
TALEN构建Fndc5基因敲除小鼠及初步分析
2016-07-27 00:37:42吴子环胡雄兵3毛凤仪庄峰锋3胡克平孙晓波匡世焕
中国比较医学杂志 2016年6期
关键词:骨骼肌
吴子环,胡雄兵3,毛凤仪,王 超,张 彬,庄峰锋3,胡克平,孙晓波,刘 晓,匡世焕,,金 文
(1.宁波大学海洋学院,浙江宁波 315000;2.中国医学科学院药用植物研究所药理毒理中心中药(天然药物)创新药物研发北京市重点实验室、中国医学科学院-普渡大学脂肪代谢联合实验室,北京 100193;3.北京唯尚立德生物科技有限公司,北京 100085;4.普渡大学动物系,美国)
TALEN构建Fndc5基因敲除小鼠及初步分析
吴子环1,2,胡雄兵3,毛凤仪2,王 超4,张 彬2,庄峰锋3,胡克平2,孙晓波2,刘 晓1,匡世焕2,4,金 文2
(1.宁波大学海洋学院,浙江宁波 315000;2.中国医学科学院药用植物研究所药理毒理中心中药(天然药物)创新药物研发北京市重点实验室、中国医学科学院-普渡大学脂肪代谢联合实验室,北京 100193;3.北京唯尚立德生物科技有限公司,北京 100085;4.普渡大学动物系,美国)
【摘要】目的 通过构建Fndc5基因敲除小鼠,为后续的研究提供动物模型。方法 运用TALEN技术在Fndc5基因FNIII domain中造成缺失突变,并通过测序进行基因型鉴定。通过配对建立稳定遗传系并在mRNA和DNA水平鉴定出生小鼠基因型;对不同年龄段出生小鼠进行体重、血糖分析;通过qPCR确定Fndc5在肾脏、肝脏、大脑、肌肉、心脏等组织中的表达情况。结果 成功构建并鉴定得到4种不同的Fndc5基因敲除小鼠品系;不同年龄段出生小鼠体重、血糖未见显著性差异;确定Fndc5在肌肉、心脏等组织中高表达。结论 本实验在国际上成功构建了Fndc5基因敲除小鼠,并进行了初步分析,为深入研究Fndc5基因在体内中的功能提供了动物模型。
【关键词】骨骼肌;Fndc5基因;纤维连接蛋白;TALEN
骨骼肌在调控系统能量稳态上扮演着重要的角色。而且骨骼作为一个分泌器官渐渐被认同,肌肉分泌和表达的细胞因子和其他多肽类,并且通过自分泌、内分泌、旁分泌的方式在不同组织发挥作用的物质叫“肌因子”[1-3]。其中,肌因子 irisin越来越受到关注,其前体Fndc5(fibronectin type III domain containing protein 5)是一个跨膜蛋白,它的胞外域裂解产生一个溶解的激素irisin[4]。Fndc5最早是在对fibronectin type III domains的基因组搜索中发现的[5]。最初,研究发现Fndc5在运动的刺激下作为PGC1-α依赖的产物,随后裂解产生irisin进入血液作用于白色脂肪,刺激UCP1表达提高产热;促使白色脂肪棕色化,从而提高和改善了肥胖和糖尿病患者的代谢状态,为治愈肥胖和糖尿病提供了可能[2]。随着研究的深入,关于Fndc5的许多其他功能都被证实。
TALEN(transcriptionactivator-like(TAL)effector nuclease)靶向基因敲除基因修饰是第二代新型基因编辑分子生物学工具,首次发现于黄单胞杆菌(Xanthomonas)中[6,7]。TALE中的重复可变双残基(repeat variable di-residue,RVD)[6]与A、C、T、G有恒定的对应关系[8,9]。因此,为识别某一特定氨基酸序列,只需设计相应TALE单元串联克隆,然后在N-端加上核定位信号,并在C-端融合上Fork I核酸内切酶的切割区,就构建成了TALEN[10,11],利用其特异性内切酶活性切割特定的DNA序列,最后细胞可以通过NHEJ(non-homologous end joining)途径修复DNA损伤,在此修复过程中随机插入或删除了一定数目的碱基,造成移码突变,形成了目标基因敲除的突变体[12,13]。目前,TALEN技术已在非洲爪 蟾(XenoPuslaevis)[14,15]、青 鳉(Oryzias latiPes)[16]、斑马鱼(Danio rerio)[17]、大鼠(Rattus norvegicus)[18]、小鼠(Mus musculus)[19]等模式动物的基因敲除研究中获得了成功。
Fndc5作为新肌因子irisin的前体从研究初始其在体内外的功能就备受热议。它在小鼠、人体内的功能研究已经取得了初步进展,但是其在肌肉中的功能研究还未开展。我们通过TALEN技术在国
际上构建Fndc5基因敲除小鼠,并进行了初步分析,为后期研究Fndc5在体内尤其肌肉中的的功能提供了敲除小鼠模型,对于Fndc5/irisin对肌肉本身的影响的认识,将填补这一认知领域的空白。
1 材料和方法
1.1 材料
1.1.1 实验动物:SPF级C57BL/6小鼠品系,购自北京维通利华有限公司,动物生产合格证号:SCXK(京)2014-0001。饲养于中国医学科学院药用植物研究所SPF级动物房,实验动物使用许可证:SYXK(京)2013-0023,动物房温度(20~26)℃,相对湿度40%~70%。
1.1.2 TALEN质粒:TALEN质粒pCS5-eTALEN-T由北京唯尚立德生物科技有限公司合成。靶位点位置 T1:TTTCTAGAAGAAGGATGT gcggatgctccggtt CATTCAGGAGGTGAACA,其中左边TALE识别序列:TTCTAGAAGAAGGATGT 17 bp,右边TALE识别序列:GTTCACCTCCTGAATG 16bp,间隔15 bp。靶 位 点 位 置T3:TCCGGCACCTCAAGGCC aactctgccgtggtcag CTGGGATGTCCTGGAGGA,其中左边TALE识别序列:CCGGCACCTCAAGGCC 16 bp,右边TALE识别序列:CCTCCAGGACATCCCAG 17 bp,间隔17 bp。
1.1.3 引物:对于靶点位置1,设计基因型检测PCR引物:CATGTTTCCTTAGCTCTACTGTGG(正向),GGAGAAAGCATGCATGGCAGTCTC(反向)PCR片段长度为518 bp;对于靶点位置3,设计基因型检测PCR 引 物:GGACCCTTGGTTTGGCCAGTCTA(正 向),CTCTACCATCATCCTCCATGCCTG(反向)PCR片段长度为 479 bp。Real-Time引物:FNDC5-F:ATGAA GGATGGGGAGGAA,FNDC5-R:GCGGCAGAAGAGAG CTATAACA,18s-F:CATGCAGAACC CACGACAGTA,18s-R:CCTCACGCAGCTTGTTG TCTA。
1.1.4 试剂:本实验的引物由北京睿博兴科生物技术有限公司合成。AmpPCRmix(TAKARA,Ro74A)试剂盒、TransZol Up Plus RNA Kit(Tran,ER501-01)、EasyScript First-Strand cDNA SynthesisSuperMxi(Tran,AE301-03)、TransStrt Top Green qPCR SupreMix(Tran,AQ131-02)、Base(碱液)、Neutrlization(中和液)等。
1.2 方法
1.2.1 Fndc5基因敲除TALEN靶点位置的设计:根据Fndc5的基因组结构,Fndc5 protein domain的分析,破坏the FNIII domain,将TALEN的靶点位置设计在外显子3上或者外显子3前,测定SSA活性,选择活性比较高的靶点进行后续的Fndc5基因敲除小鼠的构建。
1.2.2 Fndc5基因敲除鼠的构建流程:将Fndc5-T1和 Fndc5-T3的 TALEN质粒对,体外转录成mRNA后,共显微注射mRNA到小鼠的受精卵中,共注射120个受精卵,出生23只小鼠。两周龄后,剪取4周龄出生小鼠任一只耳朵大约1/3,放置在做好标记的1.5 mL的离心管中,加入75 μL碱液(base)95℃以上40 min;然后加入75 μL中和液(neutrlization),常温静置5 min,溶液即为提取的基因组DNA。以此基因组DNA为模板进行PCR反应,产物测序并根据峰图进行基因型鉴定。
2 结果
2.1 四品系Fndc5基因敲除小鼠的建立
2.1.1 Fndc5基因敲除TALEN靶点位置的设计:根据Fndc5的基因组结构,Fndc5 protein domain的分析,决定破坏the FNIII domain,因此TALEN的靶点位置将设计在外显子3上或者外显子3前,设计结果如图1箭头所示。
2.1.2 Fndc5 TALEN的SSA活性检测:根据SSA活性结果,我们选择活性比较高的靶点Fndc5-T1和Fndc5-T3进行后续的Fndc5基因敲除小鼠的构建,结果见图2。
2.1.3 Fndc5基因敲除首建鼠的鉴定:将受精卵注射后出生23只小鼠,对出生两周的小鼠进行基因型分析和鉴定。结果如图3所示,我们选取Founder 7 (T1一条缺失2 bp,一条缺失4 bp)进行测序图分析。测序峰形图在突变位点开始出现双峰,表明为杂合子。通过峰形分析,可以将突变型1的DNA序列(下指箭头所指峰形)和突变型2的DNA序列(上指箭头所指峰形)分析出来。结果为一条在CCGGTTCATTCAGGA前缺失2个碱基CT;一条在CGGTTCATTCAGG前缺失4个碱基GCTC,均发生移码突变。
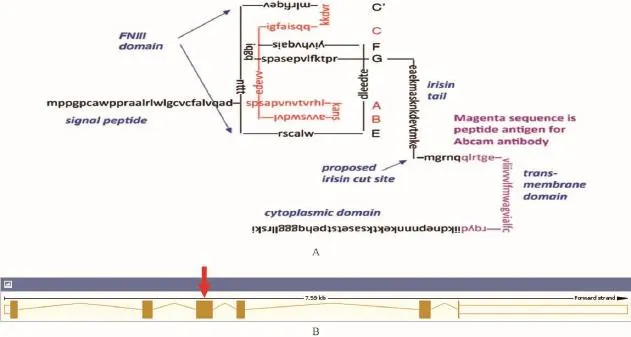
注:A为FNDC5蛋白质的一级结构,B为Fndc5基因组结构(箭头为TALEN的靶点位置)。图1 FNDC5蛋白质一级结构及Fndc5的基因组Note.A is the primary structure of FNDC5 protein;B is the Fndc5 genome structure(red arrow for TALEN target site).Fig.1 The primary structure of FNDC5 protein and the Fndc5 genome.
2.1.2 F1代四品系Fndc5基因敲除鼠的鉴定:根据发生移码突变情况,选取的 Founder 7小鼠、Founder 13小鼠、和Founder 18小鼠分别与野生型C57小鼠交配,出生的小鼠两周龄后按上述方法进行基因型鉴定。通过测序图可分析F1代小鼠的突变情况:如果测序峰全是单峰说明该小鼠是WT(野生型);如果测序图出现双峰则该小鼠为杂合子;如果测序图也是单峰但是在预期位置出现缺失突变则该小鼠为纯合基因敲除小鼠,这在图3中已经进行了类似分析。我们从中选出4种突变类型#7-T1-1(T1缺失2个碱基CT)、#7-T1-2(T1缺失4个碱基GCTC)、#13-T1-1(T1缺失2个碱基TG)、#18-T1-1(T1缺失2个碱基TC)建立稳定遗传F1小鼠品系。四品系F1小鼠的突变位点及突变后导致提前产生终止密码子的位置见表1。基因型中黑体标记部分是TALEN设计敲除的靶位点序列,下划线部分为敲除的基因序列,这些缺失突变都会导致提前产生终止密码子。
2.2 Fndc5敲除小鼠的初步分析
2.2.1 Fndc5小鼠体重分析:通过成功建立和鉴定四品系F1小鼠后,将每个品系之间进行杂交配对,对出生的小鼠进行基因分型鉴定,分别在1月龄、2月龄进行所有出生小鼠的体重测量,体重分析时将雌雄分开,进行纯合子与WT之间的体重差异分析,结果见图4。结果表明小鼠生长正常,在1月龄、2月龄纯合子和WT小鼠之间体重没有统计学差异。
2.2.2 Fndc5在小鼠不同组织的表达情况分析:检测Fndc5在不同组织内的表达情况有助于下一步的体内功能研究。提取小鼠不同组织H(心脏)、K(肾脏)、B(大脑)、L(肝脏)、TA(肌肉)进行液氮研磨提取mRNA,反转录成cDNA进行荧光定量PCR,选取18s作为内参,结果发现Fndc5在H、B、TA中表达量较高,在K、L中表达很少,如图5。
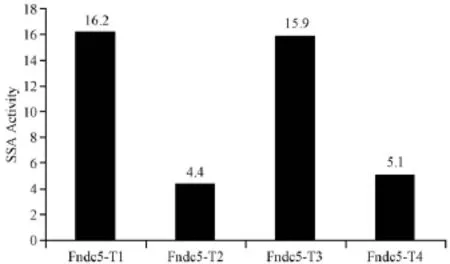
图2 Fndc5 TALEN的SSA活性检测Fig.2 Detection of the Fndc5 TALEN SSA activity
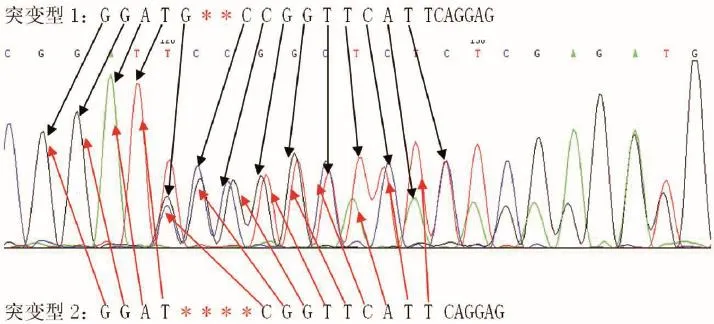
注:突变型1的DNA序列(下指箭头所指峰形)和突变型2的DNA序列(上指箭头所指峰形)。图3 Fndc5基因敲除首建鼠的鉴定Note.The DNA sequence of mutant 1 peak shape(black arrows)and the DNA sequence of mutant 2 peak shape(red arrows).Fig.3 Identification of Fndc5 first built knockout mice

表1 四品系F1小鼠的突变情况Tab.1 The mutation of four different Fndc5-KO lines of mice
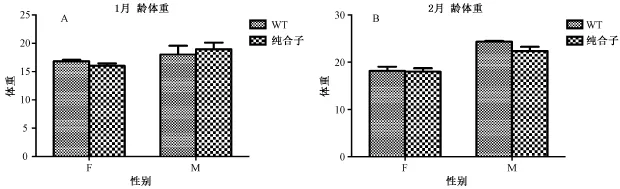
注:A为1月龄体重分析结果,B为2月龄体重分析结果。图4 Fndc5小鼠不同年龄体重分析Note.A:The Results of body weight analysis at one month of age.B:The〛Results of body weight analysis at two months of age.Fig.4 Analysis of body weight of the Fndc5 mice at different ages.
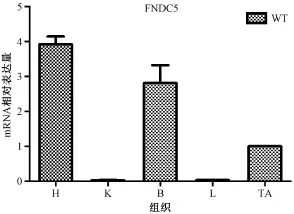
注:H-心脏,K-肾脏,B-大脑,L-肝脏,TA-肌肉图5 Fndc5在小鼠不同组织的表达情况分析。Note.H-heat.K-kidney.B-brain.L-liver.TA-muscleFig.5 Fndc5 expression in different tissues in the mice.
3 讨论
Fndc5作为Irisin(myokine)的前体,作用于白色脂肪提高了全身的产热能力,促进了白色脂肪向棕色脂肪的转化,这一研究从发现伊始就被认为可作为治疗糖尿病的靶点。但是经过几年的研究,争议一直没有平息:Fndc5裂解产生irisin的机制是什么?Irisin在不同组织细胞发挥作用的受体?Irisin在人体内是否发挥积极作用?总而言之关于它在小鼠及人体内的研究还急需广泛开展。
目前关于它在脑、骨髓中的功能研究已经取得了一些进展,但是关于它在肌肉中的研究还未见报道,我们已经成功构建了4种品系Fndc5敲除小鼠,各自的突变类型在上文中也进行了分析,并建立了稳定遗传F1小鼠。我们发现每个品系出生小鼠符合正常的孟德尔比率,生长没有受到影响,在不同年龄段体重没有出现明显差异,我们也分析了Fndc5在不同组织内的表达分析,发现其在H、B、TA中表达量较高,这为我们下一步开展其在肌肉中的功能研究提供了基础。有趣的是,通过Fndc5抗体进行检测敲除小鼠时发现WT和纯合子都出现条带,我们认为本实验中利用TALEN技术构建敲除小鼠模型只是在Fndc5的FNIII domain缺失了几个碱基,导致终止密码子提前产生,从而是蛋白质的功能缺失。但是经过缺失突变的Fndc5还是会产生70-110aa长的蛋白质,关于这部分蛋白质是否能发挥一定的功能还不知道,所以后续可能还需要其他的手段去确切的证实Fndc5敲除小鼠是否成功。
参考文献:
[1] Baskin KK,Winders BR,Olson EN.Muscle as a“mediator”of systemic metabolism [J].Cell Metab.2015,21(2):237 -248.
[2] Bostr öm P,Wu J,Jedrychowski MP,et al.A PGC1-αdependent myokine that drives brown-fat-like development of white fat and thermogenesis[J].Nature 2012,481:463-468.
[3] Pedersen BK,Febbraio MA.Muscles,exercise and obesity:skeletal muscle as a secretory organ[J].Nat Rev Endocrinol. 2012,8(8):457-465.
[4] Erickson HP.Irisin and FNDC5 in retrospect:An exercise hormone or a transmembrane receptor?[J].Adipocyte 2013,2 (4):289-293.
[5] Teufel A,Malik N,Mukhopadhyay M,et al.Frcp1 and Frcp2,two novel fibronectin type III repeat containing genes[J].Gene. 2002,297(1-2):79-83.
[6] Boch J,Bonas U.Xanthomonas AvrBs3 family-type III effectors:discovery and function[J].Annu Rev Phytopathol,2010,48:419-436.
[7] Moscou MJ,Bogdanove AJ.A simple cipher governs DNA recognition by TAL effectors[J].Science,2009,326(5959):1501-1501.
[8] Boch J,Scholze H,Schornack S,et al.Breaking the code of DNA binding specificity of TAL-type III effectors[J].Science,2009,326(5959):1509-1512.
[9] 靳玉珠,王世山,向华,等.TALEN靶向基因修饰新技术研究进展[J].广东农业科学,2013,(19):149-152.
[10] Miller JC,Tan S,Qiao G,et al.A TALE nuclease architecture for efficient genome editing[J].Nat Biotechnol,2010,29(2):143-148.
[11] Smith J,Bibikova M,Whitby FG,et al.Requirements for double-strand cleavage by chimeric restriction enzymes with zinc finger DNA-recognition domains[J].Nucleic Acids Res 2000,28(17):3361-3369.
[12] 汪加兴,张淑君.TALEN和CRISPR/Cas两个基因编辑新技术[J].上海畜牧兽医通讯,2015,(2):22-26.
[13] 倪培凌,刘畅,陈凌懿.DNA剪刀—TALEN和CRISPR/Cas [J].中国细胞生物学学报,2014,36(1):1-7.
[14] Mussolino C,Morbitzer R,Lütge F,et al.A novel TALE nucleasescaffoldenableshighgenomeeditingactivityin combination with low toxicity[J].Nucl Acids Res,2011,39 (21):9283-9293.
[15] Lei Y,Guo X,Liu Y,et al.Efficient targeted gene disruption in Xenopus embryos using engineered transcription activator-like effector nucleases(TALENs)[J].Proc Natl Acad Sci U S A,2012,109(43):17484-17489.
[16] Ansai S,Sakuma T,Yamamoto T,et al.Efficient targeted mutagenesisinmedakausingcustom-designedtranscription activator-like effector nucleases[J].Genetics,2013,193(3):739-749.
[17] Sander JD,Cade L,Khayter C,et al.Targeted gene disruption in somatic zebrafish cells using engineered TALENs[J].Nat Biotechnol,2011,29(8):697-698.
[18] Tesson L,Usal C,Ménoret S,et al.Knockout rats generated by embryo microinjection of TALENs[J].Nat Biotechnol,2011,29 (8):695-696.
[19] Sung YH,Baek IJ,Kim DH,et al.Knockout mice created by TALEN-mediated gene targeting[J].Nat Biotechnol,2013,31 (1):23-24.
〔修回日期〕2016-02-25
【中图分类号】R-33
【文献标识码】A
【文章编号】1671-7856(2016)06-0037-05
doi:10.3969.j.issn.1671-7856.2016.06.008
[基金项目]国家自然科学基金面上项目(81471070);“重大新药创制”科技重大专项(2012ZX09101、2012ZX09301002-001);诺华诺德-协和英才基金(肌信息素抵抗肥胖作用机理的研究);药植所创新团队发展计划资助。
[作者简介]吴子环(1990-),男,硕士研究生,专业:生物功能基因研究,E-mail:wuzihuan1122@163.com。
[通讯作者]金文(1971-),女,副研究员,博士,研究方向:代谢性疾病药物药效学研究,E-mail:wjin@implad.ac.cn。showed no significant differences.Finally high Fndc5 expressions in the muscles,heart and other organs were determined. Conclusions We Have for the first time successfully generated Fndc5 knockout(KO)mouse model using TALEN mediated DNA targeting technique,and performed preliminary analysis.This Fndc5 knockout(KO)mouse model provides a novel tool for further studies on the in vivo function of FNDC5.
Constructtion of a Fndc5 knockout mouse model by TALEN-mediated DNA targeting
WU Zi-huan1,2,HU Xiong-bing3,MAO Feng-yi2,WANG Chao4,ZHANG Bin2,ZHUANG Feng-feng3,HU Ke-ping2,SUN Xiao-bo2,LIU Xiao1,KUANG Shi-huan2,4,JIN Wen2
(1.School of Ocean Biology,Ningbo University,Ningbo 315000,China;2.Beijing Key Laboratory of Innovative Drug Development of Traditional Chinese Medicine(Natural Medicine)and Translational Medicine,CAMS-Purdue Associate Laboratory of Adipose Metabolism,Center of Pharmacology and Toxicology,Institute of Medicinal Plant Development,Peking Union Medical College and Chinese Academy of Medical Sciences,Beijing 100193;3.Beijing Viewsolid Biotech Company Ltd.,Beijing 100085;4.Department of Animal Science,Purdue University,USA)
【Abstract】Objective To construct Fndc5 knockout mouse models and provide animal models for related studies in the future.Methods Indels were introduced into FNIII domain of Fndc5 gene by TALEN technology in mice,and genotypes were identified by sequencing.To set stable genetic system by pairing.Then at mRNA and DNA levels identified the genetype of born mice.At the same time the body weight and blood glucose of the mice at different ages were analyzed. Finally the Fndc5 expression in the kidney,liver,brain,muscles,heart and other organs was determined by qPCR. Results Four different Fndc5-KO lines were generated.The body weight and blood glucose of the mice at different ages
【Key words】Skeletal muscle;Fndc5;Fibronectin;TALEN;Knockout mouse model
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23 13:46:58
临床军医杂志(2021年9期)2021-09-18 02:44:16
天然产物研究与开发(2018年2期)2018-04-04 02:01:18
潍坊学院学报(2016年2期)2016-12-01 12:59:45
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国组织化学与细胞化学杂志(2016年3期)2016-02-27 11:15:37
医学研究杂志(2015年5期)2015-06-10 06:43:26
军事体育学报(2015年2期)2015-02-27 16:02:00
实用老年医学(2013年7期)2013-03-11 18:39:07
