
缺铁及不同铁源对水培大白菜生物量、光合参数和矿质元素含量的影响
2016-07-25 01:10颜廷帅姜振升侯文通刘振香贾亮
江苏农业科学 2016年6期
颜廷帅++姜振升++侯文通++刘振香++贾亮


摘要:以大白菜(Brassica pekinensis L.)为试材,研究缺铁处理和硫酸亚铁、柠檬酸亚铁、柠檬酸铁、EDTA-Fe这4种相同铁元素浓度铁源对水培大白菜生物量、光合参数和氮、磷、钾、钙、镁、铁元素含量的影响。结果表明,与缺铁处理相比,加入不同铁源可显著提高大白菜植株的干质量、净光合速率、气孔导度,细胞间CO2浓度显著降低;不同铁源处理时,大白菜老叶中的全氮含量显著升高,镁含量显著降低;硫酸亚铁、柠檬酸铁处理时,大白菜根中的铁元素含量升高,柠檬酸亚铁处理时,大白菜根和新叶中的铁元素含量升高;柠檬酸亚铁处理时,大白菜新叶和老叶的钙含量、新叶镁含量显著低于缺铁处理和其他铁源处理;不同铁源处理时,大白菜根中的磷含量相对最高,而缺铁处理时大白菜新叶中的磷含量相对最高。
关键词:缺铁;大白菜;生物量;光合参数;矿质元素;柠檬酸亚铁
中图分类号: S634.106文献标志码: A文章编号:1002-1302(2016)06-0275-03
收稿日期:2016-01-14
基金项目:山东省科技重大专项(新兴产业)(编号:2015ZDXX0502B02)。
作者简介:颜廷帅(1989—),男,山东临沂人,主要从事植物营养与新型肥料研究。E-mail:yanaiyanzhi@163.com。
通信作者:姜振升,硕士,农艺师,主要从事植物营养与新型肥料研究。E-mail:jzssdau@163.com。铁是植物必需的微量元素,参与植物光合作用和叶绿素合成,在植物氧化还原反应和电子传递过程中具有重要的作用[1]。铁在土壤中虽然含量丰富,但植物体缺铁的现象仍然比较普遍,这是由于土壤中有效铁的含量很低,尤其在碱性土壤中,有效铁的浓度非常低[2]。据统计,世界上约1/3的土壤为碱性土壤。因此,土壤有效铁含量不足成为作物生长发育最大的限制因子之一[3]。铁肥的施用可以改善土壤的缺铁状况。最早的铁肥以无机铁源 FeSO4·7H2O 和FeCl3为主[4],后以有机态铁为铁源的营养液配方得到应用,但有机态铁性质不稳定在一定程度上限定了其应用[5],对于以何种铁源作为铁肥仍存在争议。本试验以大白菜(Brassica pekinensis L.)为供试材料,采用水培方式,设置缺铁和不同铁源处理,探究缺铁和不同铁源处理对大白菜生长、光合参数和矿质元素含量的影响,为铁肥的合理施用提供参考。
1材料与方法
1.1供试材料
3~4张叶片的大白菜幼苗,由营养钵催芽生长。供试水为蒸馏水,铁含量小于0.000 1 mg/L。
1.2方法及处理
试验于2015年9月5日开始,采用水培法。选取长势一致的15株大白菜幼苗,转入1/2 Hoagland和Arnon全浓度溶液[6]进行培养,每天通气2 h;待大白菜幼苗生长7 d,将大白菜幼苗随机分为5组,每组3株,分别移入缺铁、含硫酸亚铁(Fe2+)、柠檬酸亚铁(Fe2+)、EDTA-Fe(Fe3+、Fe2+混合物)、柠檬酸铁(Fe3+)的Hoagland和Arnon(全浓度)溶液中继续培养18 d,每天通气2 h,每7 d更换1次营养液,不同铁源处理的铁元素浓度均为2.8 mg/L;取样测定。
1.3测定项目与方法
1.3.1光合参数大白菜收获前,用英国PP Systems公司生产的CIRAS-2便携式光合作用测定仪,于09:00—11:00采用固定光强800 μmol/(m2·s)测定每株大白菜功能叶片相同叶位的细胞间隙CO2浓度(Ci)、蒸腾速率(E)、气孔导度(Gs)和净光合速率(Pn)等光合相关参数。
1.3.2干质量将大白菜分为根、老叶、新叶3个部分,放于信封中置于干燥箱105 ℃杀青30 min,75 ℃烘干至恒质量,称量根系和地上部的干质量。样品备存,用于测定氮、磷、钾、钙、镁、铁元素含量。
1.3.3主要元素含量测定将烘干的大白菜干样磨碎,准确称取0.200 g,用H2SO4-H2O2 联合消煮法煮至澄清,氮、磷、钾含量分别采用凯氏定氮法、钒钼黄比色法、火焰光度法测定[7],钙、镁、铁含量采用ICP-AES法测定[8]。
1.4 数据处理
试验数据采用 Microsoft Excel 2007软件进行处理,采用SPSS 16.0软件进行统计分析和差异显著性检验。
2结果与分析
2.1缺铁和不同铁源处理对水培大白菜生物量的影响
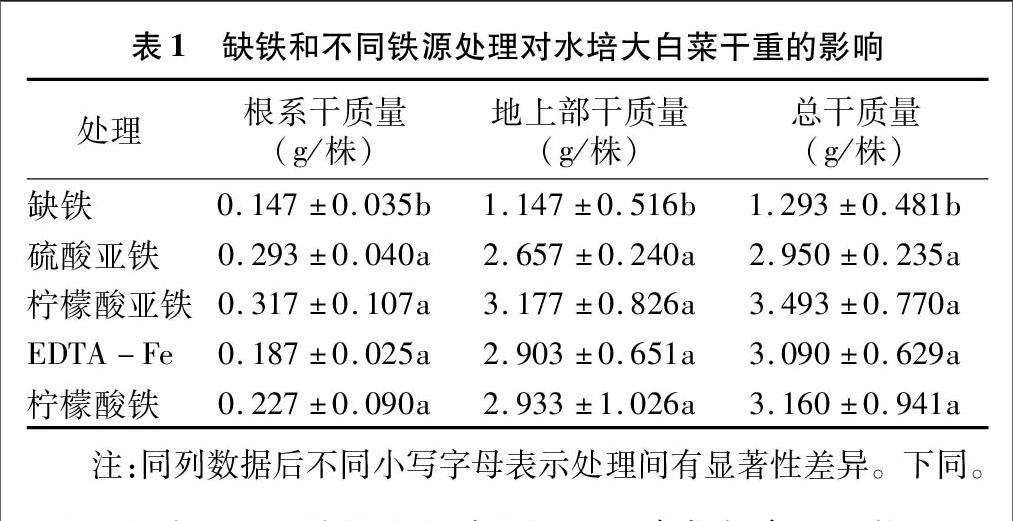
由表1可见,与缺铁处理相比,加入不同铁肥时,大白菜根系和地上部干质量均显著增加,不同铁源处理之间大白菜根系和地上部干质量无显著性差异;柠檬酸亚铁处理时,大白菜根系和地上部的干质量相对最大,较缺铁处理分别增加115.65%、176.98%。
2.2缺铁和不同铁源处理对水培大白菜光合作用的影响
由表2可见,与缺铁处理相比,不同铁源处理大白菜的净光合速率显著升高,其中,EDTA-Fe处理的大白菜净光合速率相对最高,较缺铁处理增加213.38%,与其他铁源处理的大白菜净光合速率无显著性差异;缺铁处理时,大白菜功能叶片胞间CO2浓度显著高于其他处理,这可能是由于缺铁导致光合电子传递受阻,进而抑制光合作用,造成CO2无法被吸收利用,使CO2在细胞间隙累积;不同铁源处理时大白菜的气孔导度均显著升高,其中,EDTA-Fe处理气孔导度相对最大,较缺铁处理增加了245.65%;大白菜细胞间隙CO2浓度较缺铁处理显著降低;柠檬酸亚铁处理时,大白菜的蒸腾速率相对最高,为3.10 mmol/(m2·s)。另外,试验还表明,气孔导度值与净光合速率呈正相关,这与Wong等的研究结果[9]相同。
2.3缺铁和不同铁源处理对水培大白菜各组织元素含量和分布的影响
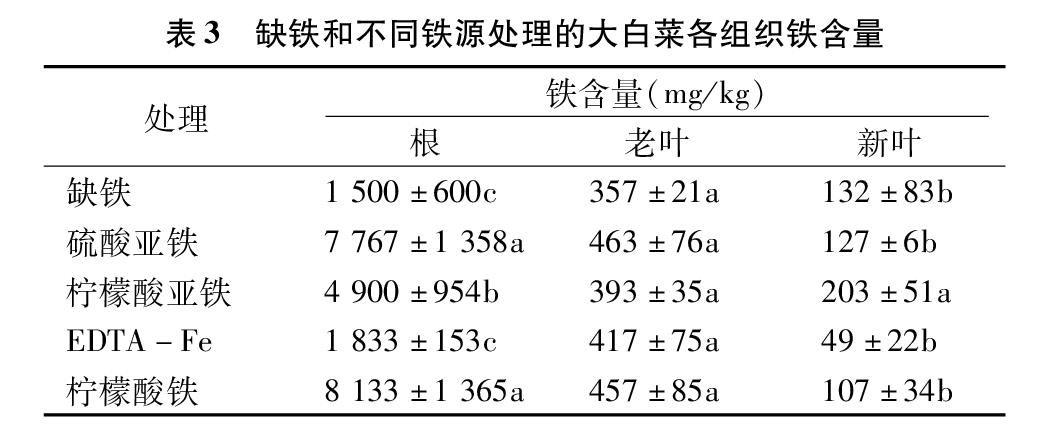
2.3.1铁元素由表3可知,各处理大白菜,根中的铁含量相对最高,老叶次之,新叶最低;在根中,硫酸亚铁、柠檬酸亚铁、柠檬酸铁处理的铁含量较缺铁处理有显著上升,分别上升417.80%、226.67%、442.20%;在老叶中,各处理的铁含量无显著性差异;在新叶中,柠檬酸亚铁处理的铁含量显著高于其他处理,较缺铁处理增加了53.79%。这说明硫酸亚铁、柠檬酸亚铁、柠檬酸铁促进了大白菜根系对铁的吸收,同时柠檬酸亚铁促进了铁向大白菜新叶运输;使用EDTA-Fe对大白菜根系铁的吸收和向地上部的转运无明显效果。
2.3.2氮、磷和钾元素由表4可知,不同铁源处理的大白菜全氮含量以新叶最高,老叶次之,根相对最低,缺铁处理大白菜根的氮含量高于老叶;在根中,硫酸亚铁处理的氮含量高于其他处理;在老叶中,不同铁源处理的氮含量均显著高于缺铁处理,其中硫酸亚铁处理最高;在新叶中,柠檬酸亚铁处理的氮含量较其他处理有显著降低。由于氮是易移动的元素,缺氮情况下老叶中的氮会向新叶转移,导致老叶氮含量降低,缺铁抑制了大白菜对氮的吸收。
由表5可见,不同铁源处理的大白菜磷含量以根最高,新叶次之,老叶最低,缺铁处理的大白菜新叶中磷含量高于根;在根中,柠檬酸铁处理的磷含量高于其他处理;在新叶中,柠檬酸亚铁处理较缺铁处理的磷含量有显著降低;在老叶中,各处理的磷含量无显著性差异;缺铁情况下,新叶中磷含量最高,这是由于在缺铁情况下,磷向大白菜新叶的运输增加,对磷的吸收也增加。
由表6可知,各处理大白菜的钾含量以老叶最高,新叶次之,根最低;在根中,硫酸亚铁、柠檬酸铁处理的钾含量显著高于其他处理;在老叶中,各处理的钾含量无显著性差异;在新叶中,硫酸亚铁处理的钾含量相对最高,柠檬酸亚铁处理的钾含量最低。
2.3.3钙、镁元素由表7可知,各处理大白菜的钙含量以老叶最高;在根中,各处理的钙含量无显著性差异;在老叶、新叶中,柠檬酸亚铁处理的钙含量显著低于其他处理,柠檬酸亚铁抑制了钙向地上部的运输和分配。
由表8可知,各处理大白菜的镁含量以老叶最高,新叶次之,根最低;在根中,硫酸亚铁、柠檬酸铁处理的镁含量较其他处理有显著上升;在老叶中,不同铁源处理的镁含量较缺铁处理有显著降低;在新叶中,柠檬酸亚铁处理的镁含量较其他处理有显著降低,柠檬酸亚铁抑制了镁向地上部的运输和分配。
3结论与讨论
铁的缺乏影响植物体的生长和代谢,缺铁植物叶绿体片层结构发生变化,严重时叶绿体解体,光合电子传递链中断,净光合速率下降,同化产物的形成受阻[10]。本试验缺铁大白菜表现出典型的缺铁黄化现象,净光合速率和生物量显著降低,与前人的研究结论[10]一致。
氮、磷、钾被称为肥料三要素,植物对氮、磷、钾的需要量较多,而土壤中含量较少,需要大量补充,而不同铁源的加入对氮、磷、钾在植物体内的含量和分布产生影响。氮是植物生长发育需求量最大的营养元素,既是构成植物有机体的结构物质,也是植物生理代谢过程中起催化作用的物质[11]。试验结果表明,缺铁处理大白菜老叶中的氮含量显著低于铁处理,这是由于缺铁抑制了大白菜对氮的吸收,而氮是可以重复利用的元素,氮含量不足情况下大白菜老叶中的氮会向新叶转移,从而造成老叶中氮含量显著降低。有研究表明,植物体对铁、磷元素的吸收存在拮抗效应[12]。本试验中,缺铁处理大白菜新叶中的磷含量相对最高,这表明在缺铁情况下磷向新叶的运输增加,大白菜对磷的吸收增加。钾元素参与植物生长发育中许多重要的生理生化过程,在维持细胞膨压、调节酶的活性等过程中具有重要的作用,并且促进了光合作用的进程和光合产物的运输[13]。本试验中,硫酸亚铁和柠檬酸铁处理的大白菜根中钾含量显著高于其他处理,表明加入这2种铁肥促进了大白菜对钾的吸收;而柠檬酸亚铁处理的大白菜新叶中钾含量显著低于缺铁处理,表明加入柠檬酸亚铁可能抑制了钾元素向地上部的转运。
镁和钙是植物所必需的中量元素,镁在光合作用、酶的活化和蛋白质合成等进程中具有重要作用[14]。钙能稳定细胞壁和细胞膜的结构,存在于液泡中的钙具有渗透调节作用,细胞的正常有丝分裂需要少量钙的参与,钙作为第二信使在细胞转导中具有重要作用[15-16]。钙含量不足容易引起大白菜干烧心[17]。缺铁情况下,大白菜老叶中的镁含量有显著下降,可能是由于缺铁情况下叶绿素合成受阻,光合速率下降[18]。缺铁情况下,柠檬酸亚铁处理老叶和新叶中的镁和钙含量有显著降低,这说明加入柠檬酸亚铁可能抑制了大白菜对钙和镁向地上部的转运。
总之,加入不同铁源,显著增加了大白菜的生物量和光合效率,促进了大白菜对氮的吸收,抑制了大白菜对磷的吸收;柠檬酸亚铁处理大白菜使其对铁的吸收效率相对最高,但抑制了对钾、钙、镁向地上部的转运;与其他铁源处理大白菜相比,EDTA-Fe处理的大白菜对铁的吸收相对最弱。硫酸亚铁、柠檬酸铁处理对大白菜的生长、生物量和元素含量均有正向影响,然而考虑到土壤施肥过程中,硫酸亚铁遇空气易氧化,造成土壤中铁的有效浓度降低,因此,综合考虑,大白菜以柠檬酸铁作为铁肥相对较好。
参考文献:
[1]吴慧兰,王宁,凌宏清. 植物铁吸收、转运和调控的分子机制研究进展[J]. 植物学通报,2007,24(6):779-788.
[2]Guerinot M L,Yi Y. Iron:nutritious,noxious,and not readily available[J]. Plant Physiology,1994,104(3):815-820.
[3]Lindsay W L,Schwab A P. The chemistry of iron in soils and its availability to plants[J]. Journal of Plant Nutrition,1982,5(4/5/6/7):821-840.
[4]Chaney R L,Muns J B,Cathey H M. Effectiveness of digested sewage sludge compost in supplying nutrients for soilless potting media[J]. J Am Soc Hort Sci,1980,105:485-489.
[5]De Bootdt M,Veronck O. The physical properties of the substratesinhorticulture[J]. Acta Horticulture,1972(26):37-44.
[6]Hoagland D R,Arnon D I. The water-culture method for growing plants without soil[M]. California Agricultural Experiment Station,1950.
[7]鲍士旦. 土壤农化分析[M]. 北京:中国农业出版社,2000.
[8]Anderson K A. Micro-digestion and ICP-AES analysis for the determination of macro and micro elements in plant tissues[J]. Atomic Spectroscopy,1996,17(1):30-33.
[9]Wong S C,Cowan I R,Farquhar G D. Leaf conductance in relation to rate of CO2 assimilation:Ⅰ. Influence of nitrogen nutrition,phosphorus nutrition,photon flux density,and ambient partial pressure of CO2 during ontogeny[J]. Plant Physiology,1985,78(4):821-825.
[10]金亚波,韦建玉,王军. 植物铁营养研究进展Ⅰ:生理生化[J]. 安徽农业科学,2007,35(32):10215-10219.
[11]郝建军,于洋,张婷. 植物生理学[M]. 北京:化学工业出版社,2013.
[12]郑录庆. 水稻铁磷互作的分子机理及富铁转基因水稻研究[D]. 杭州:浙江大学生命科学学院,2009.
[13]胡笃敬,董任瑞,葛旦之. 植物钾营养的理论与实践[M]. 长沙:湖南科学技术出版社,1993:58-109.
[14]汪洪,褚天铎. 植物镁素营养的研究进展[J]. 植物学通报,1999,16(3):245.
[15]Marschner H,Rimmington G. Mineral nutrition of higher plants[J]. Plant Cell and Environment,1988,11:147-148.
[16]Hepler P K. Calcium:a central regulator of plant growth and development[J]. The Plant Cell,2005,17(8):2142-2155.
[17]Mynard D N,Gersten B,Vernell H F. The distribution of calcium as related to internal tip-burn variety and calcium nutrition in cabbage[J]. Pro Amer Soc Hort Sci,1965,86:392-396.
[18]Spiller S,Terry N. Limiting factors in photosynthesis:Ⅱ. Iron stress diminishes photochemical capacity by reducing the number of photosynthetic units[J]. Plant Physiology,1980,65(1):121-125.
颜廷帅 姜振升 侯文通 刘振香 贾亮


摘要:以大白菜(Brassica pekinensis L.)为试材,研究缺铁处理和硫酸亚铁、柠檬酸亚铁、柠檬酸铁、EDTA-Fe这4种相同铁元素浓度铁源对水培大白菜生物量、光合参数和氮、磷、钾、钙、镁、铁元素含量的影响。结果表明,与缺铁处理相比,加入不同铁源可显著提高大白菜植株的干质量、净光合速率、气孔导度,细胞间CO2浓度显著降低;不同铁源处理时,大白菜老叶中的全氮含量显著升高,镁含量显著降低;硫酸亚铁、柠檬酸铁处理时,大白菜根中的铁元素含量升高,柠檬酸亚铁处理时,大白菜根和新叶中的铁元素含量升高;柠檬酸亚铁处理时,大白菜新叶和老叶的钙含量、新叶镁含量显著低于缺铁处理和其他铁源处理;不同铁源处理时,大白菜根中的磷含量相对最高,而缺铁处理时大白菜新叶中的磷含量相对最高。
关键词:缺铁;大白菜;生物量;光合参数;矿质元素;柠檬酸亚铁
中图分类号: S634.106文献标志码: A文章编号:1002-1302(2016)06-0275-03
收稿日期:2016-01-14
基金项目:山东省科技重大专项(新兴产业)(编号:2015ZDXX0502B02)。
作者简介:颜廷帅(1989—),男,山东临沂人,主要从事植物营养与新型肥料研究。E-mail:yanaiyanzhi@163.com。
通信作者:姜振升,硕士,农艺师,主要从事植物营养与新型肥料研究。E-mail:jzssdau@163.com。铁是植物必需的微量元素,参与植物光合作用和叶绿素合成,在植物氧化还原反应和电子传递过程中具有重要的作用[1]。铁在土壤中虽然含量丰富,但植物体缺铁的现象仍然比较普遍,这是由于土壤中有效铁的含量很低,尤其在碱性土壤中,有效铁的浓度非常低[2]。据统计,世界上约1/3的土壤为碱性土壤。因此,土壤有效铁含量不足成为作物生长发育最大的限制因子之一[3]。铁肥的施用可以改善土壤的缺铁状况。最早的铁肥以无机铁源 FeSO4·7H2O 和FeCl3为主[4],后以有机态铁为铁源的营养液配方得到应用,但有机态铁性质不稳定在一定程度上限定了其应用[5],对于以何种铁源作为铁肥仍存在争议。本试验以大白菜(Brassica pekinensis L.)为供试材料,采用水培方式,设置缺铁和不同铁源处理,探究缺铁和不同铁源处理对大白菜生长、光合参数和矿质元素含量的影响,为铁肥的合理施用提供参考。
1材料与方法
1.1供试材料
3~4张叶片的大白菜幼苗,由营养钵催芽生长。供试水为蒸馏水,铁含量小于0.000 1 mg/L。
1.2方法及处理
试验于2015年9月5日开始,采用水培法。选取长势一致的15株大白菜幼苗,转入1/2 Hoagland和Arnon全浓度溶液[6]进行培养,每天通气2 h;待大白菜幼苗生长7 d,将大白菜幼苗随机分为5组,每组3株,分别移入缺铁、含硫酸亚铁(Fe2+)、柠檬酸亚铁(Fe2+)、EDTA-Fe(Fe3+、Fe2+混合物)、柠檬酸铁(Fe3+)的Hoagland和Arnon(全浓度)溶液中继续培养18 d,每天通气2 h,每7 d更换1次营养液,不同铁源处理的铁元素浓度均为2.8 mg/L;取样测定。
1.3测定项目与方法
1.3.1光合参数大白菜收获前,用英国PP Systems公司生产的CIRAS-2便携式光合作用测定仪,于09:00—11:00采用固定光强800 μmol/(m2·s)测定每株大白菜功能叶片相同叶位的细胞间隙CO2浓度(Ci)、蒸腾速率(E)、气孔导度(Gs)和净光合速率(Pn)等光合相关参数。
1.3.2干质量将大白菜分为根、老叶、新叶3个部分,放于信封中置于干燥箱105 ℃杀青30 min,75 ℃烘干至恒质量,称量根系和地上部的干质量。样品备存,用于测定氮、磷、钾、钙、镁、铁元素含量。
1.3.3主要元素含量测定将烘干的大白菜干样磨碎,准确称取0.200 g,用H2SO4-H2O2 联合消煮法煮至澄清,氮、磷、钾含量分别采用凯氏定氮法、钒钼黄比色法、火焰光度法测定[7],钙、镁、铁含量采用ICP-AES法测定[8]。
1.4 数据处理
试验数据采用 Microsoft Excel 2007软件进行处理,采用SPSS 16.0软件进行统计分析和差异显著性检验。
2结果与分析
2.1缺铁和不同铁源处理对水培大白菜生物量的影响
由表1可见,与缺铁处理相比,加入不同铁肥时,大白菜根系和地上部干质量均显著增加,不同铁源处理之间大白菜根系和地上部干质量无显著性差异;柠檬酸亚铁处理时,大白菜根系和地上部的干质量相对最大,较缺铁处理分别增加115.65%、176.98%。
2.2缺铁和不同铁源处理对水培大白菜光合作用的影响
由表2可见,与缺铁处理相比,不同铁源处理大白菜的净光合速率显著升高,其中,EDTA-Fe处理的大白菜净光合速率相对最高,较缺铁处理增加213.38%,与其他铁源处理的大白菜净光合速率无显著性差异;缺铁处理时,大白菜功能叶片胞间CO2浓度显著高于其他处理,这可能是由于缺铁导致光合电子传递受阻,进而抑制光合作用,造成CO2无法被吸收利用,使CO2在细胞间隙累积;不同铁源处理时大白菜的气孔导度均显著升高,其中,EDTA-Fe处理气孔导度相对最大,较缺铁处理增加了245.65%;大白菜细胞间隙CO2浓度较缺铁处理显著降低;柠檬酸亚铁处理时,大白菜的蒸腾速率相对最高,为3.10 mmol/(m2·s)。另外,试验还表明,气孔导度值与净光合速率呈正相关,这与Wong等的研究结果[9]相同。
2.3缺铁和不同铁源处理对水培大白菜各组织元素含量和分布的影响
2.3.1铁元素由表3可知,各处理大白菜,根中的铁含量相对最高,老叶次之,新叶最低;在根中,硫酸亚铁、柠檬酸亚铁、柠檬酸铁处理的铁含量较缺铁处理有显著上升,分别上升417.80%、226.67%、442.20%;在老叶中,各处理的铁含量无显著性差异;在新叶中,柠檬酸亚铁处理的铁含量显著高于其他处理,较缺铁处理增加了53.79%。这说明硫酸亚铁、柠檬酸亚铁、柠檬酸铁促进了大白菜根系对铁的吸收,同时柠檬酸亚铁促进了铁向大白菜新叶运输;使用EDTA-Fe对大白菜根系铁的吸收和向地上部的转运无明显效果。
2.3.2氮、磷和钾元素由表4可知,不同铁源处理的大白菜全氮含量以新叶最高,老叶次之,根相对最低,缺铁处理大白菜根的氮含量高于老叶;在根中,硫酸亚铁处理的氮含量高于其他处理;在老叶中,不同铁源处理的氮含量均显著高于缺铁处理,其中硫酸亚铁处理最高;在新叶中,柠檬酸亚铁处理的氮含量较其他处理有显著降低。由于氮是易移动的元素,缺氮情况下老叶中的氮会向新叶转移,导致老叶氮含量降低,缺铁抑制了大白菜对氮的吸收。
由表5可见,不同铁源处理的大白菜磷含量以根最高,新叶次之,老叶最低,缺铁处理的大白菜新叶中磷含量高于根;在根中,柠檬酸铁处理的磷含量高于其他处理;在新叶中,柠檬酸亚铁处理较缺铁处理的磷含量有显著降低;在老叶中,各处理的磷含量无显著性差异;缺铁情况下,新叶中磷含量最高,这是由于在缺铁情况下,磷向大白菜新叶的运输增加,对磷的吸收也增加。
由表6可知,各处理大白菜的钾含量以老叶最高,新叶次之,根最低;在根中,硫酸亚铁、柠檬酸铁处理的钾含量显著高于其他处理;在老叶中,各处理的钾含量无显著性差异;在新叶中,硫酸亚铁处理的钾含量相对最高,柠檬酸亚铁处理的钾含量最低。
2.3.3钙、镁元素由表7可知,各处理大白菜的钙含量以老叶最高;在根中,各处理的钙含量无显著性差异;在老叶、新叶中,柠檬酸亚铁处理的钙含量显著低于其他处理,柠檬酸亚铁抑制了钙向地上部的运输和分配。
由表8可知,各处理大白菜的镁含量以老叶最高,新叶次之,根最低;在根中,硫酸亚铁、柠檬酸铁处理的镁含量较其他处理有显著上升;在老叶中,不同铁源处理的镁含量较缺铁处理有显著降低;在新叶中,柠檬酸亚铁处理的镁含量较其他处理有显著降低,柠檬酸亚铁抑制了镁向地上部的运输和分配。
3结论与讨论
铁的缺乏影响植物体的生长和代谢,缺铁植物叶绿体片层结构发生变化,严重时叶绿体解体,光合电子传递链中断,净光合速率下降,同化产物的形成受阻[10]。本试验缺铁大白菜表现出典型的缺铁黄化现象,净光合速率和生物量显著降低,与前人的研究结论[10]一致。
氮、磷、钾被称为肥料三要素,植物对氮、磷、钾的需要量较多,而土壤中含量较少,需要大量补充,而不同铁源的加入对氮、磷、钾在植物体内的含量和分布产生影响。氮是植物生长发育需求量最大的营养元素,既是构成植物有机体的结构物质,也是植物生理代谢过程中起催化作用的物质[11]。试验结果表明,缺铁处理大白菜老叶中的氮含量显著低于铁处理,这是由于缺铁抑制了大白菜对氮的吸收,而氮是可以重复利用的元素,氮含量不足情况下大白菜老叶中的氮会向新叶转移,从而造成老叶中氮含量显著降低。有研究表明,植物体对铁、磷元素的吸收存在拮抗效应[12]。本试验中,缺铁处理大白菜新叶中的磷含量相对最高,这表明在缺铁情况下磷向新叶的运输增加,大白菜对磷的吸收增加。钾元素参与植物生长发育中许多重要的生理生化过程,在维持细胞膨压、调节酶的活性等过程中具有重要的作用,并且促进了光合作用的进程和光合产物的运输[13]。本试验中,硫酸亚铁和柠檬酸铁处理的大白菜根中钾含量显著高于其他处理,表明加入这2种铁肥促进了大白菜对钾的吸收;而柠檬酸亚铁处理的大白菜新叶中钾含量显著低于缺铁处理,表明加入柠檬酸亚铁可能抑制了钾元素向地上部的转运。
镁和钙是植物所必需的中量元素,镁在光合作用、酶的活化和蛋白质合成等进程中具有重要作用[14]。钙能稳定细胞壁和细胞膜的结构,存在于液泡中的钙具有渗透调节作用,细胞的正常有丝分裂需要少量钙的参与,钙作为第二信使在细胞转导中具有重要作用[15-16]。钙含量不足容易引起大白菜干烧心[17]。缺铁情况下,大白菜老叶中的镁含量有显著下降,可能是由于缺铁情况下叶绿素合成受阻,光合速率下降[18]。缺铁情况下,柠檬酸亚铁处理老叶和新叶中的镁和钙含量有显著降低,这说明加入柠檬酸亚铁可能抑制了大白菜对钙和镁向地上部的转运。
总之,加入不同铁源,显著增加了大白菜的生物量和光合效率,促进了大白菜对氮的吸收,抑制了大白菜对磷的吸收;柠檬酸亚铁处理大白菜使其对铁的吸收效率相对最高,但抑制了对钾、钙、镁向地上部的转运;与其他铁源处理大白菜相比,EDTA-Fe处理的大白菜对铁的吸收相对最弱。硫酸亚铁、柠檬酸铁处理对大白菜的生长、生物量和元素含量均有正向影响,然而考虑到土壤施肥过程中,硫酸亚铁遇空气易氧化,造成土壤中铁的有效浓度降低,因此,综合考虑,大白菜以柠檬酸铁作为铁肥相对较好。
参考文献:
[1]吴慧兰,王宁,凌宏清. 植物铁吸收、转运和调控的分子机制研究进展[J]. 植物学通报,2007,24(6):779-788.
[2]Guerinot M L,Yi Y. Iron:nutritious,noxious,and not readily available[J]. Plant Physiology,1994,104(3):815-820.
[3]Lindsay W L,Schwab A P. The chemistry of iron in soils and its availability to plants[J]. Journal of Plant Nutrition,1982,5(4/5/6/7):821-840.
[4]Chaney R L,Muns J B,Cathey H M. Effectiveness of digested sewage sludge compost in supplying nutrients for soilless potting media[J]. J Am Soc Hort Sci,1980,105:485-489.
[5]De Bootdt M,Veronck O. The physical properties of the substratesinhorticulture[J]. Acta Horticulture,1972(26):37-44.
[6]Hoagland D R,Arnon D I. The water-culture method for growing plants without soil[M]. California Agricultural Experiment Station,1950.
[7]鲍士旦. 土壤农化分析[M]. 北京:中国农业出版社,2000.
[8]Anderson K A. Micro-digestion and ICP-AES analysis for the determination of macro and micro elements in plant tissues[J]. Atomic Spectroscopy,1996,17(1):30-33.
[9]Wong S C,Cowan I R,Farquhar G D. Leaf conductance in relation to rate of CO2 assimilation:Ⅰ. Influence of nitrogen nutrition,phosphorus nutrition,photon flux density,and ambient partial pressure of CO2 during ontogeny[J]. Plant Physiology,1985,78(4):821-825.
[10]金亚波,韦建玉,王军. 植物铁营养研究进展Ⅰ:生理生化[J]. 安徽农业科学,2007,35(32):10215-10219.
[11]郝建军,于洋,张婷. 植物生理学[M]. 北京:化学工业出版社,2013.
[12]郑录庆. 水稻铁磷互作的分子机理及富铁转基因水稻研究[D]. 杭州:浙江大学生命科学学院,2009.
[13]胡笃敬,董任瑞,葛旦之. 植物钾营养的理论与实践[M]. 长沙:湖南科学技术出版社,1993:58-109.
[14]汪洪,褚天铎. 植物镁素营养的研究进展[J]. 植物学通报,1999,16(3):245.
[15]Marschner H,Rimmington G. Mineral nutrition of higher plants[J]. Plant Cell and Environment,1988,11:147-148.
[16]Hepler P K. Calcium:a central regulator of plant growth and development[J]. The Plant Cell,2005,17(8):2142-2155.
[17]Mynard D N,Gersten B,Vernell H F. The distribution of calcium as related to internal tip-burn variety and calcium nutrition in cabbage[J]. Pro Amer Soc Hort Sci,1965,86:392-396.
[18]Spiller S,Terry N. Limiting factors in photosynthesis:Ⅱ. Iron stress diminishes photochemical capacity by reducing the number of photosynthetic units[J]. Plant Physiology,1980,65(1):121-125.
猜你喜欢
中国-东盟博览(政经版)(2023年1期)2023-03-15
中国果树(2020年2期)2020-07-25
中国果业信息(2017年4期)2017-05-09
热带农业科学(2016年10期)2016-12-12
蔬菜(2016年8期)2016-10-10
新作文(小学中高年级版)(2015年5期)2015-04-12
植物营养与肥料学报(2011年3期)2011-10-24
