
穿心莲和水飞蓟抑制MRSA41577外排系统的作用机制
2016-07-24 17:35:02袁琳慧王迪谢鲲鹏谢明杰
中国生化药物杂志 2016年11期
袁琳慧,王迪,谢鲲鹏,谢明杰
(辽宁师范大学生命科学学院 辽宁省生物技术与分子药物研发重点实验室,辽宁 大连 116081)
穿心莲和水飞蓟抑制MRSA41577外排系统的作用机制
袁琳慧†,王迪†,谢鲲鹏,谢明杰Δ
(辽宁师范大学生命科学学院 辽宁省生物技术与分子药物研发重点实验室,辽宁 大连 116081)
目的 研究穿心莲和水飞蓟抑制MRSA41577外排系统的作用机制。方法 通过荧光分光光度法检测穿心莲和水飞蓟为MRSA41577外排系统的抑制剂;通过PCR的方法检测MRSA41577的norA外排基因;通过RT-PCR的方法检测穿心莲和水飞蓟对外排基因norA表达量的影响。结果 穿心莲和水飞蓟处理MRSA41577后,环丙沙星在菌体内的的蓄积量随着药物处理时间的增加而增加,当药物处理菌体12 min后,与对照组相比,菌体内环丙沙星蓄积量分别增加49%和76%(P<0.05),其作用效果优于阳性对照利血平。MRSA41577菌株内含有norA外排基因。穿心莲和水飞蓟能降低norA基因表达量,与对照组相比,norA的相对表达量分别降低了35%和42%(P<0.05)。结论 穿心莲和水飞蓟抑制MRSA41577外排系统的作用机制是通过减少norA外排基因的表达量,进而抑制外排蛋白的合成,阻止药物的外排使菌体重新恢复对药物的敏感性。
耐甲氧西林金黄色葡萄球菌;外排系统;norA;作用机制
随着对细菌耐药性研究的深入,目前认为主动外排系统是细菌产生多重耐药性的重要机制[1-2]。主动外排系统是广泛分布于菌体细胞膜表面上的一种膜转运蛋白,它能够让进入到膜内的药物直接被泵出体外,使细胞内抗生素等药物的含量降低而产生耐药[3],这些膜转运蛋白由相关外排基因调控,如norA和qacA/B等[4]。其中norA是金黄色葡萄球菌的固有外排基因,当受到环丙沙星等药物的刺激诱导时,可通过增加该基因的表达量,使其编码的外排蛋白增多,从而将进入膜内的药物泵出膜外使其产生耐药[5-7]。近年研究表明,很多中药能够抑制MRSA的外排系统,是MRSA良好的外排泵抑制剂[8]。本实验室前期从24种中药中筛选到穿心莲和水飞蓟2种中药对MRSA的外排系统具有抑制作用。本文拟通过检测穿心莲和水飞蓟2种药物对菌体内环丙沙星蓄积量的影响,以及对外排基因norAmRNA转录水平的影响,探讨其抑制MRSA外排系统的作用机制,旨在为将其开发成外排系统抑制剂提供理论基础。
1 材料与方法
1.1 材料
1.1.1 菌株:耐甲氧西林金黄色葡萄球菌MRSA41577,由大连市中心医院提供。本文前期通过头孢西丁纸片证明其为耐药菌株。
1.1.2 主要试剂:穿心莲(Andrographis paniculata,AP)和水飞蓟(silybum marianum,SM)由辽宁师范大学中药现代化实验室提供;利血平购自大连诺威信生物技术有限公司;甘氨酸、环丙沙星(CIP)购自上海生工生物有限公司;引物及PCR反应液购自大连宝生物公司。
1.2 方法
1.2.1 中药穿心莲和水飞蓟提取物制备方法:称取一定量的穿心莲和水飞蓟中药,用索氏提取器以95%的乙醇热回流提取8 h后将提取液浓缩制成干粉。精确称取一定量的干粉,用无水乙醇配制成一定浓度的母液,0.22 μm滤器过滤后备用。
1.2.2 环丙沙星标准曲线的制备[9]:精确称取一定量的环丙沙星粉末,用0.1M甘氨酸(pH=3)配制成浓度分别为0.4、0.8、1、2、4、6、8、10、12、14 mg/L的环丙沙星标准液。于454 nm发射波长和382 nm激发波长下测定环丙沙星的荧光值,以环丙沙星的浓度为横坐标,荧光值为纵坐标绘制标准曲线。
1.2.3 环丙沙星蓄积动力学测定[9]:用终浓度为16 mg/mL的穿心莲和水飞蓟处理MRSA41577 16 h后,离心收集菌体,PBS(pH=7.0)洗涤3次后,用PBS将其重新悬浮,37 ℃孵育10 min后加入环丙沙星,使其终浓度为20 μg/mL。待作用4、8、12 min后,分别取0.5 mL菌液,离心收集菌体,然后加入1 mL pH3.0的盐酸甘氨酸,25 ℃温浴2 h后离心,取上清液于454 nm发射波长和382 nm激发波长下测定荧光值,以加20 mg/L利血平为阳性对照,不加药组为空白对照。
1.2.4 MRSA41577norA外排基因的检测:提取模板DNA,根据GenBank中已发布的金黄色葡萄球菌的基因序列,利用Primer 5.0软件及参考有关文献,设计norA基因引物,norA扩增产物预测长度分别为435 bp。然后用进行PCR扩增,扩增条件为94 ℃ 5 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s,30个循环,72 ℃ 10 min。扩增产物用1%琼脂糖凝胶电泳进行检测,后用Gel-Pro Analyzer Version3.1凝胶成像仪进行分析。norA产物长度435 bp,序列如下:
上游引物5’-GTT ACT TGT TGC TGC TTT TG-3’
下游引物5’-GCT TGT CGT AGA CTT TTT CG-3’
1.2.5 穿心莲和水飞蓟对norA外排基因表达量的影响[10]:将培养至对数期的MRSA41577菌悬液分别接种到含终浓度为32 mg/mL的穿心莲和水飞蓟的LB培养基中,37 ℃,120 rpm培养16 h;取4~6 mL MRSA41577菌悬液,4 ℃,12000 r/min离心1 min,采用Trizol法提取菌体总RNA,使用微量核酸测量仪进行RNA浓度的定量,然后采用两步法反转合成模板cDNA。根据设计合成的norA引物进行扩增,以16S RNA为内参基因,待反应结束后分析RT-PCR的扩增曲线和溶解曲线,并计算norA基因的相对表达量,以不加药物组为空白对照。norA引物长度为21 bp,16S引物长度为21 bp,序列如下:
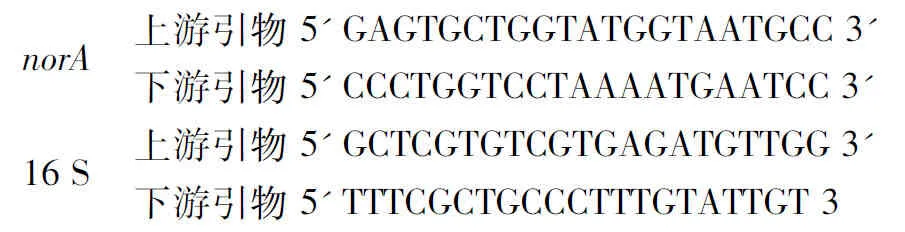
norA上游引物5GAGTGCTGGTATGGTAAT-GCC3下游引物5CCCTGGTCCTAAAATGAATCC316S上游引物5GCTCGTGTCGTGAGATGTT-GG3下游引物5TTTCGCTGCCCTTTGTATTGT3
1.3 统计学方法 采用SPSS17.0统计学软件进行数据统计分析,计数资料采用率表示,组内比较采用单因素方差分析,组间比较采用t检验,以P<0.05表示差异具有统计学意义。
2 结果
2.1 穿心莲和水飞蓟对MRSA41577体内环丙沙星蓄积量的影响 实验结果显示,环丙沙星在0.2~14 mg/L浓度间其荧光强度与浓度呈线性关系。见图1。
蓄积量实验结果显示,穿心莲和水飞蓟能够增加 MRSA41577 菌体内环丙沙星的蓄积量,且呈时间依赖。当穿心莲和水飞蓟作用菌体12 min后,菌体内环丙沙星蓄积量与对照组相比分别增加了49%和76%(P<0.05),作用效果优于阳性对照利血平。见图2。

图1 环丙沙星标准曲线Fig.1 The standard curve of ciprofloxacin
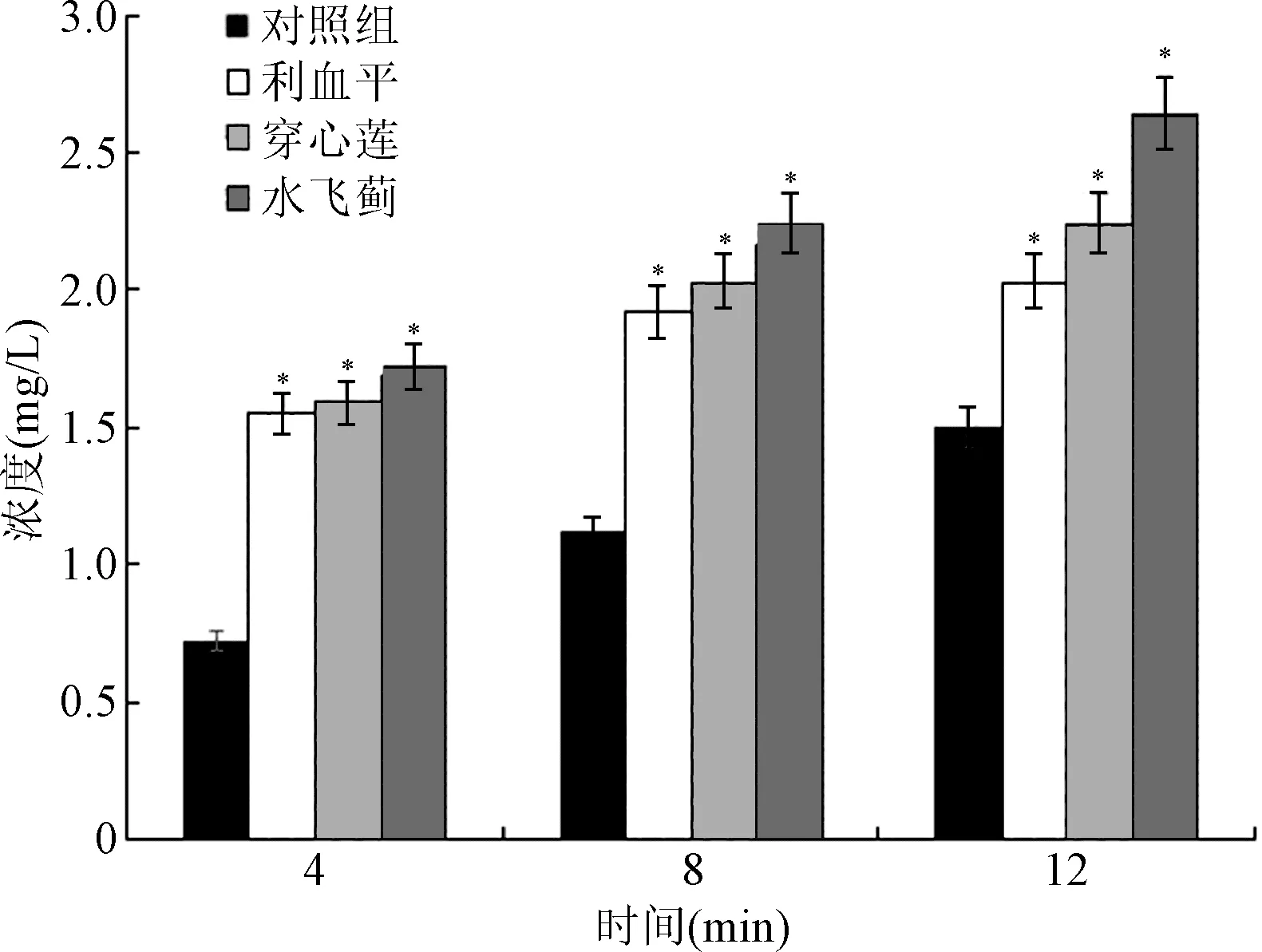
图2 穿心莲和水飞蓟对环丙沙星蓄积量的影响*P<0.05,与对照组比较Fig.2 Effect of AP and SM accumulation of ciprofloxacin*P<0.05,compared with control group
2.2 外排基因norA的检测 实验结果显示,在MRSA41577菌株中检测到norA外排基因,推测穿心莲和水飞蓟抑制MRSA41577的外排作用与norA外排基因有关。见图3。

图3 MRSA41577 norA基因的琼脂糖凝胶电泳图Fig.3 Agarose gel picture of norA gene of MRSA41577
2.3 穿心莲和水飞蓟对MRSA 414577norA外排基因表达量的影响 RT-PCR结果显示,穿心莲和水飞蓟能够显著降低MRSA 414577norA外排基因表达量,当穿心莲和水飞蓟与MRSA41577作用 16h后,其norA的表达量与对照组比较分别降低35%和42%(P<0.05)。见图4。
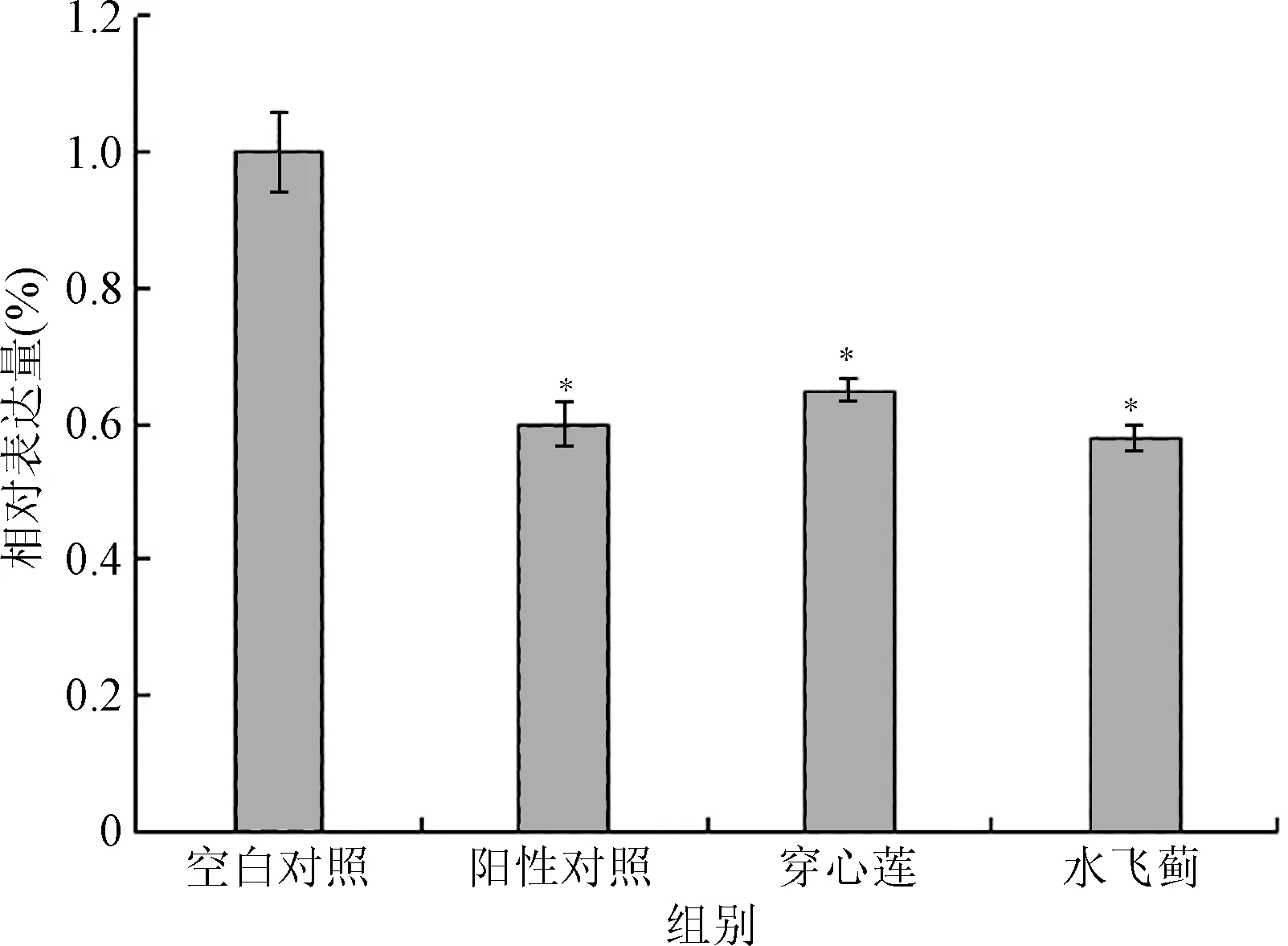
图4 穿心莲和水飞蓟对MRSA41577 norA表达量的影响*P<0.05,与对照组比较Fig.4 Effect of APand SM relative norA expression of MRSA41577.*P<0.05,compared with control group
3 讨论
主动外排系统是分布于菌体细胞膜表面上的组膜转运蛋白,其种类丰富,功能各异,由相关外排基因表达产生[11-13]。其中norA是金黄色葡萄球菌主要的多药外排蛋白,由位于染色体上的norA基因编码,是金黄色葡萄球菌的结构基因。在正常表达情况下不会导致细菌耐药,但受到环丙沙星等药物的刺激诱导后,其表达量明显增加,最终因norA蛋白的增多而将进入膜内的药物泵出膜外[14-15]。本研究实验结果显示,穿心莲和水飞蓟能显著提高环丙沙星在MRSA41577菌体内的蓄积量,与对照组相比环丙沙星在菌体内的蓄积量分别增加了49%和76%(P<0.05),其作用效果优于阳性对照利血平,表明穿心莲和水飞蓟是MRSA41577 的外排泵抑制剂。RT-PCR结果显示,穿心莲和水飞蓟可以显著降低norA基因的表达量,与对照组相比分别降低35%和42%(P<0.05),其作用效果与阳性对照利血平相当。
综上所述,水飞蓟和穿心莲是MRSA41577良好的外排泵抑制剂,其作用机制是通过抑制外排基因norA的表达量,减少norA外排蛋白的量,使菌体抑制药物外排,以提高环丙沙星等药物在MRSA41577菌体内的蓄积量,逆转细菌的耐药性。
[1] Otreebska-Machaj E,Chevalier J,Handzlik J,et al.Efflux Pump Blockers in Gram-Negative Bacteria:The New Generation of Hydantoin Based-Modulators to Improve Antibiotic Activity[J]. Front Microbiol,2016,7:622.
[2] 韩善桥,刘瑾红.细菌耐药的产生机制与控制措施[J].实用预防医学,2010,17(4):831-852.
[3] Du R,Han W,Lei L.The relation between fluoroquinolones- resistant and mRNA expression level of NorA gene in Staphylococcus aureus[J].Chinese Journal of Veterinary Scince,2006,26(1):66- 68.
[4] 任晓镤,陈伟,张利莉.甘草及醉马草水煎液对表皮葡萄球菌生物膜形成的影响[J].中国兽医学报,2013,33(1):125-128.
[5] Iwamote M,Mu Y,Lynfield R,et al.Trends in invasive Methicllin-resistant staphylococcus aureus infections[J].Pediatrics,2013,132(4):e817-824.
[6] 金春梅,朴美花,许顺姬,等.耐甲氧西林金黄色葡萄球菌的耐药性分析[J].中华医院感染学杂志,2013,23(13):3238-3242.
[7] Lomovskaya O,Watkins WJ.Efflux pumps:their role in antibacterial drug discovery[J].Curr Med Chem,2001,8(14):1699-1711.
[8] Li HB,Chen F.Simultaneous separation and purification of five bioactive coumarins from the Chinese medicinal plant Cnidium monnieri by hish-speed counter-current chromatography[J].J Sep Sci, 2005,28(3):268-272.
[9] 李洪涛,吴春民,覃慧敏,等.穿心莲内酯对铜绿假单胞菌PAO1株MexABOperM外排泵mRNA表达的影响[J].中华传染病杂志,2007,25(6),338-341.
[10] Yang Z,Yang X,Niu Y.Screening for and bioassay- guided isolation of bacterial efflux pump inhibitors by control method based on double plates[J].Nat Prod Res,2010,22(1):277-280.
[11] Li XZ,Plésiat P,Nikaido H.The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria[J].Clin Microbiol Rev,2015,28(2): 337-418.
[12] Wunderink RG.How important is methicllin-resistant Staphylococcus aureus as a cause of community-acquired pneumonia and what is best antimicrobial therapy[J].Infect Dis Clin North Am,2013,27(1):177-188.
[13] Kim HI,Kim JA,Choi EJ,et al.In vitro and in vivo antimicrobial efficacy of natural plant-derived compounds against Vibrio cholerae of O1 El Tor Inaba serotype[J].Biosci Biotechnol Biochem,2015,79(3):475-483.
[14] Thomas VC, Thurlow LR, Boyle D,et al.Regulation of autolysis-dependent extracellular DNA release by Enterococcus faecalis extracellular proteases influences biofilm development[J].J Bacteriol,2008,190(16):5690-5698.
[15] Ben CL,Margaret I,Clara BSL,et al.Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillinresistant Staphylococcus aureus(MRSA) and inhibition of MRSA Pyruvate kinase[J].J Ethnopharmacol,2011,137(1):767-773.
(编校:苗加会)
Inhibitory effects of andrographis paniculata and silybum marianum on the efflux pump of MRSA 41577
YUAN Lin-hui†, WANG Di†, XIE Kun-peng, XIE Ming-jieΔ
(School of Life Sciences, Liaoning Normal University, Key Laboratory of Biotechnology and Drug Discovery of Liaoning Province,Dalian 116081, China)
ObjectiveTo study the inhibitory effects of andrographis paniculata and silybum marianum on the efflux system of MRSA 41577.MethodsInhibitory effects of andrographis paniculata and silybum marianum on efflux system of MRSA 41577 was evaluated using fluorescence spectrophotometry. PCR was applied to detect thenorAefflux gene.By RT-PCR method for detection of andrographis paniculata and silybum marianum influence of the expression ofnorAefflux gene.ResultsAndrographis paniculata and silybum marianum significantly increased the accumulation of ciprofloxacin in MRSA 41577 in a time-dependent manner. At 12 minute, andrographis paniculata and silybum marianum respectively increased ciprofloxacin in MRSA41577 by 49% and 76%(P<0.05), which is superior to that of reserpine. Further mechanism studies indicated that andrographis paniculata and silybum marianumcould reduce the expression ofnorAin MRSA 41577. After incubated with andrographis paniculata and silybum marianum for 16 h, the relative expression ofnorAof MRSA41577 was respectively reduced by 35% and 42%(P<0.05).ConclusionAndrographis paniculata and silybum marianumcould inhibit MRSA efflux system through reducing pathogen ’s expression ofnorAand NorA protein.
MRSA; efflux system;norA; mechanism of action
10.3969/j.issn.1005-1678.2016.11.007
辽宁省教育厅科学研究一般项目(L201683675);辽宁省自然科学基金(201602462);国家级大学生创新创业训练计划项目(201610165042)
袁琳慧,女,本科,研究方向:生物科学,E-mail:2318446277@qq.com;王迪,共同第一作者,女,硕士在读,研究方向:微生物生化,E-mail:momoxiaodi@sina.com;谢明杰,通信作者,女,博士,教授,研究方向:微生物生化,E-mail:xmj1222@sina.com。
R285
A
猜你喜欢
湘潭大学自然科学学报(2022年2期)2022-07-28 05:26:24
天津中医药(2020年5期)2020-06-01 12:16:14
山东林业科技(2018年6期)2019-01-08 09:48:04
山东林业科技(2017年1期)2017-06-29 07:54:06
中成药(2017年4期)2017-05-17 06:09:27
现代检验医学杂志(2016年2期)2016-11-14 02:38:10
农业环境科学学报(2016年10期)2016-11-11 09:36:52
西安理工大学学报(2016年3期)2016-11-10 06:00:50
林业与生态(2016年2期)2016-02-27 14:23:42
特产研究(2015年1期)2015-04-12 06:36:19
