
天目山自然保护区典型阔叶林的光合特性
2016-07-28 02:03:12马锦丽舒海燕吴孟霖方成圆孙文文陈晓峰牛晓栋龚莎莎
福建农林大学学报(自然科学版) 2016年4期
马锦丽, 江 洪,2, 舒海燕, 吴孟霖, 方成圆, 孙文文, 陈晓峰, 牛晓栋, 龚莎莎
(1.浙江农林大学国际生态研究中心,浙江 杭州 311300;2.南京大学国际地球系统科学研究所,江苏南京 210093;3.西南大学生命科学学院三峡库区生态环境教育部重点实验室,重庆 400715)
天目山自然保护区典型阔叶林的光合特性
马锦丽1, 江洪1,2, 舒海燕3, 吴孟霖1, 方成圆1, 孙文文1, 陈晓峰1, 牛晓栋1, 龚莎莎1
(1.浙江农林大学国际生态研究中心,浙江 杭州 311300;2.南京大学国际地球系统科学研究所,江苏南京 210093;3.西南大学生命科学学院三峡库区生态环境教育部重点实验室,重庆 400715)
摘要:对天目山自然保护区内典型阔叶林树种交让木(Daphniphyllum macropodum)、小叶青冈(Cyclobalanopsis myrsinaefolia)、中国绣球(Hydrangea chinensis Maxim),在秋冬季节的光合作用光响应、叶绿素荧光以及相对叶绿素含量进行了测定分析.结果表明,(1)2个季节中,光饱和点(LSP)、最大净光合速率(Amax)、表观量子效率(AQY)、实际光化学效率(Fv′/Fm′)、实际量子产量(Yield)、表观电子传递速率(ETR)均是先减小后增大,均以交让木最高.光补偿点(LCP)秋季以交让木最高,小叶青冈次之,中国绣球最低;冬季交让木高于小叶青冈;暗呼吸速率(Rd)总体上交让木最低,秋季中国绣球最高,冬季小叶青冈较高.光化学猝灭系数(qP)秋季中国绣球最高,其次是小叶青冈,交让木最低;冬季小叶青冈高于交让木.非光化学猝灭系数(NPQ)的变化趋势与qP相同.胁迫的相应机制:交让木为关闭一部分光合系统,降低暗呼吸速率;小叶青冈和中国绣球主要是将过剩的光能耗散为热能.(2)交让木和小叶青冈的相对叶绿素含量均表现为秋季<冬季,且差异显著;同一时间内相对叶绿素含量表现为交让木>小叶青冈>中国绣球,差异极显著.净光合速率(Pn)表现为交让木>小叶青冈;秋季中国绣球Pn位于二者之间.这与相对叶绿素含量的变化趋势基本一致,都是随着温湿度的变化呈先下降后上升的变化规律,但均为正值.
关键词:天目山; 阔叶林; 光响应; 叶绿素荧光; 相对叶绿素含量
绿色植被碳循环是陆地生态系统碳循环的基础,了解植物光合特性是了解植物碳循环的重要途径[1-3].同时植物光合特性对环境因子的响应,是评价植物在一定环境条件下的生长能力及适应能力的基础[4-9].浙江天目山国家级自然保护区地处我国东南沿海丘陵山区中亚热带北缘、北亚热带南缘.其气候具有中亚热带向北亚热带过渡的特征,具有典型的中亚热带森林生态系统.天目山植物区系古老,成分复杂,种类丰富,是当今华东地区植被保存较为完好的地区之一[10],被誉为“大树华盖闻九州”的天然植物园.常绿阔叶林是天目山的地带性植被,集中分布于禅源寺附近和海拔850~1 100 m的山坡和沟谷[11].对天目山的研究多集中于群落特征、空间分布,以及生物量等层面[12-15].而叶绿素荧光、相对叶绿素含量等生理特性的研究尚未见报道.本研究对天目山自然保护区内典型阔叶林树种的生理生态指标进行研究,测定其气体交换参数、叶绿素荧光和相对叶绿素含量等参数,并探讨其在秋冬季不同时间的动态变化规律,以及典型树种对环境变化的适应性,旨在为天目山自然保护区植被的保护、修复、重建等提供参考.
1材料与方法
1.1研究地区自然概况
天目山国家级自然保护区位于浙江省临安市境内,地理位置30°18′30″~ 30°24′55″N,119°24′47″~119°28′27″E,面积4 284 hm2.主峰仙人顶海拔1 506 m.其中林地面积4 261.1 hm2,占99.5%,森林覆盖率98.1%.林地面积中乔木林3 711.3 hm2,占88.7%;竹林面积474.8 hm2,占11.3%.土壤厚度约为50 cm,多为黄壤土、棕黄壤土.气候具有中亚热带向北亚热带过渡的特征,四季分明,雨水充沛,光照适宜且复杂多变.保护区年平均气温8.8~14.8 ℃,最冷月平均气温-2.6~3.4 ℃,极端最低气温-20.2~-13.1 ℃,最热月平均气温19.9~28.1 ℃,极端最高气温29.1~38.2 ℃,无霜期209~235 d,年雨日159.2~183.1 d,年雾日64.1~255.3 d,年降水量1 390~1 870 mm,年太阳辐射3 270~4 460 MJ·m-2,相对湿度76%~81%.常绿落叶阔叶混交林分布于海拔850~1 100 m,常绿乔木主要有细叶青冈(Cyclobalanopsisgracilis)、石栎(Lithocarpusglaber)、交让木(Daphniphyllummacropodum)等;落叶乔木有青钱柳(Cyclocaryapaliurus)、化香(Platycaryastrobilacea)、枫香(Liquidambarformosana)、天目木姜子(Litseaauriculata)、短柄枹(Quercusglandulifera)等;灌木有柃木(EuryajaponicaThunb)有箬竹(Indocalamustessellatus)、马银花(Rhododendronovatum)等;混有针叶林有柳杉(Cryptomeriafortunei)林、金钱松(Pseudolarixamabilis)林及黄山松(Pinustaiwanensis)林等,组成较复杂的森林类型.这些森林群落大多数处于较稳定状态,达到了演替顶级.调查的2块典型样地林分土壤为山地黄壤,土层深度约100 cm;pH值5~6,枯枝落叶层厚达10~20 cm.碳通量观测塔安装在研究区的一块常绿落叶阔叶混交林样地(30°20′59″N,119°26′13.2″E)内,海拔1 139 m,坡度6.6°左右,坡向南偏西16°.主要乔木有小叶青冈(C.myrsinifolia)、交让木(D.macropodum)、小叶白辛树(Pterostyraxcorymbosus)、短柄枹(Quercusglandulifera)、青钱柳(C.paliurus)、天目槭(Acersinopurpurascens)、秀丽槭(Acerelegantulum)、糙叶树(Aphanantheaspera)等,林龄140 a,郁闭度0.7,林分密度3 125株·hm-2.林分为复层结构,分3层,第1层:15 m以上的乔木,约占3.2%.第2层:8~14 m的乔木,约占43.2%.第3层:乔木均在8 m以下.优势树种为小叶青冈(C.myrsinifolia)、交让木(D.macropodum)、小叶白辛树(P.corymbosus)等.小叶青冈活立木平均高度9.2 m,胸径24.1 cm;交让木活立木平均高度5.1 m,胸径7.8 cm;小叶白辛树活立木平均高度11.2 m,胸径20.2 cm.小乔木或灌木主要有中国绣球(HydrangeachinensisMaxim)、红脉钓樟(Linderarubronervia)、微毛柃(Euryahebeclados)、荚莲(Viburnum)、大青(Clerodendrumcyrtophyllum)、浙江大青(C.kaichianum)、野鸦椿(Euscaphisjaponica)、山胡椒(L.glauca)、鸡毛竹(Shibataeachinensis)、紫竹(Phyllostachysnigra)、牛鼻栓(Fortuneariasinensis)、四照花(Dendrobenthamiajaponicavar.chinensis)等.
1.2光响应曲线的测定及参数估算
试验分别于2014年10月、11月上旬(秋季)和12月、2015年1月上旬(冬季)晴朗的天气下进行.选取3个树种的平均木:小叶青冈高9 m,胸径24.1 cm;交让木高5 m,冠幅7.8 cm;中国绣球高2 m,胸径1.8 m.选取冠层阳面健康成熟的功能叶进行测定.采用 Li-6400XT便携式光合测定系统(LI-COR, Inc., USA)测定叶片的光响应曲线.使用开放气路,空气流速为0.5 L·min-1,叶片温度25 ℃,叶室中相对湿度70%~75%,CO2浓度 380 μmol·mol-1.诱导光强约为 2 000 μmol·m-2·s-1,诱导时间30 min.光强由强到弱依次设定为1 500、1 200、1 000、800、600、400、200、150、100、50、30、20、10、0 μmol·m-2·s-1.测定时每一光强下最长等待时间4 min,最短等待时间2 min.每次测定3个样叶,每个树种重复3次,结果取平均值.依据非直角双曲线模型,拟合Pn-PAR的曲线方程.并据此用Photosyn Assistant软件计算最大净光合速率(Amax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)和暗呼吸速率(Rd),得到胞间二氧化碳浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)、水分利用效率(WUE),其中WUE=Pn/Tr,Ls=1-Ci/Ca.
1.3叶绿素荧光参数的测定
选取的叶片同上,使用PAM-2500(德国WALZ公司生产)测定其叶绿素荧光参数,充分暗适应后,以弱调制测量光(0.05 μmol·m-2·s-1)测定初始荧光(Fo),以强饱和光(6 000 μmol·m-2·s-1)激发最大荧光(Fm) ,闪光2 s.通过以下公式计算最大光化学效率(Fv/Fm)、潜在活性(Fv/Fo)、实际最大光化学效率(Fv′/Fm′).
Fv为可变荧光且Fv=Fm-Fo;Fv/Fm=(Fm-Fo)/Fm;Fv/Fo=(Fm-Fo)/Fo;Fv′/Fm′=(Fm′-Fo′)/Fm′.
多次强饱和闪光脉冲(6 000 μmol·m-2·s-1,脉冲时间2 s),直至光下最大荧光(Fm′)稳定,读取光化学猝灭系数(qP)、非光化学猝灭系数(NPQ)、PSⅡ的实际量子产量(Yield)和表观电子传递速率(ETR)值.
1.4相对叶绿素含量的测定
相对叶绿素含量采用日本产的SPAD-502手持式叶绿素测定仪测定.以上述试验采用的植株为测定对象,每株选择35片叶片,在各个叶片中脉两侧均匀选取5个点,取平均值作为该叶片的相对叶绿素含量.
1.5数据处理
采用Excel 2007整理数据并做图,用SPSS 13.0统计分析软件检验数据,显著性检验采用LSD法(α=0.05) ;然后用One-way ANOVA进行数据分析.对交让木和小叶青冈进行T检验.用回归分析法检测3个树种的净光合速率(Pn)与环境因子的相关性.
2结果与分析
2.1典型树种不同季节光响应参数特性
从表1-3可知,交让木的LSP表现为 1月>10月>12月>11月,11月极显著低于其余各月(P<0.01);小叶青冈的LSP表现为12月>10月=1月>11月,差异性同交让木;中国绣球的LSP表现为10月>11月,差异极显著(P<0.01).同月份而言,10月、11月,交让木、中国绣球的LSP之间差异不显著(P>0.05),而极显著高于小叶青冈(P<0.01);12月和1月份,交让木LSP极显著高于小叶青冈(P<0.01).总之,3个典型树种的LSP,在秋季以中国绣球最高,在冬季以交让木最高,在秋冬季均以小叶青冈最低.
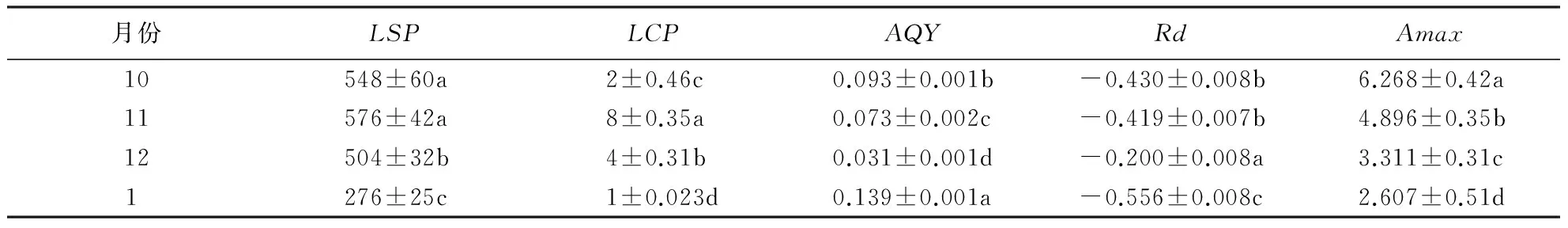
表1 交让木不同季节的光响应参数1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).
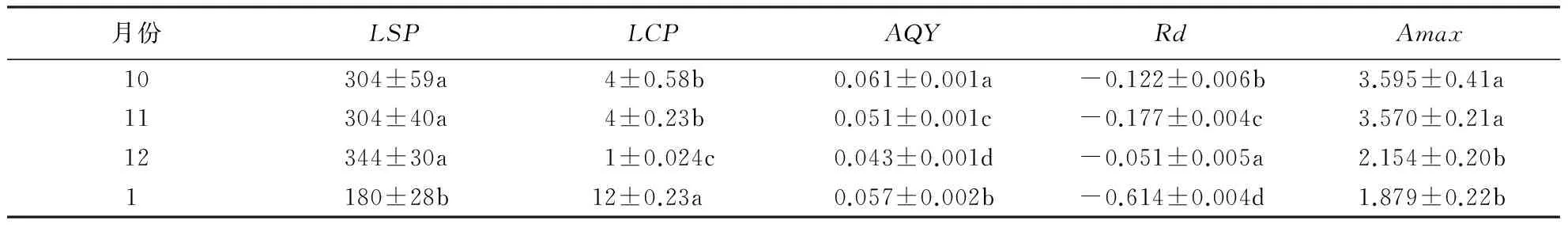
表2 小叶青冈不同季节的光响应参数1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).

表3 中国绣球不同季节的光响应参数1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).
不同月份交让木的LCP表现为1月>12月>10月>11月,各月之间差异极显著(P<0.01);小叶青冈的LCP表现为11月>10月=1月>12月,10月与1月之间差异不显著(P>0.05),其余各月之间差异极显著(P<0.01); 中国绣球的LCP表现为10月>11月,差异不显著(P>0.05).同一月份不同树种的LCP,10月、11月都表现为小叶青冈>交让木>中国绣球,其中,10月三树种间差异极显著(P<0.01);11月表现为交让木与小叶青冈之间差异不显著(P>0.05),但都极显著低于中国绣球(P<0.01);12月和1月均表现为交让木>小叶青冈,且均是差异极显著(P<0.01).总体上,在秋季小叶青冈的LCP最高,中国绣球最低;在冬季则表现为交让木>小叶青冈.
交让木的AQY表现为 11月>10月>1月>12月,差异均达到极显著水平(P<0.01);小叶青冈的AQY表现为 10月>11月>1月>12月,各月之间差异极显著(P<0.01);中国绣球的AQY表现为 10月极显著高于11月(P<0.01).同一月份来看,10月的AQY表现为交让木>中国绣球>小叶青冈,差异极显著(P<0.01);11月表现为交让木>中国绣球>小叶青冈,差异极显著(P<0.01);12月表现为小叶青冈>交让木,差异极显著(P<0.01);1月表现为交让木>小叶青冈,差异极显著(P<0.01).总之,3个典型树种的AQY总体上看以交让木为最高.
3个树种的Rd10月表现为中国绣球>小叶青冈>交让木,差异极显著(P<0.01);11月则表现为中国绣球>交让木>小叶青冈,差异极显著(P<0.01);12月和1月均表现为小叶青冈>交让木,差异极显著(P<0.01).同一树种不同月份而言,交让木的Rd表现为 12月>1月>10月>11月,1月与10月差异不显著(P>0.05),其余各月之间差异均极显著(P<0.01);小叶青冈则表现为12月>10月>1月>11月,显著性同交让木;中国绣球的Rd表现为10月>11月,差异极显著(P<0.01).由此可知,3个树种的Rd在秋季以中国绣球最高;冬季则是小叶青冈较高,交让木较低.
不同季节交让木和小叶青冈的Amax表现一致,均为10月>1月>12月>11月.其中交让木Amax11月与12月差异显著(P<0.05),其余各月之间差异极显著(P<0.01);小叶青冈Amax则表现为10月与1月之间差异不显著(P>0.05),11月与12月之间差异不显著(P>0.05),但前者极显著高于后者(P<0.01);中国绣球Amax则表现为10月>11月,差异极显著(P<0.01).同一月份时, 3个树种的Amax在秋季均表现为交让木>中国绣球>小叶青冈,交让木极显著高于中国绣球和小叶青冈(P<0.01),后两者之间差异显著(P<0.05);冬季Amax则表现为交让木>小叶青冈,差异极显著(P<0.01).由此可见,在 4个月份中,交让木的Amax均最高;小叶青冈的Amax最低.
2.2典型树种不同季节的叶绿素荧光特性
从表4~6可知,同一树种而言,交让木和小叶青冈的最大光化学效率(Fv/Fm) 表现为10月>1月>11月>12月,其中,交让木表现为12月与其他各月差异极显著(P<0.01),其他3个月之间差异不显著(P>0.05);小叶青冈表现为10月与11月之间差异显著(P<0.05),与其他月份之间差异均极显著高于12月(P<0.01).同一月份不同树种来看,10月和12月交让木和小叶青冈Fv/Fm的变化趋势相同,表现为小叶青冈>交让木,11月表现为中国绣球>交让木>小叶青冈,1月表现为交让木>小叶青冈,各个月份差异均不显著(P>0.05).由此可见,3个树种潜在最大光合能力,秋季以中国绣球最高;交让木和小叶青冈都是在10月最高,12月最低.
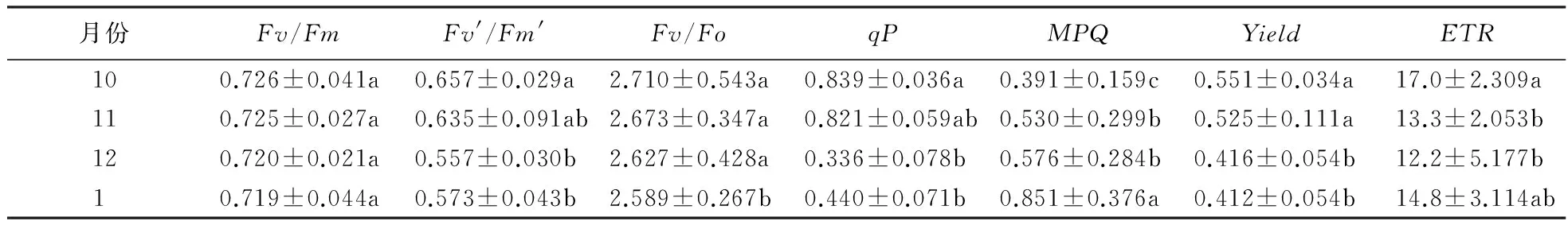
表4 交让木不同季节的叶绿素荧光1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).
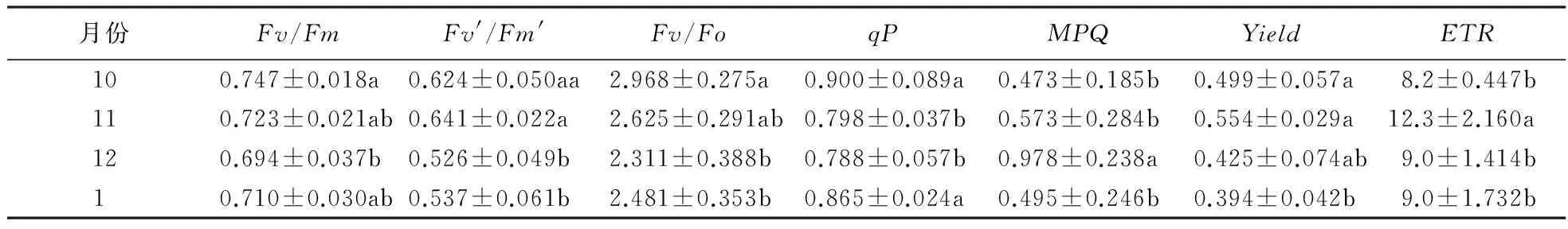
表5 小叶青冈不同季节的叶绿素荧光1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).

表6 中国绣球不同季节的叶绿素荧光1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).
Fv′/Fm′总体上表现为秋季>冬季,交让木表现为10月>11月>1月>12月,秋冬季内两月之间差异不显著(P>0.05),但10月与11月之间差异极显著高于12月(P<0.01),显著高于1月(P<0.05);小叶青冈表现为11月>10月>1月>12月, 各月之间差异均不显著(P>0.05).10月和1月Fv′/Fm′均表现为交让木>小叶青冈;11月表现为中国绣球>交让木>小叶青冈;12月表现为交让木>小叶青冈,差异均不显著(P>0.05).总体上,秋季Fv′/Fm′以中国绣球最高;交让木比小叶青冈高,二者均是在12月达到最低值.
同一月份的(Fv/Fo)在10月和12月的变化趋势相同,表现为交让木>小叶青冈,11月表现为中国绣球>交让木>小叶青冈,各月之内不同树种间差异均不显著(P>0.05).交让木不同月份的Fv/Fo表现为1月>10月>11月>12月,1月、10月、11月之间差异不显著(P>0.05),但都极显著高于12月(P<0.01);小叶青冈表现为10月>11月>1月>12月,秋季高于冬季,11月与1月之间差异不显著(P>0.05),其余各月之间差异均极显著(P<0.01).由此可见,交让木和小叶青冈的Fv/Fo均是在12月达到最低值.
qP在不同树种、不同月份下的变化趋势差别较大,交让木表现为10月>11月>12月>1月,逐月递减,10月、11月、12月之间差异不显著(P>0.05),但都极显著高于1月(P<0.01);小叶青冈则表现为12月>11月>1月>10月,12月与11月之间差异极显著高于10月(P<0.01),12月极显著高于1月(P<0.01),11显著高于1月(P<0.05).4个月份相比,交让木和小叶青冈的qP均表现为小叶青冈>交让木,且秋季两月份间差异极显著(P<0.01),冬季两月份之间差异不显著(P>0.05),其中,11月以中国绣球最低,其他3个月均以交让木最低.总体而言,交让木qP表现为秋季>冬季;小叶青冈的qP则表现为12月最高,10月最低.小叶青冈的qP大于交让木.
NPQ在两树种不同月份下的变化趋势差别较大,交让木表现为1月>10月>11月>12月,1月显著高于10月(P<0.05),极显著高于11和12月(P<0.01),10、11、12三个月之间差异不显著(P>0.05);小叶青冈则表现为10月>1月>11月>12月,10月与1月之间差异显著(P<0.05),且都极显著高于12月(P<0.01),11月与1、12月之间差异不显著(P>0.05),与10月差异极显著(P<0.01).4个月份中NPQ冬季均表现为交让木>小叶青冈,2个月之间差异都不显著(P>0.05);秋季均表现为交让木最低,11月是中国绣球最高,3个树种间差异不显著(P>0.05),10月交让木与小叶青冈间差异极显著(P<0.01).总而言之,交让木NPQ在秋季最低,在冬季最高,最低值都是在12月出现.
交让木Yield的变化趋势与qP相同,表现为秋季>冬季,10月与11月之间差异不显著(P>0.05),显著高于12月(P<0.05),1月与10月间差异极显著(P<0.01),高于12月和1月;小叶青冈则表现为11月>12月>10月=1月,11月极显著高于1月和10月(P<0.01),与12月差异不显著(P>0.05),12月、1月、10月之间差异不显著(P<0.05).10月Yield与1月的均表现为交让木>小叶青冈,10月二者差异极显著(P<0.01),1月差异不显著(P>0.05);11月和12月Yield则是小叶青冈最高,11月差异极显著(P<0.01),12月差异不显著(P>0.05).可见,交让木和小叶青冈的Yield均是在1月达到最低值.
10月、12月、1月的ETR均表现为交让木>小叶青冈,11月表现为小叶青冈>交让木>中国绣球,其中,10月、11月树种间差异极显著(P<0.01),12月、1月差异不显著(P>0.05).同一树种不同月份的变化趋势差别很大,交让木表现为10月>1月>11月>12月,各月之间差异不显著(P>0.05);小叶青冈则表现为11月>10月>1月>12月,12月和1月差异不显著(P>0.05),其余各月之间差异极显著(P<0.01).交让木的ETR大于小叶青冈,且二者均是在12月达到最低值.
2.3典型树种不同季节相对叶绿素含量特性
从表7可知,交让木相对叶绿素含量表现为12月>1月>11月,3个月之间差异极显著(P<0.01);小叶青冈则表现为12月>11月>1月,12月与1月差异显著(P<0.05).同一月份而言,11月表现为交让木>小叶青冈>中国绣球,差异极显著(P<0.01);12月和1月均表现为交让木>小叶青冈,同一月份两树种间差异极显著(P<0.01).由此可见,总体上交让木相对叶绿素含量最高,在12月达到最高值.

表7 典型树种不同季节的相对叶绿素含量1)
1)数据为平均值±标准误差,不同小写字母表示差异显著(P<0.05).
2.4典型树种Pn和环境因子的关系
从图1~3可知:随着时间的变化,交让木的Pn、Gs、Ci、Ls、Tr的变化趋势一致,均表现为先降低后有所升高;WUE则呈上升趋势;小叶青冈的Tr和WUE的走势基本上与交让木一致;中国绣球的Pn降低时,Gs降低,Ci升高,同时Tr和WUE均下降.
3讨论
研究各树种光合作用的季节变化特性,对分析其光合生产和干物质积累以及在森林生态系统碳循环中的作用具有重要意义[16].本研究结果表明,正常情况下,交让木、小叶青冈和中国绣球均具有较高的LSP和较低的LCP,即同一生境下,3种植物对弱光和强光的利用能力都较强.交让木具有较高的LSP和较低的LCP,说明其生态幅更为宽广,这与刘圣恩等[17]的研究结果吻合.AQY反映了植物在弱光下吸收、转化和利用光能的能力[18].本研究结果显示,交让木和小叶青冈的AQY显著高于中国绣球,小叶青冈和中国绣球的AQY均为0.03~0.07,交让木则在10月、12月和1月都超过了0.07(上限),这可能是因为它吸收、转换光能的色素蛋白复合体较多,故利用弱光的能力更强[19].Amax是衡量植物叶片光合作用潜力的重要指标[20],本研究结果表明,秋冬季交让木的Amax最大,故光合潜力最大.植物的呼吸作用是植物生长所必需的生理代谢活动[21],秋季时中国绣球的暗呼吸速率最低,说明它能利用林下环境,通过减少呼吸消耗来积累有机物质;冬季小叶青冈的暗呼吸速率比交让木低,这与它此时低的补偿点相一致,使得它能更有效地利用弱光来进行光合作用.3个树种的相对叶绿素含量总体上表现为交让木>小叶青冈>中国绣球,且低温反而有利于相对叶绿素含量的提高,这与王致远等[22]的研究结果相一致.
任何环境因子对植物光合作用的影响都可通过叶片叶绿素荧光动力学反映出来[23],植物光合作用受到抑制的最初影响因素就是PSⅡ[24],所以叶绿素荧光分析技术常用来检测植物光合机构对环境胁迫的响应[25-28].本研究结果表明,3种植物的Fv/Fm、Fv′/Fm′和Fv/F0均是随季节呈先下降后上升的变化趋势,通常高等植物Fv/Fm变化极小,胁迫时明显下降[30].本研究中,秋季时中国绣球的Fv/Fm最高,表明其在低温低湿环境中的适应性较强.交让木和小叶青冈在温湿度逐渐降低的11月和12月,Fv/Fm均有所减小,尤其是在温湿度最低的12月降幅最大,这说明二者的光系统受到了不同程度的胁迫,这与张毅龙等[31]研究结果相一致.然而,随着1月份温湿度的升高,交让木Fv/Fm恢复正常;小叶青冈Fv/Fm上升,接近正常范围,说明二者的光系统并未受到不可逆的破坏,且交让木的耐胁迫能力更强.qP和NPQ表征植物对胁迫做出的应对机制.本研究发现,胁迫之下,交让木是以低的qP值来减少光系统的开放比例;小叶青冈则是通过高的NPQ来将过剩的光能以热能的形式耗散掉,从而避免植株被伤害.Yield和ETR均随不同季节温湿度的降低而减小,这与余丽玲等[32-34]的研究结果相一致.
Pn决定着植物物质积累能力的高低,是植物自身生理和环境生态因子综合作用的复杂过程[35-37],其值除与自身因素如叶绿素含量、叶片厚度、叶片成熟程度密切相关外,还和环境因子息息相关[38].本研究结果表明,3个典型树种中秋季交让木的Pn最高,中国绣球次之,小叶青冈的最低;冬季亦是交让木的Pn明显高于小叶青冈,这与郭卫东等[39]的结论相一致.随着时间的变化,交让木和小叶青冈的Pn降低主要是由气孔因素引起的;中国绣球则主要是受到非气孔限制因素控制.由Tr和WUE的变化可知,两乔木能更好地协调碳同化和水分耗散.
本研究结果表明,天目山自然保护区3种典型阔叶林的光合能力均表现为秋季>冬季,交让木>小叶青冈>中国绣球,且在非生长季,Pn依然是正值,说明天目山森林生态系统在碳循环的过程中扮演碳汇的角色.
参考文献
[1] 赵海凤,闫昱霖,张彩虹,等.森林参与碳循环的3种模式:机制与选择[J].林业科学,2014(10):134-139.
[2] 汪森.森林生态系统碳循环研究进展[J].安徽农业科学,2013,41(4):1 560-1 563.
[3] 郑丽波.浅论我国森林碳汇现状及进展[J].内蒙古林业调查设计,2014,37(1):26-28.
[4] 李树朋,夏江宝,王红燕,等.盐胁迫下金银花叶片气体交换参数对光照和温度的响应[J].福建农林大学学报(自然科学版),2014,43(2):132-138.
[5] 偶春,姚侠妹,卓丽环,等.生态因子对地被石竹光合作用的影响[J].福建农林大学学报(自然科学版),2012,41(5):475-480.
[6] 姜楠楠,房义福,吴晓星,等.槭树属3种彩叶植物光合参数和叶绿素荧光参数的日变化[J].福建农林大学学报(自然科学版),2013,42(3):475-480.
[7] 郑宇,何天友,陈凌艳,等.高温胁迫对西洋杜鹃光合作用和叶绿素荧光动力学参数的影响[J].福建农林大学学报(自然科学版),2012,41(6):608-615.
[8] HE Y H, GUO L S, TIAN Y L. Photosynthetic rates and chlorophyll fluorescence of niraria tangutorum at different leaf water potentials[J]. Acta Bot Boreal Occident Sin, 2005,25(11):2 226-2 233.
[9] ZHANG S R. A discussion on chlorophy fluorescence kinetics parameters and their significance[J]. Chinese Bulletin of Botan, 1999,16(4):444-448.
[11] 杨淑贞.天目山兰科植物的分类和区系特点[J].浙江林学院学报,1997,14(4):363-369.
[12] 芮璐.天目山国家级自然保护区植物资源及环境的综合评价[J].科技信息,2008(15):179.
[13] 王敬,韦新良,徐建,等.天目山针阔混交林空间分布格局特征[J].浙江农林大学学报,2014,31(5):668-675.
[14] 秦登,刘鹤,赵明水,等.天目山柳杉叶水势日变化及其与空气温湿度和PAR的相关性[J].植物资源与环境学报,2014,23(1):39-43.
[15] 仇建习,汤孟平,沈利芬,等.天目山近自然毛竹林空间结构与胸径的关系[J].生态学报,2015,35(12):1-13.
[16] 方国景,汤孟平.天目山常绿阔叶林优势种群胸径的空间连续性分析[J].浙江农林大学学报,2014,31(5):663-667.
[17] 刘圣恩,林开敏,石丽娜,等.夹竹桃与珊瑚树光合响应及PSⅡ叶绿素荧光特性比较[J].福建农林大学学报(自然科学版),2015,44(4):384-390.
[18] 许大全.光合速率、光合效率与作物产量[J].生物学通报,1998,34(8):8-9.
[19] 李合生.现代植物生理学[M].北京:高等教育出版社,2002:125.
[20] 丁俊祥,邹杰,唐立松,等.克里雅河流域荒漠——绿洲交错带3种不同生活型植物的光合特性[J].生态学报,2015,35(3):1-13.
[21] 汪赛,伊力塔,余树全,等.模拟酸雨对青冈光合及叶绿素荧光参数的影响[J].应用生态学报,2014,25(8):2 183-2 192.
[22] 王致远,赵广东,王兵,等.丝栗栲、苦槠和青冈幼苗叶片光合生理指标对人工增温和施氮的响应[J].水土保持学报,2014,28(4):293-298.
[23] 殷秀敏,余树全,江洪,等.酸雨胁迫对秃瓣杜英幼苗叶片叶绿素荧光特性和生长的影响[J].应用生态学报,2012,21(6):1 374-1 380.
[24] 管铭,金则新,李月灵,等.千岛湖次生林优势种植物光合生理生态特性[J].生态学报,2015,35(7):1-13.
[25] 李晓,冯伟,曾晓春.叶绿素荧光分析技术及应用进展[J].西北植物学报,2006(10):2 186-2 196.
[26] 温国胜,田海涛,张明如,等.叶绿素荧光分析技术在林木培育中的应用[J].应用生态学报,2006,17(10):1 973-1 977.
[27] 徐德聪,吕芳德,潘晓杰.叶绿素荧光分析技术在果树研究中的应用[J].经济林研究,2003,21(3):89-91.
[28] 殷秀敏,伊力塔,余树全,等.酸雨胁迫对木荷叶片气体交换和叶绿素荧光参数的影响[J].生态环境学报,2010,19(7):1 556-1 562.
[29] 王琰,陈建文,狄晓燕.不同油松种源光合和荧光参数对水分胁迫的响应特征[J].生态学报,2011,31(23):7 031-7 038.
[30] 罗明华,胡进耀,吴庆贵,等.干旱胁迫对丹参叶片气体交换和叶绿素荧光参数的影响[J].应用生态学报,2010,21(3):619-623.
[31] 张毅龙,张卫强,甘先华.低温胁迫对6种珍贵树种苗木光合荧光特性的影响[J].生态环境学报,2014,23(5):777-784.
[32] 余丽玲,陈凌艳,何天友,等.低温胁迫对西洋杜鹃叶片叶绿素荧光参数的影响[J].西南林业大学学报,2013,33(6):1-6.
[33] 刘建,项东云,陈健,等.低温胁迫对桉树光合和叶绿素荧光参数的影响[J].桉树科技,2009,26(1):1-6.
[34] 杨华庚,林位夫.低温胁迫对油棕幼苗光合作用及叶绿素荧光特性的影响[J].中国农学通报,2009,25(24):506-509.
[35] 郑元,陈诗,王大玮,等.滇青冈光合响应曲线研究[J].西南林业大学学报,2013,33(3):25-30.
[36] 管铭,金则新,王强,等.千岛湖次生林优势种植物光合特性对不同光环境的响应[J].应用生态学报,2014,25(3):1 615-1 622.
[37] 张娇,施拥军,朱月清,等.浙北地区常见绿化树种光合固碳特征[J].生态学报,2013,33(6):1 740-1 750.
[38] 吴统贵,虞木奎,孙海菁,等.林药复合系统林下植物光合特性对生长光强的响应[J].中国生态农业学报,2011,19(2):338-341.
[39] 郭卫东,郑建树,张真真,等.短暂低温对佛手光合生理的影响[J].生态学报,2009,29(5):2 286-2 293.
(责任编辑:叶济蓉)
收稿日期:2015-06-02修回日期:2015-09-17
基金项目:国家自然科学基金资助项目(61190114和41171324);高等学校博士学科点专项科研基金资助项目(20110091110028);科技部国家科技基础条件平台资助项目(2005DKA32300);浙江省重点科技创新团队资助项目(2010R50030).
作者简介:马锦丽(1990-),女,硕士.研究方向:植物生理生态.通讯作者江洪(1955-),男,教授,博士生导师.研究方向:全球变化生态学、定量环境遥感和生态系统进化.
中图分类号:Q945.78
文献标识码:A
文章编号:1671-5470(2016)04-0381-10
DOI:10.13323/j.cnki.j.fafu(nat.sci.).2016.04.004
Photo-ecologicalcharacteristicsoftypicalbroad-leavedspeciesinTianmushanNationalNatureReserve
MAJinli1,JIANGHong1,2,SHUHaiyan3,WUMenglin1,FANGChengyuan1,SUNWenwen1,CHENXiaofeng1,NIUXiaodong1,GONGShasha1
(1.InternationalEcologicalResearchCenterofZhejiangForestryUniversity,Hangzhou,Zhejiang311300,China; 2.InternationalEarthSystemScientificInstitute,NanjingUniversity,Nanjing,Jiansu210093,China; 3.MinistryofEducationKeyLaboratoryofEco-environmentsofThreeGorgesReservoirRegion,SchoolofLifeScience,SouthWestUniversity,Chongqing400715,China)
Abstract:To understand the internal mechanism of photosynthesis ability, adaptability and stress resistance of typical broad-leaved species in Tianmushan National Nature Reserve, photosynthesis light response, chlorophyll fluorescence and relative chlorophyll content were measured in autumn and winter. Broad-leaved species studied included Daphniphyllum macropodum, Cyclobalanopsis myrsinaefolia and Hydrangea chinensis Maxim. Results showed that light saturation point (LSP), maximum net photosynthetic rate (Amax), apparent quantum efficiency (AQY), actual photochemical efficiency (Fv′/Fm′), actual quantum yield (Yield) and apparent electron transport rate (ETR) all decreased and then increased in autumn and winter. All these parameters were higher in D. macropodum than the other two species. Light compensation point (LCP) was highest in D. macropodum and lowest in H. chinensis in autumn.Dark respiration rate (Rd) was overally low in D. macropodum, with that of C. myrsinaefolia being the peak in autumn while that of H. chinensis being the peak in winter. Photochemical quenching coefficient (qP) and non photochemical quenching coefficient (NPQ) shared a similar pattern, which qP and NPQ were highest in H. chinensis, and followed by C. myrsinaefolia and then D. macropodum in autumn, while in winter they were higher in Cyclobalanopsis myrsinaefolia than D. macropodum. With regard to stress resistance, three species demonstrated different mechanisms, in which D. macropodum slowed down photosynthesis rate and dark respiration rate while C. myrsinaefolia and H. chinensis transfered excessive light into heat. Relative chlorophyll content was significantly lower in autumn than winter for both D. macropodum and C. myrsinaefolia. At the same point, relative chlorophyll content was highest in D. macropodum and lowest in H. chinensis, with C. myrsinaefolia being the intermediate. Correspondingly, average net photosynthesis (Pn) was higher in D. macropodum than C. myrsinaefolia in both seasons, and in autumn average Pn of H. chinensis was between the other two species, matching the pattern of chlorophyll relative content and temperature and humidity change, which started with a decline and followed by a rise.
Key words:Tianmushan; broad-leaved species; photosynthesis light response; chlorophyll fluorescence; relative chlorophyll content
猜你喜欢
上海文化(文化研究)(2021年2期)2021-05-08 06:08:22
绿色中国(2019年18期)2020-01-04 01:57:08
文化交流(2018年1期)2018-01-09 00:16:12
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
现代农业科技(2016年24期)2017-04-19 13:13:35
山东农业科学(2017年2期)2017-03-15 18:55:09
现代园艺(2016年2期)2016-03-15 16:05:02
江苏农业科学(2015年11期)2016-01-27 14:59:30
江苏农业科学(2015年8期)2015-09-10 16:07:34
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:55
