
晋西北黄土丘陵区柠条锦鸡儿根系分布特征
2016-08-08 00:56梁胜发茹豪雍鹏武秀娟奥小平
东北林业大学学报 2016年8期
梁胜发 茹豪 雍鹏 武秀娟 奥小平
(山西省林业种苗管理总站,太原,030012) (山西省林业科学研究院)
晋西北黄土丘陵区柠条锦鸡儿根系分布特征
梁胜发茹豪雍鹏武秀娟奥小平
(山西省林业种苗管理总站,太原,030012)(山西省林业科学研究院)
摘要以晋西北黄土高原10年生独生和簇生两种生长方式的柠条锦鸡儿(Caragana korshinskii Kom.)为研究对象,采用完全开挖采集根系的方法对不同土层深度内现存不同径级根系生物量、根长密度及比根长进行了研究。结果表明,独生根根系生物量、根长密度及比根长均高于簇生根,根系生态型是影响柠条锦鸡儿根系特征的根本原因。土壤资源的异质性是导致柠条锦鸡儿在土壤垂直方向上调整根系的形态和数量的主要原因。独生根根系较大的根长密度与比根长,具有更高的种内竞争力,提高了其适应环境的机制,比簇生根有更好的改良土壤、涵养水源与保持水土效应。
关键词黄土丘陵区;柠条锦鸡儿;根系生物量;根长密度;比根长
根系在地下与土壤颗粒盘旋缠绕结成根系网,将植物牢牢地固定在土壤中,起到保持水土的作用,一方面,根系将从土壤中吸收的水分与养分源源不断地输送至地上部分以维持整株植物的正常生长发育[1-3],另一方面,根系与土壤的相互作用既影响了根系在土壤中的分布特征,也促进了土壤理化性质的改变[4-5]。植物根系通过长期对土壤环境的适应与调整,以此来维持种群的生存和更新,这是构建稳定群落的基础[6-7]。
柠条锦鸡儿(Caragana korshinskiiKom.)属于豆科锦鸡儿属旱生落叶灌木,主要分布于我国北方干旱半干旱地区,是该地区最主要的灌木造林树种之一,具有良好的防风固沙、水土保持和土壤改良功能[9-10]。晋西北黄土丘陵区位于黄土高原东缘,是半湿润、半干旱区典型的风沙过渡区、农牧交错区和生态脆弱区[11-12],柠条锦鸡儿不仅是该区生态重建与恢复时选择的重要灌木树种,而且还是优质的灌木饲料,具有较高的生态和经济价值[13-14]。
目前,对柠条锦鸡儿人工林的研究多集中在单株植株根系动态及其垂直分布特征,但是经调查发现,在自然状态下,柠条锦鸡儿根系普遍存在独生根与簇生根两种生态型,这是自然选择的结果,与植物适应环境机制有关。本文通过对柠条锦鸡儿人工林不同土层内独生根及簇生根不同径级植物根系进行研究,分析柠条锦鸡儿人工林根系空间分布特征及独生根与簇生根根系之间的差异,以期为提出基于柠条锦鸡儿根系生态型的栽培技术提供理论支撑。
1研究区概况
研究区位于山西省西北部的五寨县(111°28′~113°E,38°44′~39°17′N),海拔1 300~2 736m,为半湿润地区向半干旱地区过渡地带。属温带寒冷半干旱气候,四季分明,春季干旱多风,夏秋雨量集中。年降水量400mm左右,年蒸发量1 913.2mm,年平均气温5.5 ℃,平均无霜期125d。采样地位于五寨县城边的李家口村,地势较为平坦,土壤为黄绵土,土壤肥力较低。土地利用受人类活动干扰比较大,自然植被破坏严重,主要土地利用类型有农田、灌木林地等。该地在2000年退耕还林工程开始大量种植柠条锦鸡儿,到目前已经形成了长达10年生的柠条锦鸡儿人工林。
2材料与方法
2.1材料采集及处理
2014年9月在10年生柠条锦鸡儿林地内设置面积为10m×20m的标准地,详细测定与记录灌木林中柠条锦鸡儿的高、地径、冠幅等指标,根据其平均值选择标准丛进行根系样品的测定。采用全根挖掘法挖出全部根系后,区分出独生根柠条锦鸡儿与簇生根柠条锦鸡儿,参考毕建琦等[15]对黄土丘陵区柠条锦鸡儿根系研究中的根系分级,将除根桩外的不同根系(d)分为5级,即d≤1mm,1mm
2.2根系生物量、根长密度及比根长测定
将分级后的各级别新鲜根系样品放在上面盖有玻璃的网格纸上,用镊子拉直两端测定根系长度(精确到0.5mm)。测定完毕后将其放入烘箱在65 ℃恒温下烘干至恒质量(48h),电子天平称量干质量。土壤样品体积内根系总长度为其根长密度,根系长度与其干质量的比值为比根长。计算4次取样中各参数的总和,并换算为单位体积生物量(kg·m-3)、根长密度(m·m-2)和比根长(m·g-1)。
3结果与分析
3.1根系生物量、根长密度及比根长的垂直分布
柠条锦鸡儿独生根与簇生根根系生物量、根长密度及比根长的垂直分布如表1所示。从表1中可以看出,独生根根系主要分布在0~40cm土层内,而簇生根根系主要分布在0~30cm土层内。0~30cm土层内,独生根根系生物量为26.74kg·m-3,而簇生根根系生物量为15.61kg·m-3,独生根根系生物量是簇生根根系生物量的1.71倍,并且每一层内的独生根根系生物量均高于簇生根;独生根与簇生根在>20~30cm土层内的根系分布最多,分别占0~30cm土层内根系的47.26%、40.15%。这表明独生根柠条锦鸡儿根系比簇生根柠条锦鸡儿在地下长势好。
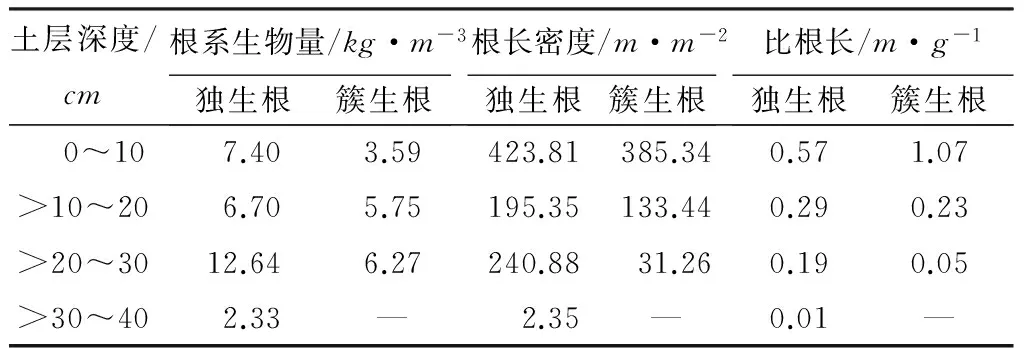
表1 两种生长方式根系生物量、根长密度及比根长垂直分布
根长密度表示单位土体内植物根系的长度之和,根长密度越大表明根系占据的土壤空间越大,更有利于植物根系吸收土壤中的水分和养分。由表1可见,在0~40cm土层内,独生根根系根长密度显著高于簇生根根系,在0~10cm土层内,独生根根长密度是簇生根的1.10倍,在>10~20cm土层内,独生根根系根长密度是簇生根根系的1.46倍,而在>20~30cm土层内,独生根根长密度是簇生根根系的7.71倍,在>30~40cm土层内没有发现簇生根这种生态型存在,随着土层加深,二者的根长密度相差越大。这说明独生根柠条锦鸡儿根系在土壤内占据的空间大于簇生根柠条锦鸡儿根系,其吸收水分和养分的能力也要高于簇生根柠条锦鸡儿根系。
比根长是指单位生物量植物根系的长度之和,比根长越大,表明植物根系的直径越细。根系直径越细,单位生物量根系的表面积越大,与土壤进行营养交换的面积就越大,一方面根系可以从土壤中吸收更多的水分与养分,另一方面,根系分泌物也可以较均匀地分泌至土壤中,柠条锦鸡儿作为豆科植物,比根长越大可以更好地起到改良土壤的作用。从表1中可以看出,在0~10cm土层内,簇生根根系比根长显著高于独生根根系,比根长是独生根根系的1.88倍,而在>10~20cm、>20~30cm土层内,独生根根系比根长分别是是簇生根的1.25倍、3.81倍。说明簇生根柠条锦鸡儿根系在0~10cm土层内的细根数量显著多于独生根柠条锦鸡儿根系,更容易吸收土壤中的水分和养分,说明簇生根柠条锦鸡儿根系对表层土壤的改良作用高于独生根。相反,在>10~40cm土层内,独生根柠条锦鸡儿根系的细根数量显著多于簇生根,说明独生根柠条锦鸡儿根系对下层土壤的改良作用高于簇生根。
从根系生物量、根长密度和比根长与根系生态型、土层深度之间的统计学关系可以看出,土层深度对各根系特征的影响不显著(p>0.05);根系生态型对各根系特征的影响是显著的(p<0.01)(表2)。由此可以认为,根系生态型是影响柠条锦鸡儿根系特征的主导因素。

表2 根系生态型和土层深度对根系特征影响的双因素方差分析
3.2不同径级根系生物量、根长密度及比根长垂直分布
3.2.1不同径级根系生物量垂直分布
独生根与簇生根不同径级根系生物量垂直分布如表3所示,0~40cm土层内独生根d≤5mm细根生物量为8.39kg·m-3,d>5mm粗根生物量为20.68kg·m-3;簇生根d≤5mm细根生物量为5.10kg·m-3,d>5mm粗根生物量为10.52kg·m-3。从表3中可以看出,0~10cm土层中,d≤1mm的独生根与簇生根生物量相差不大,独生根生物量仅比簇生根生物量少0.07kg·m-3,1mm
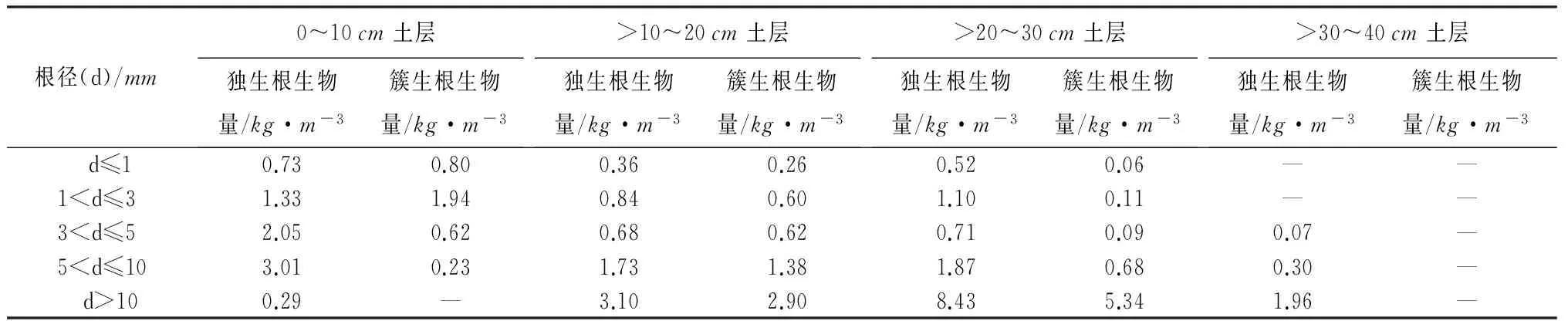
表3 不同土层深度下不同径级根系生物量垂直分布
3.2.2不同径级根长密度垂直分布
植物具有较高根长密度可以起到改良土壤、提高土壤入渗能力的作用[19]。根长密度一方面能够反映根系在土体中的延伸、穿插、交织程度,另一方面影响着养分向植物体的输送以及在土壤的运移[20],故根长密度越大,根系的穿插、交织程度越强,水分沿根孔入渗越快。此外,由于根长密度能反映根系在土体中的稠密度和交织穿插能力,且易于测量,常被用于研究根系分布与土壤渗透的关系[21]。柠条锦鸡儿独生根与簇生根不同径级根系根长密度垂直分布如表4所示,在0~10cm土壤表层内,根长密度随着根系径级的增大而减小,表明根径越细,根长密度越大。d≤1mm的细根根长密度最大,独生根达到341.16m·m-2,簇生根达到300.66m·m-2;1mm
以上分析表明,在0~30cm土层内,d≤3mm的根系是植物—土壤—水分系统中最关键的枢纽,其不仅起疏松土壤、固持土壤的作用,而且也是水分在植物、土壤之间进行频繁交互的重要通道。此外,独生根根系的根长密度大于簇生根,这与植物的种内竞争有关,簇生根根系之间争夺土壤中的水分与养分,互相制约,最终形成簇生根的根长密度小于独生根的格局。这种格局也反应出独生根根系与簇生根相比具有较好的水源涵养与水土保持效应。
3.2.3不同径级比根长垂直分布
比根长是根长和生物量的比值,是反映细根生理功能的重要指标之一,比根长较大的植物更容易从土壤中获取水分和养分,并且与根系生长、发育、生殖等生理功能密切相关[22-25]。柠条锦鸡儿独生根与簇生根不同径级根系比根长垂直分布如表5所示,比根长随着根系径级的增大而减小,这与根长密度的变化趋势相同。0~10cm土层内,d<1mm的细根比根长最大,独生根比根长为4.67m·g-1,簇生根比根长为3.74m·g-1,并且显著高于其他径级根系的比根长;与表层0~10cm土层内根系的比根长相比,10~20、20~30cm土层内d<1mm根系的比根长略小,随着土层深度的增加,比根长呈现逐渐减少的趋势,说明柠条锦鸡儿d<1mm根系在各土层内吸收水分与养分的能力逐层下降,这与上层土壤比下层土壤的水分、养分丰富有关。
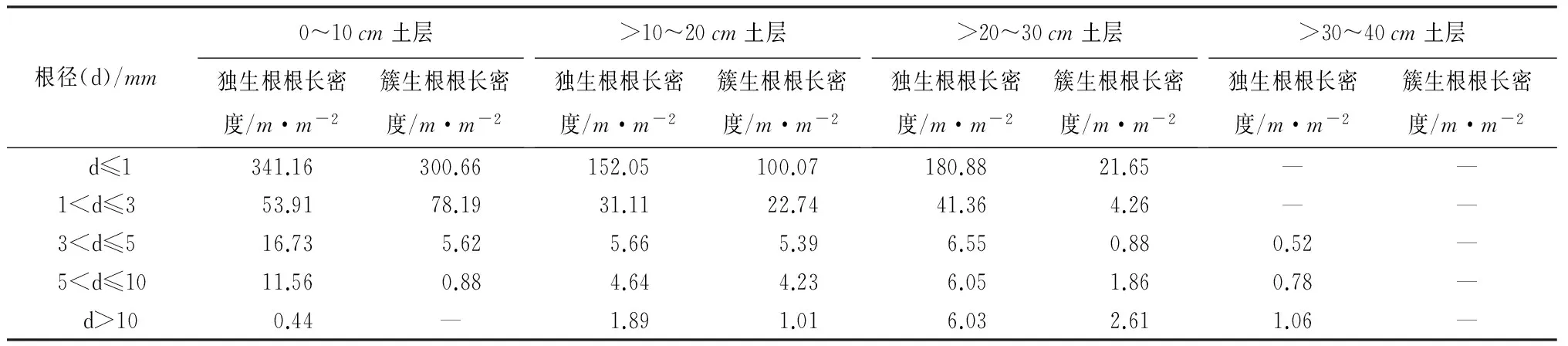
表4 不同土层深度下不同径级根系根长密度垂直分布
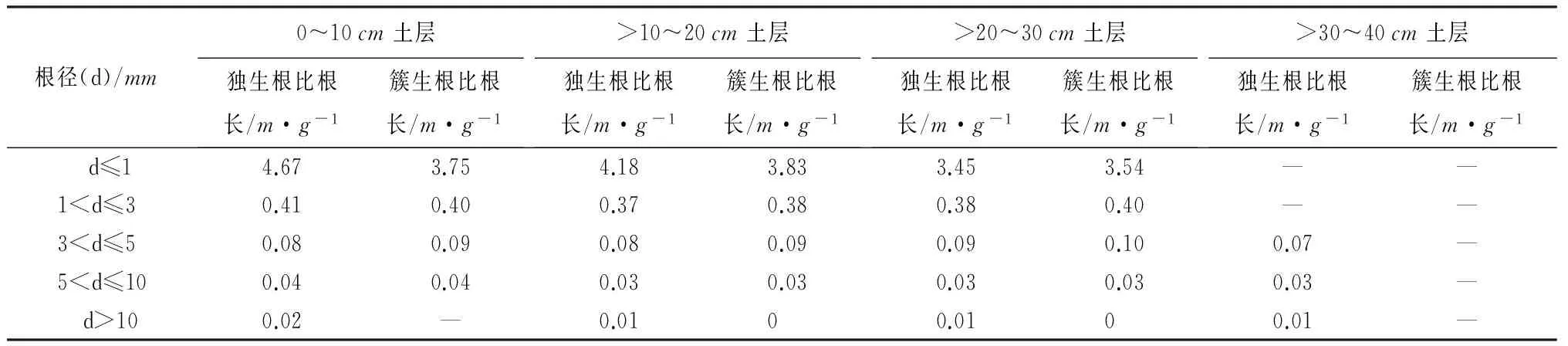
表5 不同土层深度下不同径级根系比根长垂直分布
在0~10、10~20cm土层内,柠条锦鸡儿独生根比根长较簇生根大,而在20~30、30~40cm土层内簇生根比根长较独生根略大,说明在0~20cm土层内独生根根系的表面积大于簇生根,与土壤接触的面积就大,更容易吸收土壤中的水分与养分,并且对土壤的改良作用也优于簇生根。相反,在20~40cm土层内簇生根更易吸收水分与养分,对土壤的改良作用优于独生根。
4结论与讨论
植物个体之间普遍存在竞争,主要竞争土壤中的水分以及矿物质等资源[26]。种内竞争通过改变植物密度、对生长和生殖间进行合理的营养分配,影响群落里植株的大小和生殖量,进而对群落大小和基因结构起作用[27]。研究表明,植物能够辨别自身的根系和其他植物的根系,为了避免自我竞争,自身的根系生长便会受到抑制以避免内部竞争,相反,当遇到其他植物的根系时,根系则会加快生长,争夺更多的土壤资源[28-30]。柠条锦鸡儿采用播种造林,在自然条件下形成独生与簇生两种生态型,在同一土层内,独生根比簇生根具有较大的生物量、根长密度、比根长;此外,独生根根系在土壤中的深度比簇生根深,利用垂直空间内土壤资源的能力比簇生根强。说明柠条锦鸡儿独生根在水平以及垂直方向竞争土壤中水分及矿物质资源的能力大于簇生根,更适宜在干旱、半干旱的环境中生长。此外,独生根具有较大的根长密度、比根长,根径小、呼吸快、氮富集量大,植株生长速度也较快,这些都与植物生长理论吻合[31-33]。
植物个体之间竞争土壤资源的能力一方面取决于土壤水分和养分的浓度、空间分布和供给速率,另一方面取决于植物根系表面密度、空间分布、与土壤接触面积以及对水分和养分的吸收速率[34]。因此,土壤资源的竞争主要发生在根系分布范围内[35]。在土壤垂直剖面上,土壤资源的异质性是植物通过调整根系的形态和数量获取土壤资源的根本原因[36]。柠条锦鸡儿独生根系的生物量、根长密度以及比根长在土壤垂直分布上均高于簇生根,这是根系缓解干旱压力和提高竞争能力的重要机制[37],因为干旱发生时深土层的水分较表层丰富,具有深根系的植物可以吸收更深土层的水分,更具存活及生长优势[38]。
晋西北地区位于我国北方农牧交错带上,是典型的生态脆弱区,该区生态环境严重恶化、自然资源显著匮乏,这是人与自然关系恶化的体现[11-12]。柠条锦鸡儿作为该区的主要造林树种,其生长状况直接影响当地的生态环境。本研究结果表明,独生根柠条锦鸡儿比簇生根柠条锦鸡儿更具有竞争力,这就需要在该地区造林时通过人为干预减少簇生根柠条锦鸡儿的数量,提高独生根柠条锦鸡儿群落的营林面积,在有限的土壤资源条件下,减少“冗余”植物对资源的消耗。
参考文献
[1]李建兴,何丙辉,谌芸,等.不同护坡草本植物的根系分布特征及其对土壤抗剪强度的影响[J].农业工程学报,2013,29(10):144-152.
[2]李强,刘国彬,许明祥,等.黄土丘陵区撂荒地土壤抗冲性及相关理化性质[J].农业工程学报,2013,29(10):153-159.
[3]杨吉华,李红云,李焕平,等.4种灌木林地根系分布特征及其固持土壤效应的研究[J].水土保持学报,2007,21(3):48-51.
[4]李建兴,何丙辉,谌芸,等.不同护坡草本植物的根系特征及对土壤渗透性的影响[J].生态学报,2013,33(5):1535-1544.
[5]李青山,王冬梅,信忠保,等.漓江水陆交错带典型灌木群落根系分布与土壤养分的关系[J].生态学报,2015,35(15):5104-5109.
[6]张宇清,朱清科,齐实,等.梯田埂坎立地植物根系分布特征及其对土壤水分的影响[J].生态学报,2005,25(3):500-506.
[7]杜明新,周向睿,周志宇,等.毛乌素沙南缘紫穗槐根系垂直分布特征[J].草业学报,2014,23(2):125-132.
[8]贾丽,曲式曾.豆科锦鸡儿研究进展[J].植物研究,2001,21(4):515-518.
[9]王孟本,陈建文,史建伟,等.柠条锦鸡儿人工幼林细根生长和死亡的季节变化[J].生态学报,2010,30(19):5121-5130.
[10]陈建文,王孟本,史建伟.柠条锦鸡儿人工林幼林与成林细根动态比较研究[J].生态学报,2011,31(22):6978-6988.
[11]秦作栋,董光荣,马志正.晋西北地区土地荒漠化现状分析[J].中国沙漠,1995,15(3):244-251.
[12]李秀芬,刘利民,齐鑫,等.晋西北生态脆弱区土地利用动态变化及驱动力[J].生态学报,2014,25(10):2959-2967.
[13]牛西午.中国锦鸡儿属植物资源研究:分布及分种描述[J].西北植物学报,1999,19(5):107-133.
[14]牛西午,张强,杨治平,等.柠条锦鸡儿人工林对晋西北土壤理化性质变化的影响研究[J].西北植物学报,2003,23(4):628-632.
[15]毕建琦,杜峰,梁宗锁,等.黄土高原丘陵区不同立地条件下柠条锦鸡儿根系研究[J].林业科学研究,2006,19(2):225-230.
[16]JACKSONRB,CANADELLJ,EHLERINGERJR,etal.Aglobleanalysisofrootdistributionforterrestrialbiomes[J].Oecologia,1996,108(3):389-411.
[17]PREGITZERKS,HENDRICKRL,FOGELR.Thedemographyoffinerootsinresponsetopatchesofwaterandnitrogen[J].NewPhytologist,1991,260(2):516-521.
[18]BURTONAJ,PREGITZERKS,HENDRICKRL.RelationshipsbetweenfinerootdynamicsandnitrogenavailabilityinMichigannorthernhardwoodforests[J].Oecologia,2000,125(3):389-399.
[19]王国梁,刘国彬.黄土丘陵区长芒草群落对土壤水分入渗的影响[J].水土保持学报,2009,23(3):227-231.
[20]韩凤鹏,郑纪勇,张兴昌.黄土退耕坡地植物根系分布特征及其对土壤养分的影响[J].农业工程学报,2009,25(2):50-55.
[21]徐少君,曾波,类淑桐,等.三峡库区几种耐水淹植物根系特征与土壤抗水蚀增强效应[J].土壤学报,2011,18(1):160-167.
[22]徐琨,李芳兰,苟水燕,等.岷江干旱河谷25种植物一年生植株根系功能性状及相互关系[J].生态学报,2012,32(1):215-225.
[23]EISSENSTATDM.Costsandbenefitsofconstructingrootsofsmalldiameter[J].JournalofPlantNutrition,1992,15(6/7):763-782.
[24]ESSSENSTATDM,CALDWELLMM.Competitiveabilityislinkedtoratesofwaterextraction[J].Oecologia,1988,75(1):1-7.
[25]EISSENSTATDM.Ontherelationshipbetweenspecificrootlengthandtherateofrootproliferation:afieldstudyusingcitrusrootstocks[J].NewPhytologist,1991,118(1):63-68.
[26]DRIEVERSM,NESEHV,ROIJACKERSRMM.GrowthlimitationofLemnaminorduetohighplantdensity[J].AquaticBotany,2005,81(3):245-251.
[27]HAGIHARA.TheoreticalconsiderationsontheC-Deffectinself-thinningplantpopulations[J].ResearchesonPopulationEcology,1999,41(2):151-159.
[28]FALIKO,REIDESP,GERSANIM,etal.Self/non-selfdiscriminationinroots[J].JournalofEcology,2003,91(4):525-531.
[29]GRUNTMANM,NOVOPLANSKYA.Physiologicallymediatedself/non-selfdiscriminationinroots[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2004,101(11):3863-3867.
[30]MAHALLBE,CALLAWAYRM.EffectsofregionaloriginandgenotypeonintraspecificrootcommunicationinthedesertshrubAmbrosiadumosa(Asteraceae)[J].AmericanJournalofBotany,1996,83(1):93-98.
[31]XUEL,HAGIHARAA.DensityeffectsoftreeorgansinselfthinningPinus densifloraSieb.etZuccstands[J].EcologicalResearch,2008,23(4):689-695.
[32]陈伟,薛立.根系间的相互作用——竞争与互利[J].生态学报,2004,24(6):1243-1251.
[33]薛立,傅静丹.影响植物竞争的因子[J].中南林业科技大学学报,2012,32(2):6-15.
[34]BIONDINIM.Athree-dimensionalspatialmodelforplantcompetitioninanheterogeneoussoilenvironment[J].EcologicalModelling,2001,142(3):189-225.
[35]TESSIERJT,MCNAUGHTONSJ,RAYNALDJ.InfluenceofnutrientavailabilityandtreewildlingdensityonnutrientuptakebyOxalisacetosellaandAcersaccharum[J].EnvironmentalExperimentalBotany,2001,45(1):11-20.
[36]HODGEA.Theplasticplant:rootresponsestoheterogeneoussuppliesofnutrients[J].NewPhytologist,2004,162(1):9-24.
[37]KINGJ,GAYA,SYLVESTER-BRADLEYR,etal.Modellingcerealrootsystemsforwaterandnitrogencapture:towardsaneconomicoption[J].AnnalsofBotany,2003,91(3):383-390.
[38]HOMD,ROSASJC,BROWNKM.etal.Rootarchitecturaltradeoffsforwaterandphosphorusacquisition[J].FunctionalPlantBiology,2005,32(8):737-748.
第一作者简介:梁胜发,男,1962年4月生,山西省林业种苗管理总站,高级工程师。E-mail:Liangshengfa369@163.com。 通信作者:茹豪,山西省林业科学研究院,工程师。E-mail:ruhao1987@163.com。
收稿日期:2016年1月5日。
分类号S727.23
RootDistributionCharacteristicsofCaragana korshinskiiintheLoessHillyAreaofNorthwesternShanxiProvince//
LiangShengfa
(ShanxiForestrySeedandSeedlingAdministrationCentralStation,Taiyuan030012,P.R.China);RuHao,YongPeng,WuXiujuan,AoXiaoping(ShanxiAcademyofForestrySciences)//JournalofNortheastForestryUniversity,2016,44(8):24-28.
WestudiedthedifferenceofCaragana korshinskiiKom.rootsystemdistributioncharacteristicsbetweenthesinglerootandtheclusterrootina10-aoldplantationintheLoessPlateauareaofNorthwestShanxi,China.Bytakingroottotalbiomass(RTB),rootlengthdensity(RLD),andspecificrootlength(SRL)asbasicparameters,weanalyzedthecharacteristicsofrootsbywholeplantexcavationsbasedonselectionofrepresentativeplantsamples.TheRTB,RLDandSRLofsinglerootsweremorethanthatofclusterroots.RootecotypewasthekeycauseofaffectingtherootscharacteristicsofC. korshinskii.HeterogeneityofsoilresourceswasthemainreasonofleadingC. korshinskiitoadjustthemorphologyandthenumberintheverticaldirectiontocapturesoilresources.TherewerelargerRLDandSRLofthesingleroot,anditsintraspecificcompetitionandenvironmentadaptationmechanismwerehigherthanthoseoftheclusterroot.Thesingleroothasbettereffectofimprovingthequalityofthesoilandwaterresourceconservation.
KeywordsThe loess hilly area; Caragana korshinskii Kom.; Root biomass; Root length density; Specific root length
责任编辑:潘华。
