
竹林环境中木质素降解菌株的分离鉴定
2016-08-07 07:46:29李新鑫余学军蒋玉俭
竹子学报 2016年2期
李新鑫, 余学军,俞 暾,蒋玉俭
(浙江农林大学亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
竹林环境中木质素降解菌株的分离鉴定
李新鑫, 余学军,俞 暾,蒋玉俭
(浙江农林大学亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
为筛选竹林环境中较为高效的木质素降解菌,采用分析苯胺蓝和愈创木酚培养基变色情况和相关酶活大小,从浙江临安市竹林地中分离筛选出一株高效木质素降解菌27-1,根据形态学观察和ITS序列分析,确定该菌株为多孔菌科栓菌属毛栓菌(Trameteshirsuta)。
木质素降解菌;变色;漆酶;筛选
纤维素是自然界中储量最丰富的有机物[1],是一种潜在的十分重要的生物能源。木质素的去除是纤维素进一步高效利用的关键[2-3]。竹子作为一种木本禾本科植物,生长更新速度很快,每年能够产生大量的纤维素类有机物。木质素的存在使得纤维素的的利用和降解程度大大降低。如何降解木质素成为了提高纤维素利用率的关键。竹林中残存过多的覆盖物和枯落物也不利于竹林的良好更新,必须在次年对覆盖物进行及时的清除,而木质素的去除是进一步加快纤维类有机物降解的关键,也是造纸行业中制浆的关键。在过去,纤维类有机物在利用前均会进行不同程度、不同方式的预处理,如碾压,蒸汽爆破[4],酸碱处理[5],超流体萃取[6],有机溶剂处理等[7-9]。但这些方法处理成本高,且容易造成环境污染,因此如何快速、有效、安全、方便的去除木质素一直是纤维类有机物质利用或降解的重点,生物降解是降解竹材木质素和林地覆盖物的一种安全可靠的方法。
国内对木质素降解微生物的研究起步于20世纪90年代中期,并且分别从造纸废液、药渣、堆肥和森林土壤中分离筛选出不同的木质素降解菌。晋果果等[10]人从张家界土壤中分离筛选出一株只降解木质素而对纤维素没有降解能力的高温木质素降解菌地芽孢杆菌GeobacilluscaldoxylosilyticusJ16,李红亚等[11]人从牛粪中分离出一株解淀粉芽孢杆菌,该细菌对玉米秸秆中的木质素有很好的降解作用。习兴梅[12]从堆肥中分离到一株黑曲霉,不仅对木质素有一定降解能力,对半纤维素和纤维素也有很好的降解能力。木质素的微生物降解主要通过木质素降解酶系完成[13],其中漆酶、木质素过氧化氢酶、锰过氧化氢酶3种酶起着主要的作用[14,15]。本研究通过测定分离菌株的变色能力大小和3种酶活大小来筛选降解木质素的优良竹林微生物。
1 材料和方法
1.1 材料
1.1.1 样品来源 本实验55个样品为采自浙江临安及其周边地区竹林地的朽竹和腐殖质丰富的土壤,采集时间为2014年8月到9月和2015年4月到8月。
1.1.2 培养基 液体PDA培养基:马铃薯200 g,葡萄糖20 g,KH2PO43 g,MgSO4·7H2O 1.5 g,VB10.01 g,蛋白胨10 g,蒸馏水1 L。
固体PDA培养基:在液体培养基的基础上加入20 g·L-1的琼脂。
定性筛选培养基:分别在固体培养基中加入0.1 g·L-1的苯胺蓝和0.04%愈创木酚。
产酶培养基:参照刘小刚,吕聪等[16-17]的产酶培养基配比,将葡萄糖浓度改为10 g·L-1,氮源改为酒石酸铵0.2g·L-1, NH4NO30.1g·L-1和麸皮8g·L-1的混合氮源,培养基pH为4.5。
以上培养基均在121 ℃,灭菌15分钟。
1.1.3 实验器材 高压蒸汽灭菌锅,电子分析天平,紫外-可见光分光光度计,离心机,电磁炉,超净工作台,恒温培养箱,恒温摇床,电泳仪,PCR仪,连接仪,超低温冰箱。
1.2 实验方法
1.2.1 菌株分离与筛选 样品破碎后,称取适量样品(土壤样品5 g,植物样品2 g)加入到装有100 mL PDA液体培养基的250 mL三角瓶中,在恒温培养箱中培养1 d(30 ℃,160 r·min-1),取1 mL培养液梯度稀释,取20 μl稀释倍数在10-3,10-4,10-5的样品涂布于平板培养基上,每个样品及梯度做3次重复并在30 ℃的恒温培养箱中培养3-5 d,挑取单菌落在平板培养基中划线培养,培养3-5 d,反复划线以获得单一菌株。将获得的纯菌株划线接种于苯胺蓝PDA平板培养基和愈创木酚PDA平板培养基中培养观察。对引起苯胺蓝PDA培养基褪色或愈创木酚PDA培养基出现红褐色变化的菌株进行连续接种培养,观察其在培养期间的变色能力有无明显差异,将在培养过程中均能引起筛选培养基有效变色的菌种斜面接种,保存于4 ℃冰箱备用。
将筛选的菌株活化后点状接种于苯胺蓝PDA平板培养基和愈创木酚PDA平板培养基中,观察菌落生长和变色情况,并测量菌落及变色圈直径。对比各菌株对苯胺蓝平板的褪色效果和愈创木酚平板的变色能力。
1.2.2 菌株酶活复筛 将变色能力强的菌株接种于PDA培养基中,30 ℃培养数天,以获得足够的菌株。用打孔器取下直径7 mm的菌饼三片接种于装有100 mL产酶培养基的250 mL三角瓶中,在恒温震荡培养箱中震荡培养,培养条件30 ℃,200转每分钟,从第7天起每两天取样测定各菌株发酵液的酶活活力,至第15天。
1.2.3 酶活测定方法
(1)粗酶液提取。取适量发酵液,在4 ℃环境下8 000 r·min-1离心10 min,移取上清液备用。以沸水灭活10 min的酶液作为对照。
(2)漆酶(lac)酶活测定方法。漆酶酶活测定采用以2,2-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐(ABTS)为反应底物的方法测定,反应环境为25 ℃,反应体系3 mL,反应底物为2 mL的溶解于0.1 mmol·L-1的醋酸-醋酸钠(pH=5.0)缓冲液中的浓度为0.5 mmol·L-1的ABTS,加入1 mL的酶液启动反应,测定最初3分钟内420 nm处的吸光值变化。定义每分钟氧化1 μmol的ABTS为一个酶活单位(ε=3.6×104[(mol/L)-1·cm-1])[18]。
(3)锰过氧化氢酶(MnP)酶活的测定方法。反应体系4 mL,50 mmol·L-1(pH=4.5)的乳酸-乳酸钠缓冲溶液3.4 mL,1.6 mmol·L-1的MnSO4水溶液0.1 mL,酶液0.4 mL,混匀后37 ℃水域保温,反应时添加1.6 mmol·L-1的H2O20.1 mL启动反应,测定最初3分钟内240 nm处的吸光值变化。定义每分钟使Mn2+转化为Mn3+所需的酶量为一个酶活单位(ε=6.5×103[(mol/L)-1·cm-1])[19]。
(4)木质素过氧化氢酶(Lip)酶活测定方法。反应体系3 mL,0.24 mol·L-1(pH=3)的酒石酸-就是酸钠缓冲液1.85 mL,24 mmol·L-1的藜芦醇0.1 mL,酶液1 mL,混匀后37 ℃水域保温,反应时添加6 mmol·L-1的H2O20.05 mL启动反应,测定最初3分钟内310 nm处的吸光值变化。定义每分钟氧化1 μmol的藜芦醇为一个酶活单位(ε=9.3×106[(mol/L)-1·cm-1])[20]。
1.2.4 菌种鉴定
(1)菌种的形态观察。采用插片发制作菌体玻片,采用革兰氏染色法对其进行染色,将玻片置于显微镜下观察。参考《真菌鉴定手册》[21]和《中国真菌志》[22]对其进行初步的分类。
(2)菌种的ITS序列分析。采用CTAB法提取菌株DNA,引物采用通用引物ITS1(TCCGATGGTGA ACCTGCGG)和ITS4(TCCTCCGCTTATTGATATGC)并以提取的DNA为模板进行PCR扩增,PCR扩增条件如下:95 ℃预变性3 min,94 ℃变形30 s,55 ℃退火30 s,72 ℃延伸45 s,35个循环,72 ℃保持10 min,12 ℃保温。对PCR产物进行回收,连接到PMD-19载体上,导入DH5α型感受态细胞,并将转化后的感受态细胞涂布于含有AMP的LB平板培养基上,培养12-15小时,挑取单菌落进行菌落PCR。将菌落PCR结果为阳性的菌液送交生工生物工程有限公司进行序列测定,对所得序列进行人工剪切并在NCBI上进行BLAST。选取同源性较高的菌种,采用Mega.5.0软件NJ法进行聚类分析,构建系统进化树。
2 结果与分析
2.1 菌株初筛与复筛
从富含腐殖质的竹林土壤和朽竹中初步选出85份可以引起培养基变色的菌株,经过连续接种培养,筛选出一些能稳定的使定性培养基变色的菌株。通过测量变色圈直径、菌落直径测定初步判断各菌株的产酶活力大小。各菌株在定性筛选培养基上的变色能力大小如表1所示:菌株6-1,27-1,39-1的Hb较大,菌株1-1,48-2,48-5等有较大的Ha比值。通过酶活测定,从中选出了8株酶活较为明显的的菌株。其中以菌株6-1和27-1有最大的Hb值,但在培养10天后,两者的液体发酵产酶活力相差较大,菌株27-1的漆酶酶活可以达到94.389 U,而6-1只有2.111U左右(部分菌株酶活见表2),究其原因可能是6-1的适宜培养条件与当前发酵条件相差较大或该菌株分泌的是其他类型的多酚氧化酶。48-5有最大的Hb值。虽然27-1的Hb值很小,但培养菌株十天后,其MnP活性最大,达到了34.87 U·mL-1,其他菌株均有一定的Mnp活性,但酶活不高,一般在3-7 U左右。而Lip酶活活性均未检测到。综合27-1有着较高的Lac活性和Mnp活性,故确定最佳木质素降解菌株为27-1。
2.2 菌株27-1的产酶周期测定
对选出的27-1进行发酵周期测定,结果如图1,由图中可知,菌株27-1在第9天的时候表现出一定的产酶活性。其中Lac活性在第15天达到74.86 U,比菌株复筛时酶活性大小略有差异且产酶略晚,可能是不同传代次数的菌株间的产酶能力大小差异引起;Mnp活性仅在第11天时达到一般正常产酶水平,为3.248 U,在第23天时达到最大为16.068 U。与酶活筛选时酶活性差异较大,且产酶较晚。说明菌株27-1的Mnp活性并不是十分稳定;整个周期中依然没有表现出Lip活性。

表1 部分菌株的定性筛选结果

表2 8株菌的三种酶活力大小
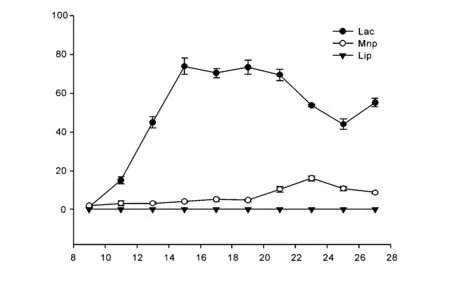
图1 菌株27-1发酵过程中三种酶活的活性大小Fig.1 The activities of three enzymes during the fermentation of fungus 27-1
2.3 菌株27-1鉴定
菌体形态特征见图 。菌株27-1在PDA培养基上生长时,菌丝白色,绒毛状,有横隔与分枝,有锁状联合,菌落圆形边缘不整齐,初步判断菌株27-1为担子菌纲的真菌。采用真菌通用引物序列ITS1和ITS4对所提取的菌株DNA进行PCR扩增后,得到一组长度大约在600 bp左右的片段,将其扩增序列通过连接转化,然后交由生工生物工程有限公司进行测序,并将其序列与GenBank中已有DNA序列进行同源性比对,发现同源性在99%以上的ITS序列均为栓菌属的毛栓菌。故判断菌株27-1为多孔菌科栓菌属毛栓菌Trameteshirsuta,是一种白腐真菌。

图2 菌株27-1及来自Genbank的相似物种的ITS序列构建的NJ系统发育树Fig.2 NJ phylogenetic tree of fungus 27-1 and similar speices from Genbank
3 结论讨论
本研究通过培养基变色初筛、酶活测定复筛的方法,从竹林地中分离出1株高效木质素降解菌,通过生物学初步观察和ITS序列分子鉴定,确定该菌株为担子菌纲多孔菌科栓菌属毛栓菌。测定了该菌株的Lac酶活和MnP酶活。目前,对木质素降解菌的分离筛选已经有了很多的报道,参与降解木质素的微生物包括细菌、放线菌和真菌,其中真菌是最为重要的一类,且关于竹林中相关类型的微生物的研究报道不多,这些筛选到的微生物广泛的应用到,生物制浆、造纸、废水及污染处理、废弃物再利用、生物预处理等方面。本研究筛选到的毛栓菌27-1分泌的木质素降解酶系中Lac酶活性较高,在30 ℃,pH 4.5葡萄糖为碳源,麸皮、硝酸铵、酒石酸铵为混合氮源的条件下,Lac酶活达到74.86 U,高于曾祥康[23]从枯朽木中筛选到的毛栓菌SYBC-LZ(漆酶酶活为60.50 U),由于真菌漆酶是一种多酚氧化酶[24],能够催化多种酚类[25]、羧酸、以及甲氧基酚酸的脱甲基反应[26]等,可以用于废水处理、土壤修复、重金属处理和木质素降解等多个方面[25-27]。尤其是近年来雷竹林地由于长期覆盖经营引起的多酚氧化酶活性降低,酚酸类物质和重金属积累等问题[28-29]。筛选到的毛栓菌可以配合一些纤维素降解菌共同施放到覆盖竹林中,共同促进木质素和纤维素的降解,降低林地的C/N比;消除林地重金属;降低酚酸含量,修复林地土壤。
[1] Lynd L R, Wyman C E, Gerngross T U. Biocommodity engineering[J]. Biotechnology progress, 1999, 15(5): 777-793.
[2] 吴薇,姜训鹏,吕程序,等.高效木质素降解菌的分离筛选[J].食品科技,2008,3:22-25.
[3] 赵红霞, 杨建军, 詹勇. 白腐真菌在秸秆作物资源开发中的研究[J]. 饲料工业, 2002, 23(11): 40-42.
[4] Beltrame P L, Carniti P, Visciglio A, et al. Fractionation and bioconversion of steam-exploded wheat straw[J]. Bioresource technology, 1992, 39(2): 165-171.
[5] Han Y W, Callihan C D. Cellulose fermentation: effect of substrate pretreatment on microbial growth[J]. Applied Microbiology, 1974, 27(1): 159-165.
[6] 刘江燕,武书彬.稻草和毛竹CO_2超临界流体萃取脱木质素研究燕[J].北京林业大学学报,2011,33(2):96-104.
[7] Wyman C E, Dale B E, Elander R T, et al. Coordinated development of leading biomass pretreatment technologies[J]. Bioresource technology, 2005, 96(18): 1959-1966.
[8] Demirba A. Utilization of lignin degradation products from hazelnut shell via supercritical fluid extraction[J]. Energy sources, 2002, 24(9): 891-897.
[9] 张学俊, 张陶芸. 超临界流体萃取原理用于制浆研究[J]. 中国造纸学报, 1995: 77-85.
[10] 晋果果,翁海波,李萍萍,等.高温木质素降解菌Geobacil_省略_J16的筛选及其产酶发酵性质研究[J].中国农学通报,2011,27(8):334-339.
[11] 李红亚,李术娜,王树香,等.产芽孢木质素降解菌MN_8的筛选及其对木质素的降解[J].中国农业科学,2014,47(2):324-333.
[12] 习兴梅, 曾光明, 郁红艳, 等. 黑曲霉 Aspergillus niger 木质纤维素降解能力及产酶研究[J]. 农业环境科学学报, 2007, 26(4): 1506-1511.
[13] Keyser P, Kirk T K, Zeikus J G. Ligninolytic enzyme system of Phanaerochaete chrysosporium: synthesized in the absence of lignin in response to nitrogen starvation[J]. Journal of bacteriology, 1978, 135(3): 790-797.
[14] 张辉.木质素降解酶系研究新进展[J].天津农业科学,2006,12(3):8-12.
[15] 侯红漫,周集体,陈丽.白腐菌漆酶特性及异生芳香化合物的降解[J].林产化学与工业,2003,23(1):89-94.
[16] 刘小刚.葡萄枝条木质素降解菌的筛选[D].杨凌:西北农林科技大学,2009:1-51.
[17] 吕聪. 茯苓菌产锰过氧化物酶及其应用的研究[D]. 中南林业科技大学, 2008.
[18] Arora D S, Gill P K. Effects of various media and supplements on laccase production by some white rot fungi[J]. Bioresource Technology, 2001, 77(1): 89-91.
[19] Zunong L Y G P W. NUTRITIONAL REGULATION OF SYNTHESIS OF LIGNIN PEROXIDASE BY PHANEROCHATE CHRYSOSPORIUM ME-446 [J]. Acta Microbiologica Sinica, 1994, 1: 005.
[20] 李娜. 木质素降解菌的筛选及其产酶性质的研究 [D]. 西北农林科技大学, 2008.
[21] 张纪忠. 真菌鉴定手册[J]. 1990.
[22] 中国真菌志: Flora fungorun sinicorun. Vol. 3, Polyporaceae. 多孔菌科. 第三卷[M]. 科学出版社, 1998.
[23] 曾祥康. 毛栓菌发酵产漆酶及其在合成染料脱色中的应用[D]. 江南大学, 2012.
[24] 吴晴. 白腐真菌及其漆酶的应用研究[J]. 安徽科技, 2010 (3): 47-48.
[25] 王华, 金大勇, 赵建夫. 真菌漆酶及其在环境污染控制中的应用[J]. 环境科学与技术, 2003, 26(6): 58-60.
[26] 王国栋, 陈晓亚. 漆酶的性质, 功能, 催化机理和应用[J]. 植物学通报, 2003, 20(4): 469-475.
[27] 周雪婷, 张跃华, 罗志文, 等. 真菌漆酶的研究进展及其应用前景[J]. 农业与技术, 2013, 32(9): 2-4.
[28] 刘国群, 庄舜尧, 孙绣华. 不同种植年限雷竹林土壤微生物, 量和酶活性变化[J]. 安徽农业科学, 2012, 40(2): 866-868.
[29] 郑仁红. 覆盖栽培对雷竹林衰退的化感效应研究[D]. 北京: 中国林业科学研究院, 2006.
Separation and Identification of Lignin-degrading Microorganisms from Bamboo Stand Soils
LI Xin-xin, YU Xue-jun, YU Tun, JIANG Yu-jian
(The Nurturing Station for the State Key Laboratory of Subtropical Silviculture,Zhejiang A&F University, Lin'an 311300, Zhejiang, China)
Cellulose is the most extensive and abundant renewable resource in nature, being a potentially important biological energy. As the cellulose and hemicelluloses are coated by the lignin, the removal of lignin is the key to efficiently use cellulose. At present, few studies on the separation and identification of lignin-degrading microorganisms from bamboo stand soils have been described. Based on the analysis of discoloration and related enzyme activity of aniline blue and guaiacol medium, an efficient lignin-degrading fungus 27-1, was obtained from the bamboo stand soils in Lin'an and the surrounding area, and identified asTrameteshirsuteby morphological observation and sequence analysis.
Lignin degrading microorganisms; Discoloration; Laccase; Separation and Identification
2015-11-10
浙江省重大推广工程项目(2012T201)
李新鑫(1989-),男,硕士研究生,从事竹林培育与利用研究。E-mail:muyelxx@163.com。通信作者:余学军,男,副研究员,从事竹子栽培与利用研究。E-mail:yuxj@zafu.edu.cn
猜你喜欢
上海包装(2019年8期)2019-11-11 12:16:14
小学生必读(低年级版)(2019年12期)2019-04-20 08:39:22
童话世界(2018年23期)2018-08-23 09:13:16
小学阅读指南·低年级版(2017年7期)2017-08-04 04:23:38
天津造纸(2016年1期)2017-01-15 14:03:29
山东工业技术(2016年15期)2016-12-01 05:31:50
中国酿造(2016年12期)2016-03-01 03:08:19
中国造纸学报(2015年1期)2015-12-16 19:35:25
应用海洋学学报(2015年3期)2015-11-22 07:39:30
中国塑料(2014年4期)2014-10-17 03:00:45
