性30s;95 ℃ 5s、59 ℃ 15s、72 ℃ 10s;81 ℃读板1s,45个循环(引物序列见表1)。
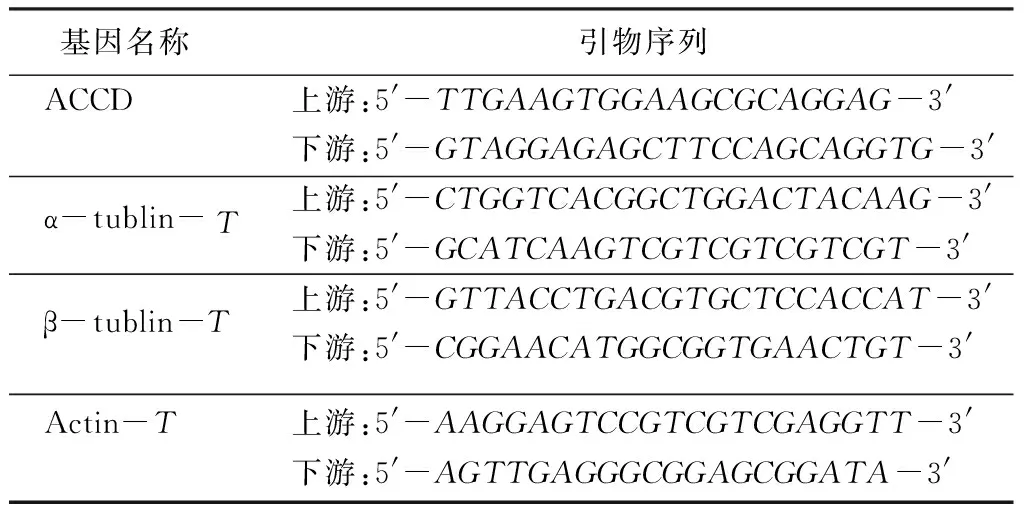
表1 RT-qPCR引物
2结果与分析
2.1深绿木霉ACCD基因的克隆与生物信息学分析
深绿木霉ACCD基因的cDNA全长为1 083bp,共编码360个氨基酸。ProtParam预测表明,该蛋白的相对分子质量为38140,理论等电点为6.13。通过BlastP进行保守区预测(见图1),该蛋白基因属于Try-synth-beta-Ⅱ家族,具有Try-synth-beta-Ⅱ家族的保守区结构域。

图1 ACCD保守区预测
通过NCBI的BlastP对ACCD基因进行同源性搜索,在GenBank的数据库中与深绿木霉ACCD基因氨基酸序列匹配的序列共有100个,相似性为65%~91%。随机选取14个ACCD基因进行多序列比对,结果表明:深绿木霉ACCD基因氨基酸序列与里氏木霉(XP-006967764)氨基酸序列的同源性最高为91%。含有2个高度保守的结构域,其中氨基酸序列的第295位和322位分别为Glu(E)和Leu(L),证明该蛋白质具ACC脱氨酶活性(见图2)。
2.2深绿木霉ACCD基因在不同诱导条件下的表达
采用实时荧光定量PCR方法检测了5种不同培养基诱导的深绿木霉ACCD基因在转录水平上的变化。RT-qPCR分析结果显示(见表2):在MM培养基培养条件下,深绿木霉ACCD基因在24h内呈连续上调表达,在16h时达到最大值,为未诱导时的23.5倍。在C、N饥饿培养条件下,该基因在培养的24h内均呈先上调后下降又大幅上调的表达趋势。在C饥饿培养中培养8h时,ACCD基因表达量达到最大值,为未诱导时的22.7倍;在N饥饿培养条件下,ACCD基因表达量在培养16h时达到最大值,为未诱导时的22.4倍。在山新杨叶粉培养基培养条件下,ACCD基因在24h内呈上调表达趋势,诱导仅1h表达量开始急剧上升,在24h时达到最大值,为未诱导时的22.8倍。经山新杨茎粉诱导后的ACCD基因表达量急剧上升,在第2h达到最大值,为未诱导时的27.3倍;此后,随培养时间延长,该基因表达量急剧下降。

XP_006967764为里氏木霉(Trichoderma reesei);XP_013951488为绿色木霉(Trichoderma virens);KKO97124为哈茨木霉(Trichoderma harzianum);ACX94231为棘孢木霉(Trichoderma asperellum);XP_013945914为深绿木霉(Trichoderma atroviride);KUE98859为盖姆斯木霉(Trichoderma gamsii);XP_007816500为绿疆菌(Metarhizium robertsii);KID86421为绿疆菌(Metarhizium guizhouense);XP_008597946为白僵菌(Beauveria bassiana);KPM40848为杓兰菌根菌(Neonectria ditissima);CCT63766为镰刀菌(Fusarium fujikuroi);KPA39012为镰刀菌(Fusarium langsethiae);KLP01246为镰刀菌(Fusarium fujikuroi);KIL92883为镰刀菌(Fusarium avenaceum)。

图2 ACCD氨基酸序列多序列比对
注:表中数据为“平均值±标准差”。
3结论与讨论
1-氨基环丙烷-1-羧酸脱氨酶(ACCD)是一种能抑制植物体内乙烯合成的胞内聚合酶,能切断植物激素乙烯合成前体,从而降低了环境与病原菌诱导对植物生长的抑制[3]。本研究克隆了深绿木霉ACCC30153中ACCD基因的全长cDNA序列,该基因的cDNA全长为1083bp,编码区可编码360个氨基酸。通过预测得知该蛋白属于Try-synth-beta-Ⅱ家族,且蛋白不含信号肽,是一种胞内酶。有研究表明,土壤中的天然细菌产生的ACC脱氨酶可将植物细胞中的乙烯前体分解,并利用其分解物做C源和N源,供自身利用[13]。本研究发现,在C、N饥饿条件下,深绿木霉中的ACCD基因表达呈上调趋势,说明该基因在木霉应对缺乏营养的条件过程中起到一定作用。而通过模拟深绿木霉与植物互作的研究发现,在与植物茎的互作过程中,ACCD基因会在互作的前期上调表达,表明当木霉受到山新杨茎中的ACC诱导后,导致其ACC脱氨酶转录水平提高,而随着培养时间的增强,植物茎所含ACC被分解,ACCD基因的表达也会随之下降[14]。ACCD最早是从土壤细菌中分离出来的。近年来发现,ACCD不仅存在于细菌中,在真菌中也检测到了它的活性。但是,具体作用还没有进行深入研究,本研究结果可为今后进一步研究深绿木霉ACCD基因的功能提供参考。
参考文献
[1]王海滨,王平,陈永华.ACC脱氨酶的作用机理和转基因的应用[J].生物技术通报,2009(4):40-43.
[2]黄盖,高焓,王琛,等.ACC脱氨酶活性菌株ACC30的分离、鉴定及其促生作用[J].微生物学通报,2013,40(5):812-821.
[3]GLICKBR,TODOROVICB,CZARNYJ,etal.PromotionofplantgrowthbybacterialACCdeaminase[J].CriticalReviewsinPlantScience,2007,26(5/6):227-242.
[4]GLICKBR,JACOBSONCB,SCHWARZEMMK,etal. 1-aminocyclopropane-1-carboxylicaciddeaminasemutantsoftheplant-growthpromotingrhizobacteriumPseudomonas putidaGR12-2donotstimulatecanolarootelongation[J].CanadianJournalofMicrobiology,1994,40(11):911-915.
[5]吉云秀,黄晓东.植物促生菌对燕麦初生苗盐分胁迫下的促生效应[J].大连海事大学学报,2007,33(3):86-89.
[6]魏素娜,蒋帅,黄锡云,等.旱地小麦根际细菌中产生1-氨基环丙烷-1-羧酸(ACC)脱氨酶菌株的分离和鉴定[J].微生物学通报,2011,38(5):722-728.
[7]KAYMAKHC,GUVENCI,YARALIF,etal.Theeffectsofbio-primingwithPGPRongerminationofradish(Raphanus sativusL.)seedsundersalineconditions[J].TurkJAgricFor,2009,33(2):173-179.
[8]INDIRAGANDHIP,ANANDHAMR,KIMK,etal.InductionofdefenseresponsesintomatoagainstPseudomnas syingaepv. tomatobyregulatingthestressethylenelevelwithMethylobacterium oryzaeCBMB20contating1-aminocyclopropane-1-carboxylicaciddeaminase[J].WorldJournalofMicrobiologyandBiochemistry,2008,24(7):1037-1045.
[9]龚凤娟,恩特马克·布拉提白,张宇凤,等.具有ACC脱氨酶活性的杜仲内生细菌的分离鉴定及其抗菌活性[J].微生物学通报,2011,38(10):1526-1532.
[10]遇文婧.深绿木霉刺激植物响应蛋白TatEpl1诱导杨树系统抗病性机制[D].哈尔滨:东北林业大学,2015.
[11]GRAVELV,ANTOUNH,TWEDDELRH.GrowthstimulationandfruityieldimprovementofgreenhousetomatoplantsbyinoculationwithPseudomonas putidaorTrichoderma atroviride:Possibleroleofindoleaaceticacid(IAA)[J].SoilBiologyandBiochemistry,2007,39(8):1968-1977.
[12]VITERBOA,LANDAUU,KIMS,etal.CharacterizationofACCdeaminasefromthebiocontrolandplantgrowth-promotingagentTrichoderma asperellumT203[J].FemsMicrobiologyLetters,2010,305(1):42-48.
[13]BELIMOVAA,HONTZEASN,SAFRONOVAVI,etal.Cadmium-tolerantplantgrowth-promotingbacteriaassociatedwiththerootofIndianmustard(Brassica junceaL.Czern.)[J].SoilBiologyandBiochemistry,2005,37(2):241-250.
[14]GLICKBR,CHENGZY,CZARNYJ,etal.PromotionofplantgrowthbyACCdeaminase-producingsoilbacteria[J].EuropeanJournalofPlantPathology,2007,119(3):329-339.
第一作者简介:刁桂萍,女,1981年11月生,东北林业大学林学院,副教授。E-mail:dgp2003@126.com。 通信作者:王志英,东北林业大学林学院,教授。E-mail:zyw0451@sohu.com。
收稿日期:2016年4月13日。
分类号Q933
CloningandDifferentialExpressionofAnACCDeaminaseGenefromTrichoderma atrovirideACCC30153//
DiaoGuiping,LiSongyang,WangZhiying
(NortheastForestryUniversity,Harbin150040,P.R.China)//JournalofNortheastForestryUniversity,2016,44(8):104-107.
ThecDNAofACCdeaminase(ACCD)genewasisolatedfromTrichoderma atrovirideACCC30153byusingPCR.ThecDNAsequenceofACCDwas1 083bpinlength,encoding360aminoacids.BlastPsearchindicatedthatACCDbelongedtoTry-synth-beta-Ⅱfamilyanditshownthehighestsimilarity91%withaproteinofT. reesei (XP-006967764). ACCDwasproventobedifferentiallytranscribedunderfivedifferenttreatments. ACCDtranscriptswereup-regulatedundernutritionalstressconditionswithpeaktranscriptionlevelsof22.7and22.4times,at8hand16h,respectively. ACCDtranscriptionwasalsoup-regulatedby1%leafandstempowderofPopulus davidiana×P. albavar. pyramidalis,withpeaktranscriptionlevelsof22.6and27.3times,at24hand2h,respectively.
KeywordsTrichoderma atroviride; ACC deaminase; Differential expression
1)中央高校基础科研业务费专项资金项目(2572014BA08);中国博士后科学基金面上项目(20110491014);黑龙江省博士后资助经费。
责任编辑:张玉。
猜你喜欢
南方农业(2022年6期)2022-04-13
商洛学院学报(2021年2期)2021-05-11
当代临床医刊(2021年1期)2021-04-01
医药前沿(2018年24期)2018-08-24
天津农业科学(2018年5期)2018-06-23
中国医药指南(2018年3期)2018-03-23
临床医药文献杂志(电子版)(2017年52期)2017-11-01
东方食疗与保健(2016年9期)2016-04-26
现代医药卫生(2015年4期)2015-12-26
中国医药指南(2015年29期)2015-10-27
