
发酵条件对苹果渣发酵饲料中4种水解酶活性的影响
2016-08-04 06:47任雅萍来航线陈姣姣薛泉宏
西北农林科技大学学报(自然科学版) 2016年7期
任雅萍,郭 俏,来航线,陈姣姣,薛泉宏
(西北农林科技大学 a 生命科学学院,b 资源环境学院,陕西 杨凌 712100)
发酵条件对苹果渣发酵饲料中4种水解酶活性的影响
任雅萍a,郭俏b,来航线b,陈姣姣b,薛泉宏b
(西北农林科技大学 a 生命科学学院,b 资源环境学院,陕西 杨凌 712100)
[摘要]【目的】 研究发酵菌种、原料组成及发酵原料灭菌处理对苹果渣发酵饲料中4种重要水解酶(蛋白酶、纤维素酶、果胶酶及植酸酶)活性的影响。【方法】 以酵母菌(Yeast)、黑曲霉A8(Aspergillus niger A8)和米曲霉(Aspergillus oryzae)为发酵菌剂,通过单一菌种及双菌混合接种方法,分别对2种原料(原料A为苹果渣+尿素,原料B为苹果渣+尿素+油渣粉)在自然发酵(原料不灭菌)和灭菌发酵(原料121℃灭菌30 min)条件下进行固态发酵,利用比色法测定发酵产物中蛋白酶、纤维素酶、果胶酶及植酸酶4种水解酶的活性。【结果】 发酵可大幅度提高发酵产物中影响饲料消化利用率的4种重要水解酶活性,且混合发酵剂对发酵产物中4种水解酶活性的提高作用优于单菌发酵,添加油渣粉和原料灭菌处理有助于提高发酵产物中4 种水解酶的活性。灭菌和添加油渣粉工艺条件下,在发酵产物中:蛋白酶活性58.85~368.43 U,发酵增率67.7%~949.6%,接菌增率109.4%~526.0%,灭菌增率22.2%~118.6%;纤维素酶活性3 617.53~6 278.22 U,发酵增率13.8%~97.6%,接菌增率2.3%~73.5%,灭菌增率8.3%~63.9%;果胶酶活性203.20~358.47 U,发酵增率34.5%~137.4%,接菌增率26.4%~76.4%,灭菌增率为17.1%~64.8%;植酸酶活性19.45~54.96 U,发酵增率336.1%~1 132.3%,接菌增率41.4%~182.6%,灭菌增率1.2%~92.0%。【结论】 混菌发酵、添加油渣粉以及发酵原料灭菌处理对提高苹果渣发酵饲料中蛋白酶、纤维素酶、果胶酶及植酸酶活性均有显著作用。
[关键词]苹果渣;固态发酵;蛋白酶活性;纤维素酶活性;果胶酶活性;植酸酶活性
我国是世界上苹果产量最多的国家之一,年产量在2 000万 t 以上,每年由果汁加工而产生的苹果渣达120万 t。苹果渣含水量约80%,其干物质中蛋白质含量仅占3%~5%,直接用作饲料营养价值较低,提高苹果渣的营养品质已成为其作为饲料资源开发利用的关键。采用特定工艺通过微生物发酵提高苹果渣发酵产物的蛋白质含量及其营养价值,是苹果渣合理利用的重要途径之一。在苹果渣发酵饲料中,除蛋白质以外,发酵产生的蛋白酶、纤维素酶等水解酶类对动物营养也具有重要作用。已有研究表明,发酵饲料中存在的水解酶类可大幅度提高营养物质的消化利用率,促进家畜生长,增强动物免疫力[1]。因此,在关注苹果渣发酵饲料蛋白质品质及其发酵工艺优化的同时,进一步研究不同发酵工艺处理对果渣发酵饲料水解酶活性的影响,对苹果渣发酵饲料的品质评价也具有重要的意义。
目前关于利用苹果渣发酵生产蛋白饲料的发酵工艺已有较多研究[2-4],但对苹果渣单细胞蛋白发酵饲料中的水解酶研究不多。张高波等[5]探索了酵母菌与霉菌不同接种菌剂组合对苹果渣发酵饲料中蛋白酶和纤维素酶活性的影响,但对苹果渣固态发酵饲料中的果胶酶及植酸酶活性尚无报道,且关于发酵原料中辅料添加及原料灭菌处理对苹果渣发酵饲料中水解酶活性的影响尚无系统研究。
本研究以苹果渣为原料,以酵母菌和2株霉菌(黑曲霉A8和米曲霉)为发酵剂,系统研究了发酵剂、原料组成及原料灭菌方式等3种因素不同组合处理对苹果渣发酵饲料中蛋白酶、纤维素酶、果胶酶及植酸酶活性的影响,旨在为苹果渣发酵饲料品质评价及生产工艺优化提供科学依据。
1材料与方法
1.1材料
菌种:酵母菌(Yeast)、黑曲霉A8(AspergillusnigerA8)及米曲霉(Aspergillusoryzae),均由西北农林科技大学资源环境学院微生物资源研究室提供。
原料:干苹果渣(自然条件下风干),由乾县海升果汁厂提供。油渣粉(油菜籽榨油后所余残渣粉碎而成),市购。
发酵干原料:原料A为干苹果渣和尿素按质量比19∶1混合;原料B为干苹果渣、油渣粉和尿素按质量比17∶2∶1混合。
发酵湿原料:由发酵干原料与自来水按质量比1∶2混合。
培养基:马铃薯葡萄糖琼脂(PDA)培养基,其中含100 g马铃薯,10 g葡萄糖,7.5 g琼脂,500 mL水,121 ℃灭菌20 min。
1.2方法
1.2.1发酵剂制备向250 mL三角瓶中装入50 mL PDA灭菌培养基,待冷凝后将活化好的酵母菌悬液1 mL接入该瓶中,用金属刮铲涂匀后于28 ℃下培养3 d;向瓶中加100 mL无菌水制得菌悬液。经血球计数板测定,其活菌数为4.0×109个/mL。按相同方法制备黑曲霉A8及米曲霉孢子悬液,于28 ℃下培养5 d,所得孢子数分别为8.7×108和1.1×109CFU/mL。
1.2.2固态发酵设计设发酵剂、灭菌方式及原料组成3个因素,发酵剂设不接菌(CK1)、单接酵母菌(Y)、单接黑曲霉A8(H)、单接米曲霉(M)、酵母菌+黑曲霉A8(YH)、酵母菌+米曲霉(YM)6个处理;灭菌方式设自然发酵(原料不灭菌)和灭菌发酵(原料121 ℃灭菌30 min)2个处理;原料组成设原料A(苹果渣+尿素)和原料B(苹果渣+尿素+油渣粉)2种类型。试验共24个处理,每处理重复3次,以未发酵纯苹果渣原料作为对照(CK)。
1.2.3试验步骤按照1.1节中的比例配制发酵原料,然后分别称取50.0 g发酵湿原料于250 mL组培瓶中,121 ℃下灭菌30 min,冷却至室温,接入发酵剂(单菌接种量为5 mL,混菌各2.5 mL),用灭菌竹签搅匀,28 ℃下培养72 h,发酵结束后将样品在45 ℃下鼓风烘干并粉碎备用。自然发酵中原料不灭菌直接接种。
1.3发酵产物中水解酶活性的测定
1.3.1蛋白酶称烘干样品1.000 g于三角瓶中,加pH 7.2磷酸缓冲液20 mL,40 ℃水浴保温60 min,过滤后选择适宜稀释倍数采用福林法[6]测定蛋白酶活性。将1.0 g发酵产物在40 ℃下每1 min水解干酪素产生1 μg酪氨酸的酶活定义为1个蛋白酶活性单位,用1 U表示。
1.3.2纤维素酶称烘干样品0.500 g于三角瓶中,加蒸馏水20 mL,40 ℃水浴保温45 min,过滤后选择合适稀释倍数以羧甲基纤维素纳(CMC)为底物,用DNS(3,5-二硝基水杨酸)法[7]测定还原糖含量,计算纤维素酶活性。将在pH 5.0、40 ℃下每 1.0 g发酵产物在1 min内水解CMC生成1 μg葡萄糖的酶活性定义为1个纤维素酶活性(CMC酶活)单位,用1 U表示。
1.3.3果胶酶称烘干样品0.500 g于三角瓶中,加pH 4.8乙酸-乙酸钠缓冲液20 mL,48 ℃水浴45 min,过滤后选择合适稀释倍数用DNS(3,5-二硝基水杨酸)法[8]测定果胶酶的活性。将1.0 g发酵产物在48 ℃、pH 4.8的条件下1 h内水解果胶底物转化成1 mg D-半乳糖醛酸的果胶酶活性定义为1个果胶酶活性单位,用1 U表示。
1.3.4植酸酶称烘干样品0.500 g于三角瓶中,加pH 5.5乙酸-乙酸钠缓冲液20 mL,37 ℃水浴30 min,过滤后选择合适的稀释倍数用偏钒酸铵法[9]测定植酸酶活性。将1.0 g发酵产物在植酸钠浓度为5 mmol/L、温度37 ℃、pH 5.5条件下反应1 min,从植酸钠中释放1 μmol无机磷的植酸酶活性定义为1个植酸酶活性单位,用1 U表示。
1.4数据处理
以未发酵原料为对照,由发酵引起蛋白酶活性的增率称为发酵增率(Δf);以未接菌处理为对照,由接菌引起蛋白酶活性的增率称为接菌增率(Δi);以原料未灭菌处理为对照,由原料灭菌引起蛋白酶活性的增率称为灭菌增率(Δs);蛋白酶活性的发酵增率(Δf)、接菌增率(Δi)和灭菌增率(Δs)均采用下式计算:
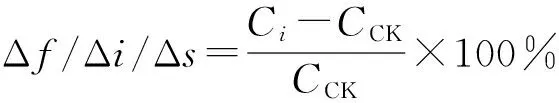
式中:CCK分别表示未发酵、未接菌及未灭菌处理的蛋白酶活性(U);Ci分别为发酵、接菌和灭菌处理的蛋白酶活性(U)。纤维素酶、果胶酶及植酸酶酶活性增率的计算与蛋白酶相同。
2结果与分析
2.1不同处理对苹果渣发酵产物中蛋白酶活性的影响
2.1.1自然发酵不同菌剂及辅料对未灭菌原料发酵产物中蛋白酶活性的影响结果见表1。
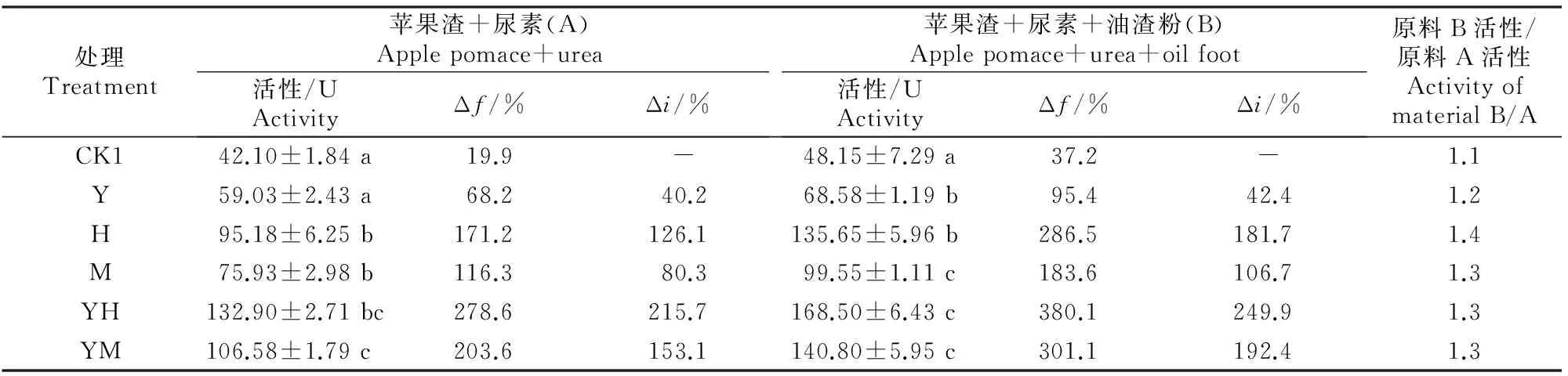
表 1 不同菌剂及辅料对未灭菌原料发酵产物中蛋白酶活性的影响Table 1 Effect of different microbial inoculums and auxiliary materials on protease activity in fermentation product of unsterilized material
注:未发酵纯苹果渣的蛋白酶活性为35.10 U;同列数据后标不同小写字母者表示差异显著(P<0.05);下表同。
Note:The protease activity in blank sample was 35.10 U.Different lowercase letters mean significant difference (P<0.05).The same below.
由表1可以看出,在自然发酵条件下,原料A不接种发酵剂,单靠原料及空气中微生物自然接种,发酵产物的蛋白酶活性由未发酵纯苹果渣的35.10 U提高到42.10 U,发酵增率(Δf)为19.9%,而接种酵母菌+黑曲霉A8、酵母菌+米曲霉、黑曲霉A8、米曲霉及酵母菌后,发酵产物中蛋白酶活性较未发酵纯果渣原料显著提高,其发酵增率(Δf)分别为278.6%,203.6%,171.2%,116.3%及68.2%;与不接发酵剂的原料A相比,接种发酵剂处理的蛋白酶活性接种增率(Δi)为40.2%~215.7%。原料B与之类似,特别是在接种酵母菌+黑曲霉A8时,蛋白酶活性的发酵增率(Δf)及接菌增率(Δi)分别为380.1%及249.9%,即原料中加入油渣粉能大幅度提高酵母菌+黑曲霉A8发酵产物中的蛋白酶活性。
2.1.2灭菌发酵由表2可知,在灭菌条件下,发酵能大幅提高发酵产物的蛋白酶活性。在原料A中,发酵产物的蛋白酶活性为55.70~246.05 U,发酵增率(Δf)为58.7%~601.0%。在不同发酵剂处理中,蛋白酶活性的接菌增率(Δi)为57.5%~341.7%,其中接种酵母菌+黑曲霉A8时蛋白酶活性较不接菌(CK1)增加341.7%。在原料B中,发酵产物中蛋白酶活性高达58.85~368.43 U,蛋白酶活性的发酵增率(Δf)为67.7%~949.6%,接菌增率(Δi)为109.4%~526.0%,且以酵母菌+黑曲霉A8接种处理的接菌增率最高(526.0%)。
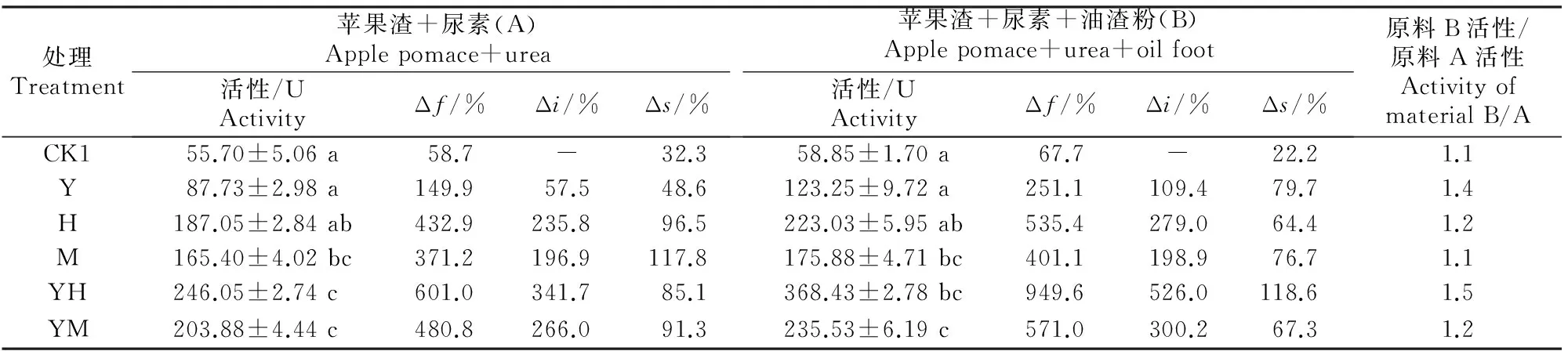
表 2 不同菌剂及辅料对灭菌原料发酵产物中蛋白酶活性的影响Table 2 Effect of different microbial inoculums and auxiliary materials on protease activity in fermentation product of sterilized material
注:未发酵纯苹果渣的蛋白酶活性为35.10 U。
Note:The protease activity in blank sample was 35.10 U.
2.1.3原料灭菌及添加油渣粉对发酵产物中蛋白酶活性的影响比较表1和表2并结合表2中的灭菌增率(Δs)可以看出,灭菌条件下,发酵产物的蛋白酶活性均明显大于未灭菌的自然发酵。在原料A中,自然发酵与灭菌发酵产物的蛋白酶活性分别为42.10~132.90 U与55.70~246.05 U,其灭菌增率(Δs)为32.3%~117.8%,灭菌处理发酵产物蛋白酶活性增幅明显,说明原料灭菌能显著提高发酵产物的蛋白酶活性,且原料B与原料A类似。
由表1和表2可以看出,无论原料是否灭菌,同一发酵处理中,原料B发酵产物的蛋白酶活性及其发酵增率(Δf)均明显高于原料A的。上述分析表明,向原料中添加油渣粉有助于提高发酵产物中的蛋白酶活性及发酵增率。
2.2不同处理对苹果渣发酵产物中纤维素酶活性的影响
2.2.1自然发酵不同菌剂及辅料对未灭菌原料发酵产物中纤维素酶活性的影响见表3。
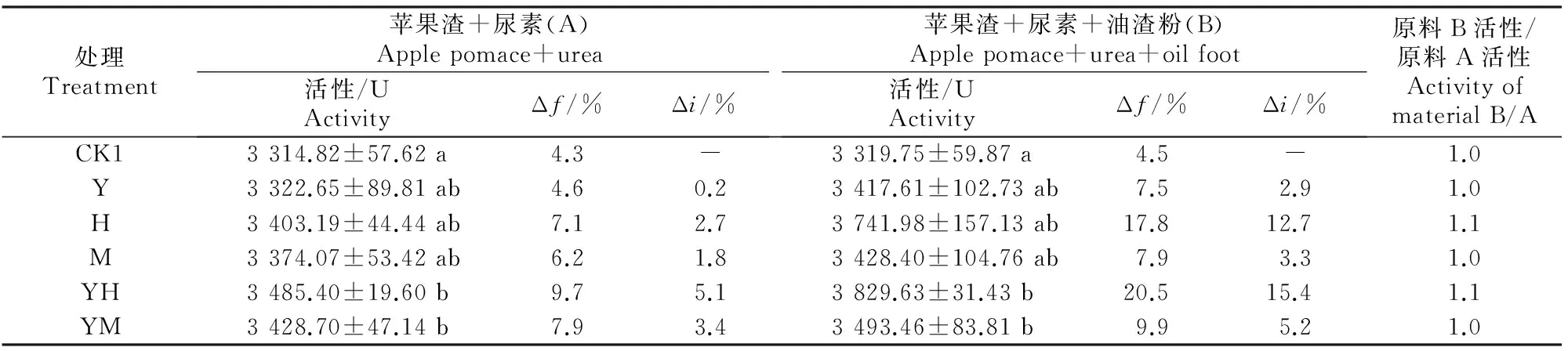
表 3 不同菌剂及辅料对未灭菌原料的发酵产物中纤维素酶活性的影响Table 3 Effect of different microbial inoculums and auxiliary materials on cellulase activity in fermentation product of unsterilized material
注:未发酵纯苹果渣的纤维素酶活性为3 177.78 U。
Note:The cellulase activity in blank sample was 3 177.78 U.
由表3可以看出,原料A在不灭菌和不接种发酵剂的自然条件下,靠原料及空气中微生物自然接种,可以使发酵产物的纤维素酶活性由未发酵纯苹果渣的3 177.78 U增加到 3 314.82 U,发酵增率(Δf)为4.3%;接种酵母菌+黑曲霉A8、酵母菌+米曲霉、黑曲霉A8、米曲霉及酵母菌后,发酵产物中纤维素酶活性的发酵增率(Δf)分别为9.7%,7.9%,7.1%,6.2%及4.6%;与不接种对照处理相比,纤维素酶活性的接菌增率(Δi)为0.2%~5.1%。原料B与之类似,特别是在接种酵母菌+黑曲霉A8时,纤维素酶活性的发酵增率(Δf)及接菌增率(Δi)分别为20.5%和15.4%。
2.2.2灭菌发酵由表4看出,在灭菌发酵中,接菌处理能大幅提高发酵产物的纤维素酶活性。在原料A中,发酵产物的纤维素酶活性为3 465.43~5 695.56 U,黑曲霉A8+酵母菌、黑曲霉A8、米曲霉+酵母菌及米曲霉处理与不接菌对照的纤维素酶活性差异显著;纤维素酶活性的发酵增率(Δf)为 9.1%~79.2%,其中不接菌处理纤维素酶活性的发酵增率(Δf)为9.1%;在不同发酵剂处理中,纤维素酶活性的接菌增率(Δi)为1.1%~64.4%,其中接种酵母菌+黑曲霉A8混菌时纤维素酶活性较对照增加64.4%,增幅明显。在原料B中,发酵产物中纤维素酶活性高达3 617.53~6 278.22 U,发酵增率(Δf)为13.8%~97.6%,纤维素酶活性的接菌增率(Δi)为2.3%~73.5%。
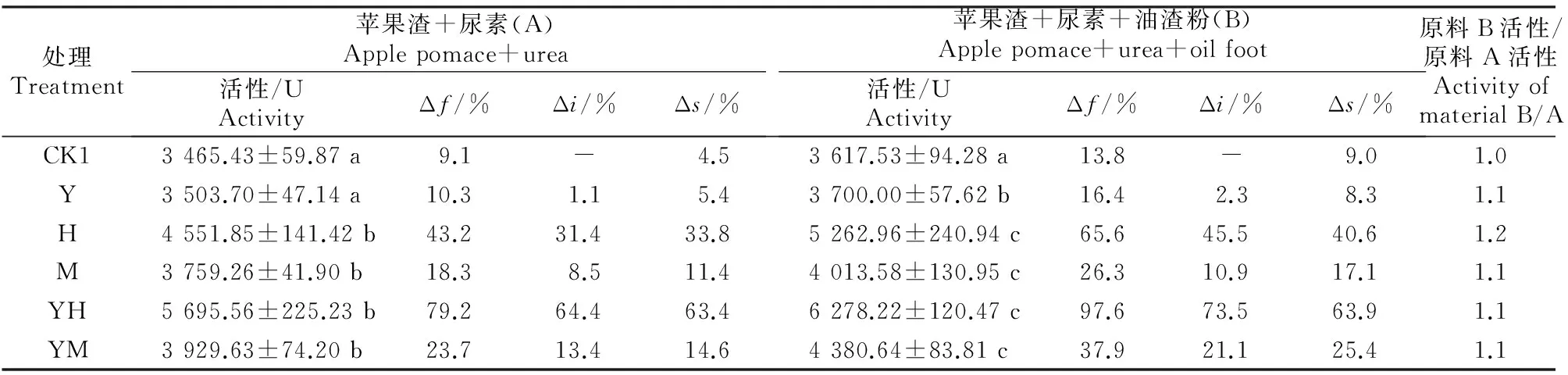
表 4 不同菌剂及辅料对灭菌原料发酵产物中纤维素酶活性的影响Table 4 Effect of different microbial inoculums and auxiliary materials on cellulase activity in fermentation product of sterilized material fermentation products of sterilized material
注:未发酵纯苹果渣的纤维素酶活性为3 177.78 U。
Note:The cellulase activity in blank sample was 3 177.78 U.
2.2.3原料灭菌及添加油渣粉对发酵产物中纤维素酶活性的影响由表4灭菌增率(Δs)可知,灭菌条件下发酵产物的纤维素酶活性均明显大于自然发酵,原料A、原料B的灭菌增率(Δs)分别为 4.5%~63.4%和8.3%~63.9%,说明灭菌能提高发酵产物的纤维素酶活性。结合表3、表4可以看出,在未灭菌及灭菌条件下,原料B发酵产物纤维素酶活性均明显高于原料A,原料B的发酵增率也明显高于原料A。由此可见,原料中添加油渣粉对提高发酵产物纤维素酶活性及发酵增率有一定促进作用。
2.3不同处理对苹果渣发酵产物中果胶酶活性的影响
2.3.1自然发酵不同菌剂及辅料对未灭菌原料发酵产物中果胶酶活性的影响见表5。
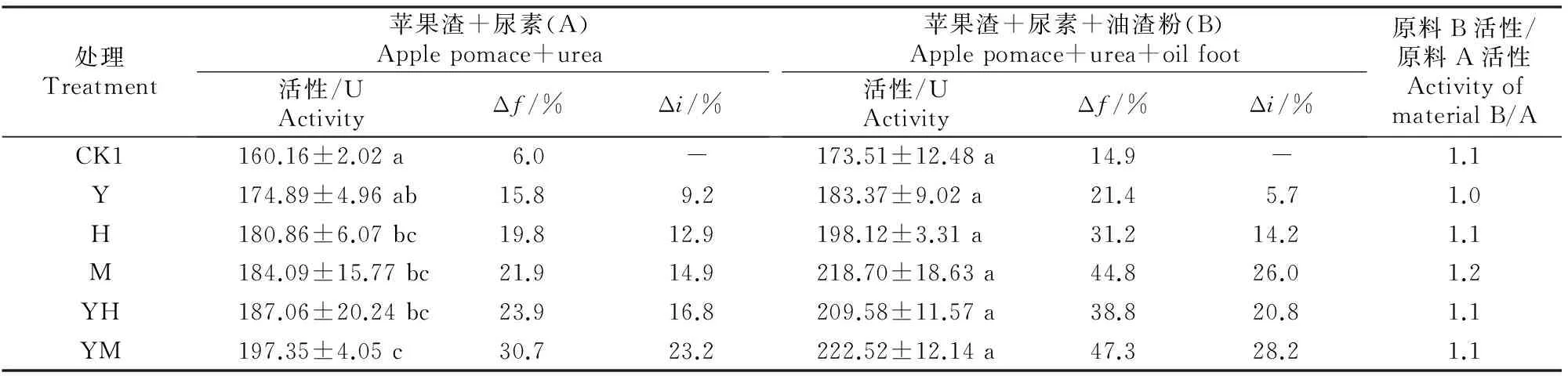
表 5 不同菌剂及辅料对未灭菌原料发酵产物中果胶酶活性的影响Table 5 Effect of different microbial inoculums and auxiliary materials on pectinase activity in fermentation product of unsterilized material
注:未发酵纯苹果渣的果胶酶活性为151.03 U。
Note:The pectinase activity in blank sample was 151.03 U.
由表5可知,在自然发酵条件下,原料A不接种发酵剂而仅靠原料及空气中的微生物自然接种,就可以使苹果渣的果胶酶活性由未发酵纯苹果渣的151.03 U提高到160.16 U,即自然发酵条件下果胶酶活性的发酵增率(Δf)为6.0%;接种酵母菌+米曲霉、酵母菌+黑曲霉A8、米曲霉及黑曲霉A8后,发酵产物中果胶酶活性较未发酵纯果渣分别提高30.7%,23.9%,21.9%和19.8%;与不接种对照处理相比,果胶酶活性的接菌增率(Δi)为9.2%~23.2%。原料B与之类似,特别是在接种酵母菌+米曲霉时,果胶酶活性的发酵增率(Δf)及接菌增率(Δi)分别为47.3%和 28.2%。
2.3.2灭菌发酵由表6可以看出,在灭菌发酵中,发酵能显著增加产物中果胶酶活性。在原料A中,果胶酶活性的发酵增率(Δf)为27.1%~120.6%,其中不接菌处理果胶酶活性的发酵增率(Δf)为 27.1%。接菌处理能大幅提高发酵产物的果胶酶活性,向苹果渣中接种各供试菌株发酵所得果胶酶活性为 235.12~333.21 U,且接种黑曲霉A8、米曲霉、黑曲霉A8+酵母菌和米曲酶+酵母菌均与不接菌对照差异显著(P<0.05),果胶酶活性的接菌增率(Δi)为 22.5%~73.6%,其中接种酵母菌+米曲霉时果胶酶活性较对照增加73.6%。在原料B中,发酵产物中果胶酶活性为203.20~358.47 U,其发酵增率(Δf)为34.5%~137.4%;与不接菌对照相比,果胶酶活性的接菌增率(Δi)为26.4%~76.4%。
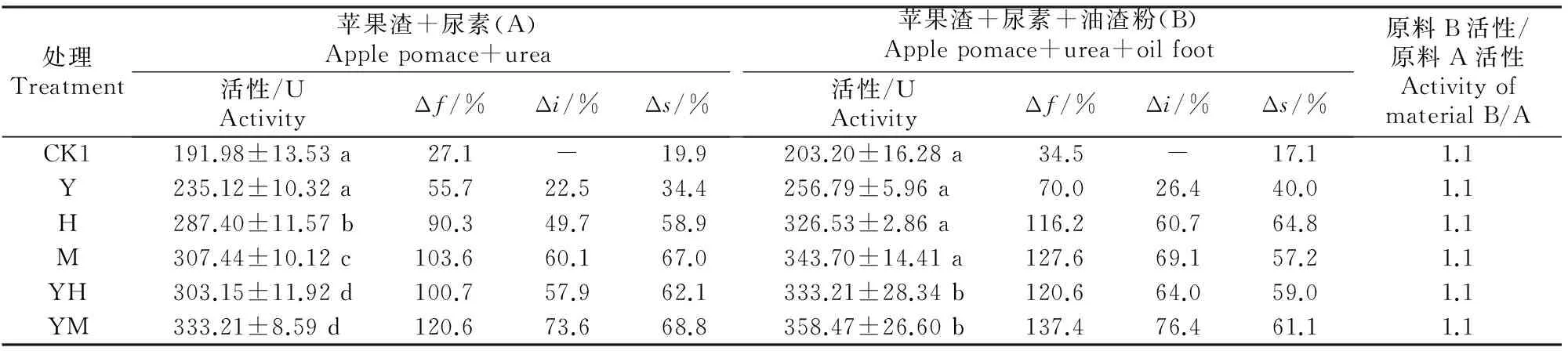
表 6 不同菌剂及辅料对灭菌原料发酵产物中果胶酶活性的影响Table 6 Effect of different microbial inoculums and auxiliary materials on pectinase activity in fermentation product of sterilized material
注:未发酵纯苹果渣果胶酶活性为151.03 U。
Note:The pectinase activity in blank sample was 151.03 U.
2.3.3原料灭菌及添加油渣粉对发酵产物中果胶酶活性的影响由表6中的灭菌增率(Δs)可以看出,灭菌条件下发酵产物的果胶酶活性均明显大于自然发酵,原料A和原料B的灭菌增率(Δs)分别为19.9%~68.8%和17.1%~64.8%,说明灭菌处理能提高发酵产物果胶酶活性。结合表5、表6可知,在自然及灭菌条件下,原料B发酵产物中果胶酶活性均明显高于原料A发酵产物。在自然发酵中,单接米曲霉时原料B中的果胶酶活性是原料A的1.2倍;原料A、原料B的发酵增率(Δf)分别为6.0%~120.6%和14.9%~137.4%,即原料B的发酵增率明显高于原料A,表明原料中添加油渣粉有助于提高发酵产物中的果胶酶活性及其发酵增率。
2.4不同处理对苹果渣发酵产物中植酸酶活性的影响
2.4.1自然发酵不同菌剂及辅料对未灭菌原料的发酵产物中植酸酶活性的影响见表7。
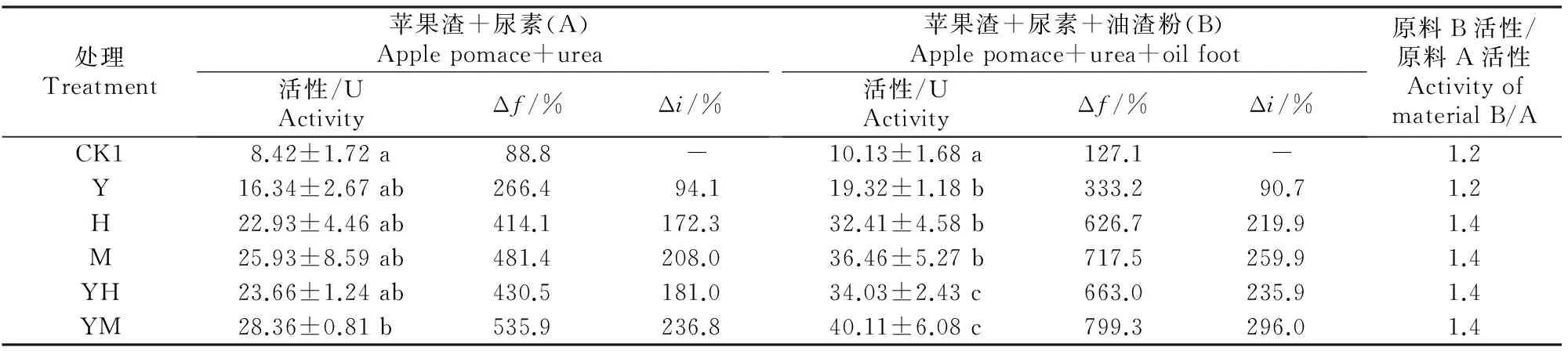
表 7 不同菌剂及辅料对未灭菌原料发酵产物中植酸酶活性的影响Table 7 Effect of different microbial inoculums and auxiliary materials on phytase activity in fermentation product of unsterilized material fermentation products of unsterilized material
注:未发酵纯苹果渣的植酸酶活性为4.46 U。
Note:The phytase activity in blank sample was 4.46 U.
由表7可以看出,原料A在不灭菌和不接种发酵剂的自然条件下,靠原料及空气中的微生物自然接种,可以使苹果渣的植酸酶活性由未发酵纯苹果渣的4.46 U增加到8.42 U,发酵增率(Δf)为 88.8%;接种酵母菌+米曲霉、米曲霉、酵母菌+黑曲霉A8、黑曲霉A8及酵母菌后,发酵产物中植酸酶活性的发酵增率(Δf)分别为535.9%,481.4%,430.5%,414.1%和266.4%;与不接种对照处理相比,植酸酶活性的接菌增率(Δi)为 94.1%~236.8%。原料B与之类似,特别是在接种酵母菌+米曲霉时,植酸酶活性的发酵增率(Δf)及接种增率(Δi)分别为799.3%和296.0%。
2.4.2灭菌发酵由表8可知,在灭菌发酵中,接菌处理能大幅提高发酵产物的植酸酶活性。在原料A中,向苹果渣中接种各供试菌株,发酵产物植酸酶活性为13.37~36.85 U,且米曲霉+酵母菌、米曲霉和黑曲霉A8+酵母菌处理均与不接菌对照植酸酶活性差异显著;植酸酶活性的发酵增率(Δf)为199.8%~726.2%,其中不接菌处理条件下植酸酶活性的发酵增率(Δf)为199.8%;在不同发酵剂处理中,植酸酶活性的接菌增率(Δi)为 58.6%~175.6%,其中接种酵母菌+米曲霉混菌时植酸酶活性较对照增加175.6%,增幅明显。在原料B中,发酵产物中植酸酶活性高达19.45~54.96 U,发酵增率(Δf)为336.1%~1 132.3%,植酸酶活性的接菌增率(Δi)为41.4%~182.6%。
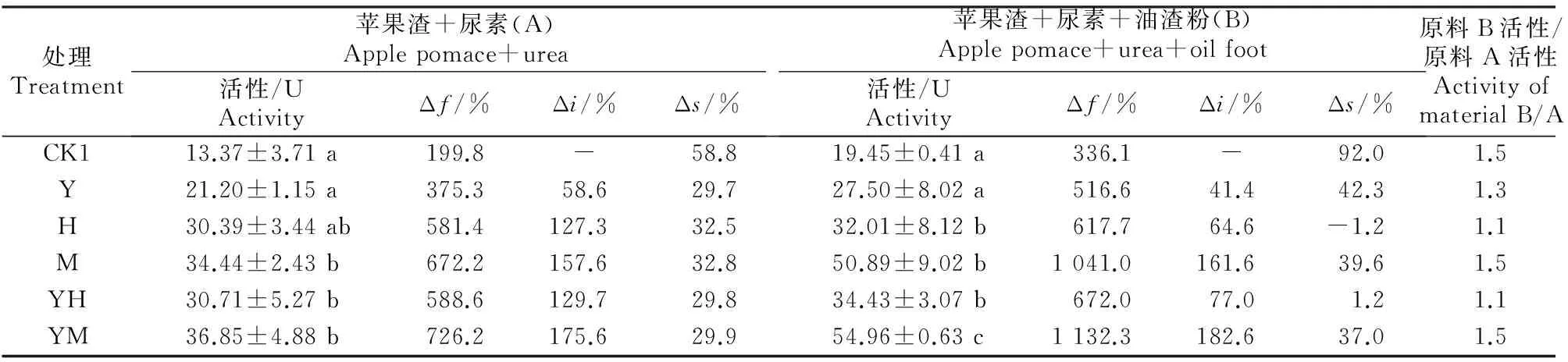
表 8 不同菌剂及油渣粉对灭菌原料发酵产物中植酸酶活性的影响Table 8 Effect of different microbial inoculums and auxiliary materials on phytase activity in fermentation product of sterilized material
注:未发酵纯苹果渣植酸酶活性为4.46 U。
Note:The phytase activity in blank sample was 4.46 U.
2.4.3原料灭菌及添加油渣粉对发酵产物中植酸酶活性的影响由表8中的灭菌增率(Δs)可以看出,除黑曲霉A8处理以外,灭菌条件下发酵产物的植酸酶活性均明显大于自然发酵,原料A和原料B的灭菌增率(Δs)分别为29.7%~58.8%和-1.2%~92.0%,说明灭菌能提高发酵产物植酸酶活性。结合表7、表8可知,在未灭菌及灭菌条件下,原料B发酵产物的植酸酶活性明显高于原料A。在自然发酵中,单接黑曲霉A8、米曲霉及酵母菌+黑曲霉A8、酵母菌+米曲霉混接,原料B发酵产物中的植酸酶活性均为原料A的1.4倍;在灭菌发酵中,单接米曲霉、酵母菌+米曲霉混接处理的植酸酶活性也均为原料A的1.5倍。此外,原料A、原料B的发酵增率(Δf)分别为88.8%~726.2%和 127.1%~1 132.3%,即原料B的发酵增率高于原料A,表明原料中添加油渣粉有助于提高发酵产物中的植酸酶活性及发酵增率。
3讨论与结论
酶是一种由活细胞产生的具有生物催化能力的蛋白质,在动物体内消化与新陈代谢过程中发挥着重要作用。自1975年美国饲料工业首次将酶制剂作为添加剂应用于配合饲料中并取得显著效果后,饲用酶制剂日益受到养殖业的重视。外源加入的饲用蛋白酶可增强动物消化道酶系作用,降解蛋白质成为易被吸收的氨基酸等小分子物质;纤维素酶和果胶酶通过破坏植物细胞壁,将纤维素、半纤维素及果胶等大分子分解成易被消化吸收的小分子糖类,同时释放出植物细胞内的淀粉及蛋白质等,使其充分水解为小分子后被吸收利用[10-12];植酸酶催化植酸盐水解,释放出游离态的无机磷酸盐和肌醇,提高饲料中磷和肌醇等养分的利用率[13]。大量试验表明,将复合酶添加到不同动物的基础日粮中,均可显著提高其生产性能[14-15]。
本研究发现,黑曲霉A8和米曲霉分别与酵母菌混合接种处理苹果渣,其发酵产物4种水解酶活性均高于2种霉菌单独接种,其主要原因在于酵母菌通过利用2种霉菌水解酶水解形成的小分子糖等产物,解除了这些小分子糖对纤维素酶等水解酶合成的阻遏作用[16],促进了水解酶的大量合成。供试发酵菌种中,黑曲霉A8具有很强的纤维素酶、果胶酶、糖化酶、植酸酶及蛋白酶等水解酶的合成能力,米曲霉合成蛋白酶及淀粉酶的能力也很强,而酵母菌则不具备对上述水解酶的合成能力。在固态发酵过程中,黑曲霉A8分泌到固态基质中的纤维素酶可水解发酵原料中的纤维素生成葡萄糖,葡萄糖等小分子糖的大量累积可阻遏纤维素酶的进一步合成,导致酶活性降低。而酵母菌则可利用这些葡萄糖,消除或减弱葡萄糖对纤维素酶合成的阻遏作用,促进纤维素酶的合成,从而提高发酵产物的纤维素酶活性[16]。其他3种酶合成也有类似作用。
此外,本研究还发现,发酵原料灭菌处理可明显提高发酵产物中4种水解酶的活性,其原因之一是灭菌处理使发酵原料充分熟化,原因之二是灭菌处理消除了发酵原料中杂菌对接入菌生长的竞争影响,这两种效应均有利于接入菌的大量生长繁殖,进而增加了接入菌合成的水解酶量,提高了发酵产物中4种水解酶的活性。以苹果渣为主料,通过固态发酵生产出具有较高蛋白质及动物必需氨基酸含量[16]且含有多种水解酶的发酵饲料,是苹果渣高效利用的重要途径之一,但目前的研究主要集中在提高发酵饲料中蛋白质含量的工艺优化及氨基酸变化[17-21]方面,对发酵饲料中多种具有重要作用的水解酶活性的关注较少,更缺乏深入研究。本研究系统探讨了不同发酵剂、辅料及灭菌与否对苹果渣固态发酵产物中蛋白酶、纤维素酶、果胶酶及植酸酶酶活性的影响,结果表明,以供试发酵剂进行固态发酵获得的苹果渣发酵产物中含多种活性较高的水解酶系,所得产物用于饲料原料时不仅可为动物提供优质蛋白质和多种动物必需氨基酸,还可提供多种水解酶系,兼有复合酶制剂的部分功能。因此,不能将苹果渣发酵产物单纯视为单细胞蛋白质原料,而是兼备优质蛋白质及复合酶制剂多种功能的饲料原料。若将该发酵产物以一定比例添加到饲料中,在发酵产物及其中复合水解酶的共同作用下,动物饲料中的一些抗营养因子将被破坏,饲料的消化利用率得到提高,对加快动物生长速度、提高免疫力及健康水平均有重要的促进作用,本研究结果将为苹果渣发酵饲料品质评价及发酵工艺优化提供新的科学依据。
[参考文献]
[1]Oxenboll K M,Pontoppidan K,Fru-Nji F.Use of a protease in poultry feed offers promising environmental benefits [J].International Journal of Poultry Science,2011,10(11):842-848.
[2]王丽媛,仇农学.固态发酵技术提高苹果渣附加值的应用研究 [J].生物技术通报,2009(7):63-67.
Wang L Y,Qiu N X.The research progress of application of solid-state fermentation technology in enhancing the added value of apple pomace [J].Biotechnology Bulletin,2009(7):63-67.
[3]王伟军,李延华,张兰威,等.利用苹果渣生产单细胞蛋白的研究 [J].中国酿造,2008(20):55-57.
Wang W J,Li Y H,Zhang L W,et al.Study on producing single cell protein by apple pomace [J].China Brewing,2008(20):55-57.
[4]武运,李焕荣,陶咏霞,等.发酵苹果渣生产菌体蛋白饲料工艺的研究 [J].中国酿造,2009(1):83-86.
Wu Y,Li H R,Tao Y X,et al.Fermentation conditions of cell protein feed produced by apple pomace [J].China Brewing,2009(1):83-86.
[5]张高波,李巨秀,来航线,等.菌种对苹果渣发酵饲料中蛋白酶活、纤维素酶活及总酚含量的影响 [J].食品与发酵工业,2013,39(11):118-123.
Zhang G B,Li J X, Lai H X,et al.The effect of the strains on proteinase activity,cellulase activity and total phenolic content in apple pomace fermented feed [J].Food and Fermentation Industries,2013,39(11):118-123.
[6]Aoyama M,Yasuda M,Nakachi K,et al.Soybean-milk-coagulating activity ofBacilluspumilusderives from a serine proteinase [J].Applied Microbiology and Biotechnology,2000,53(4):390-395.
[7]Konig J,Grasser R,Pikor H,et al.Determination of xylanase,β-glucanase,and cellulase activity [J].Analytical and Bioanalytical Chemistry,2002,374(1):80-87.
[8]李忠福,徐建国.分光光度计法测定果胶酶活性方法的研究 [J].黑龙江医药,2002,15(6):428-430.
Li Z F,Xu J G.Dtermination of pectinase activity by spectrophotometry [J].Heilongjiang Medical Journal,2002,15(6):428-430.
[9]陈琛.植酸酶活性测定方法的研究进展 [J].中国饲料,2010(20):16-18,22.
Chen C.Advance on determination of phytase activity [J].China Feed,2010(20):16-18,22.
[10]Bhat M K.Cellulases and related enzymes in biotechnology [J].Biotechnology Advances,2000,18(5):355-383.
[11]Khattak F M,Pasha T N,Hayat Z,et al.Enzymes in poultry nutrition [J].Journal of Animal and Plant Sciences,2006,16(1/2):1-7.
[12]钟发刚,王新华.饲用纤维素酶研究进展 [J].中国微生态学杂志,2002,14(5):308-309.
Zhong F G,Wang X H.Advance on the research of feeding cellulase [J].Chinese Journal of Microecology,2002,14(5):308-309.
[13]李海宁.饲料酶制剂种类及应用前景 [J].四川畜牧兽医, 2004,30(9):59-60.
Li H N.Sorts and application future of feeding enzyme [J].Hubei Journal of Animal and Veterinary Sciences,2004,30(9):59-60.
[14]邓永平,艾瑞波,刘晓兰.饲用复合酶开发与应用研究进展 [J].安徽农业科学,2009,37(26):12566-12568.
Deng Y P,Ai R B,Liu X L.Research progress on development and application of feeding compound enzymes [J].Journal of Anhui Agriculture Science,2009,37(26):12566-12568.
[15]李红玉,杨桂英.复合酶制剂的生理功能及在动物生产中的应用 [J].饲料研究,2006(6):57-58.
Li H Y,Yang G Y.Physiological function and application of composite enzyme in animal production [J].Feed Research,2006(6):57-58.
[16]司美茹,薛泉宏,蔡艳.混合发酵对纤维素酶和淀粉酶活性的影响 [J].西北农林科技大学学报(自然科学版),2002,30(5):69-73.
Si M R,Xue Q H,Cai Y.The effect of mixed fermentation on cellulose and amylase activities [J].Journal of Northwest A&F University(Natural Science Edition),2002,30(5):69-73.
[17]陈姣姣,来航线,马军妮,等.发酵剂及玉米浆对苹果渣发酵饲料氨基酸含量及种类的影响 [J].饲料工业,2014,35(6):42-46.
Chen J J,Lai H X,Ma J N,et al.The effect of fermentation agents and corn steep liquor on amino acids contents and varieties in apple pomace fermented feed [J].Feed Industry,2014,35(6):42-46.
[18]贺克勇,薛泉宏,司美茹,等.苹果渣发酵饲料蛋白质含量的影响因素研究 [J].西北农林科技大学学报(自然科学版),2004,32(4):83-87.
He K Y,Xue Q H,Si M R,et al.Research into influential factors on protein content of fermented feed with pomace [J].Journal of Northwest A&F University(Natural Science Edition),2004,32(4):83-87.
[19]贺克勇,薛泉宏,来航线,等.氮素及原料配比对苹果渣发酵饲料纯蛋白质含量和氨基酸组成的影响 [J].饲料工业,2004,25(8):34-37.
He K Y,Xue Q H,Lai H X,et al.The effect of nitrogen and material ratiomaterial ratio on the content of pure protein and amino acids in apple pomace fermented feed [J].Feed Industry,2004,25(8):34-37.
[20]贺克勇,杨帆,薛泉宏,等. 鲜苹果渣蛋白饲料发酵工艺研究 [J].西北农林科技大学学报(自然科学版),2007,35(11):90-94.
He K Y,Yang F,Xue Q H,et al.Studies on fermenting technology of protein-fodder from pomace [J].Journal of Northwest A&F University(Natural Science Edition),2007,35(11):90-94.
[21]贺克勇,杨帆,薛泉宏,等.固态复合发酵剂用于苹果渣蛋白饲料发酵效果研究 [J].饲料工业,2005,26(9):9-11.
He K Y,Yang F,Xue Q H,et al.The effect of soild-state composite fermentation agents on the quality of apple pomace fermented feed [J].Feed Industry,2005,26(9):9-11.
DOI:网络出版时间:2016-06-0816:2110.13207/j.cnki.jnwafu.2016.07.027
[收稿日期]2014-11-19
[基金项目]陕西省科技统筹创新工程项目(2015KTTSNY03-06);国家科技支撑计划项目(2012BAD14B11)
[作者简介]任雅萍(1985-),女,陕西渭南人,硕士,主要从事微生物资源利用研究。E-mail:renyaping19850618@163.com [通信作者]薛泉宏(1957-),男,陕西白水人,教授,主要从事微生物资源利用研究。E-mail:xuequanhong@163.com
[中图分类号]S816.46
[文献标志码]A
[文章编号]1671-9387(2016)07-0193-08
Effect of fermention conditions on the four hydrolases in apple pomace fermented feed
REN Yapinga,GUO Qiaob,LAI Hangxianb,CHEN Jiaojiaob,XUE Quanhongb
(aCollegeofLifeScience,bCollegeofNatrualResourcesandEnvironment,NorthwestA&FUniversity,Yangling,Shaanxi712100,China)
Abstract:【Objective】 The study investigated the effects of microbial inoculums,auxiliary materials and sterilization of material on activities of 4 important hydrolases (protease,cellulose,pectinase and phytase) in fermentation product of apple pomace.【Method】 Apple pomace with different auxiliary materials (urea or urea and oil residue) was fermented with three fungal species,yeast,Aspergillus niger A8 and Aspergillus oryzae.The effects of microbial inoculums (single-strain fermentation and mixed-strains fermentation),auxiliary materials and sterilization of material on activities of 4 important hydrolases (protease,cellulose,pectinase and phytase) were analyzed by colorimetric spectroscopy.【Result】 The activities of the 4 hydrolases were increased significantly in fermentation product of apple pomace.The activities of all 4 hydrolases in mixed-strains fermentation product were much higher than in the single-strain fermentation product.Adding oil residues and sterilization significantly improved the activities of protease,cellulose,pectinase and phytase.In the fermentation product of sterilized material containing oil residues,the activities of protease,cellulose,pectinase and phytase reached 58.85-368.43 U,3 617.53-6 278.22 U,203.20-358.47 U,and 19.45-54.96 U,respectively.The fermentation rates were increased by 67.7%-949.6%,13.8%-97.6%,34.5%-137.4%,and 336.1%-1 132.3%,the activities with microbial inoculation were increased by 109.4%-526.0%,2.3%-73.5%,26.4%-76.4%,and 41.4%-182.6%,and the activities in fermentation product of sterilized material were 22.2%-118.6%,8.3%-63.9%,17.1%-64.8%,and 1.2%-92.0%,respectively.【Conclusion】 Mixed-strains fermentation,auxiliary materials (oil residues) and sterilization of material significantly improved activities of protease,cellulose,pectinase and phytase in fermentation product of apple pomace.
Key words:apple pomace;fermented feed;protease activity;cellulose activity;pectinase activity;phytase activity
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160608.1621.054.html
