
水分胁迫对3个不同地区橡胶草生理特性的影响
2016-07-12 02:20徐建欣高玉尧孙正美刘实忠
干旱地区农业研究 2016年3期
关键词:聚乙二醇
徐建欣,杨 洁,高玉尧,吴 景,孙正美,刘实忠
(中国热带农业科学院湛江实验站/热带旱作农业研究中心, 广东 湛江 524013)
水分胁迫对3个不同地区橡胶草生理特性的影响
徐建欣,杨洁,高玉尧,吴景,孙正美,刘实忠
(中国热带农业科学院湛江实验站/热带旱作农业研究中心, 广东 湛江 524013)
摘要:本研究以3个不同地理来源的橡胶草群体为研究对象,使用不同浓度的聚乙二醇(PEG-600)溶液分别对3个供试群体成员苗期进行模拟水分胁迫,测定了其叶片相对含水量、叶绿素含量、抗氧化保护酶活性、渗透调节物质含量、丙二醛含量、叶片长度、叶片宽度以及最大根长。结果表明,随着胁迫浓度的提高3个供试群体的叶片相对含水量、叶绿素含量、抗氧化保护酶活性以及生长发育指标呈下降趋势,渗透调节物质含量以及丙二醛含量呈上升趋势;综合各指标表明,3个供试橡胶草群体抗旱性表现为昭苏群体 >石河子群体 >临夏群体。
关键词:橡胶草;水分胁迫;聚乙二醇;生理反应
我国农业中旱地农业比重很大,旱作农业有很大的增产潜力。广大干旱半干旱地区,如何在其可承受的限度内通过不同途径高效利用有限水资源以及在少雨年份或季节减轻干旱的不利影响,从总体上提高光合生产力,是发掘这一地区增产潜力的基本问题,所以加强旱作农业基础理论研究,特别是抗旱生理研究就显得特别重要,探讨干旱胁迫下作物生理指标的变化对于作物抗旱栽培具有重要意义。
橡胶草(Taraxacumkok-saghyzRodin, TKS),为菊科蒲公英属多年生草本植物,它原产于哈萨克斯坦、欧洲以及中国大陆的新疆等地,常生长在盐碱化草甸、河漫滩草甸及农田水渠边[1-2],中国新疆、甘肃、陕西以及东北、华北、西北等地也有野生种分布。橡胶草具有适应性强、适应区域广,生长收获期短,可在田间储存,并可轮种或间作,适合机械化种植和采收,具有较强的生长繁殖能力,能够生长在较寒冷的地区,容易种植,并且具有较强抗菌能力和抗虫害能力以及生物相容性好、抗过敏性能强等优点,使其成为具有发展前途的巴西橡胶树橡胶替代作物[3]。另外,橡胶草组织培养相对较为容易,遗传转化也更易显型,因此它也是研究植物产胶机理的良好模式植物和生物反应器,其根、叶中可以提取菊糖,提胶后的残渣可以加工酒精、沼气,并且具有一定药用价值。在原材料和能源日趋紧张的今天,橡胶草作为科研材料、工业原料和生物能源等均具有比较优势[4]。
目前我国对于橡胶草水分胁迫下的理化特性研究鲜见报道,仅有少量研究涉及干旱胁迫[5]、光照强度[6-7]以及盐胁迫[8]对蒲公英属其它近缘植物生理特性的影响。随着人们对天然橡胶需求量的迅猛增加,要求产量以更快的速度提高,因此今后天然橡胶产量的提高将主要依赖于新的产胶植物的发掘以及品种改良和新品种选育,加速改良进程的主要途径就是进一步了解限制产量提高的生理生化特征,从而减小育种的盲目性。天然橡胶产量的提高是个非常复杂的过程,它不仅取决于产胶植物自身的遗传特性,而且受生长期间环境条件的影响,所以我们需要了解天然橡胶产量提高过程中起重要作用的生理生化机理,才能更好为生产实际提供理论指导。
中国是世界天然橡胶消费大国,近年来的年消费量均在300万t以上,进口依存度达到80%以上。据预测,2030年中国天然橡胶消费量将达480万t。然而,受地理条件限制,中国适于种植橡胶树的区域不到国土面积的5%,实际可用于种植橡胶树的土地面积在1.2×106hm2左右,依靠扩大种植规模、扩大产能来保证中国天然橡胶供应非常不现实[4]。按中国目前的天然橡胶生产以及科技的发展态势,中国天然橡胶供需缺口还将长期存在,过度依赖国际市场的局面将在未来相当长的一段时间内不会发生改变,这严重威胁了我国的社会经济发展与国家战略安全。寻找其它天然橡胶源,扩大天然橡胶生产,确保天然橡胶可持续供应便显得尤为重要。橡胶草作为近年来新兴的重要产胶作物,在我国还缺乏系统的研究与完善的保护策略,这使得很多优秀的种质资源尚未得到有效的保护与利用便在地球上永远地消失了。因此,深入研究水分胁迫下影响橡胶草抗旱性的重要生理生化因子,阐明这些生理生化特性与橡胶草抗旱性及橡胶产量的关系,进一步探讨橡胶草应对干旱胁迫的调节机制,对于橡胶草新品种的选育及种植产业化发展具有重要意义。
1材料与方法
1.1供试材料
本研究所使用的橡胶草种质资源分别采集自新疆维吾尔自治区昭苏县胡松图喀尔逊蒙古族乡74兵团、新疆维吾尔自治区石河子市北湖以及甘肃省临夏回族自治州永靖县刘家峡水库。
1.2实验设计
选择子粒饱满、健康的种子,使用10%次氯酸钠消毒,清水冲洗4~5次,33℃催芽露白后,播种于装有营养土的塑料盆中,待幼苗长至5~7叶龄时从塑料盆中移出,尽量不损伤根系。选取长势一致的植株,移入用草炭∶珍珠岩∶蛭石(体积比2∶1∶1)的混合土作为基质的悬浮育苗盘中,培养于人工气候箱,温度25℃,湿度70%~80%,每天照光16 h。使用木村营养液[9]进行培养(pH 6.5),7 d后开始使用不同浓度PEG(PEG-600)处理T0=0%(CK)、T1=5%(轻度干旱)、T2=10%(中度干旱)、T3=20%(严重干旱),处理48 h后开始对供试材料进行各项生理指标检测,实验重复3次,测试叶片一般采用位于整株靠近中心的第2层叶片(上数第4~6片叶)测定。
1.3测定指标与方法
每个重复随即选择10个单株进行生理指标检测,叶片相对含水量采用烘干称重法[5],叶绿素含量采用丙酮乙醇法[6],SOD(超氧化物歧化酶)活性采用NBT(氮蓝四唑)法[10],POD(光氧化物酶)活性采用愈创木酚比色法[10],CAT(过氧化氢酶)活性采用滴定法[11],可溶性糖含量采用蒽酮比色法[10],Pro(脯氨酸)含量采用茚三酮显色法[10],MDA(丙二醛)含量采用硫代巴比妥酸显色法[10]。
1.4数据处理
使用Excel 2003和Minitab 15软件[12]进行数据处理与图表制作。
2结果与分析
2.1PEG模拟干旱对不同地区橡胶草叶片相对含水量的影响
由表1可以看出,随着PEG胁迫浓度的增加,来自3个不同地区的橡胶草叶片相对含水量均表现为不断下降的趋势。当PEG胁迫浓度为T3时,昭苏橡胶草群体较T0下降了7%,石河子群体与临夏群体分别下降了16.5%与21.3%。差异显著性分析表明,昭苏群体在各个胁迫浓度下均显著高于其其它两个群体(石河子、临夏),而石河子群体与临夏群体之间不存在显著差异,说明来自昭苏的橡胶草群体具有更强的叶片保水能力。
2.2PEG模拟干旱对不同地区橡胶草叶片光合特性的影响
叶绿素参与光合作用过程中光能的吸收、传递和转化是植物生长发育不可或缺的关键物质,叶绿
注:同一列内带有相同字母者表示多重比较差异未达5%显著水平。
Note: Values within a column followed by the same letters are not significantly different at 0.05 probability level.
素含量在一定程度上反映了植物光合作用的能力。由表2可知,三个不同地区的橡胶草叶片叶绿素a、叶绿素b以及总叶绿素含量均随着水分胁迫程度的加深呈不断下降的趋势,但昭苏群体的下降幅度(53.4% T3)小于其它两个群体(石河子58.5%,临夏59.6% T3)且在不同胁迫浓度均高于其它两个群体,表现出对干旱胁迫更好的耐受能力。叶绿素a/b亦随着水分胁迫程度的加深而不断下降,表明叶绿素a更易受到水分胁迫的影响。
2.3PEG模拟干旱对不同地区橡胶草抗氧化保护酶的影响
由表3可知,随着PEG胁迫浓度的增加昭苏群体的SOD活性呈现先缓慢提高后迅速下降的趋势,而其它两个群体(石河子、临夏)则表现为先下降后上升而后又下降的趋势(所有胁迫浓度活性均低于T0对照)。3个群体POD活性均表现为先上升后下降的趋势且所有胁迫浓度活性均高于T0对照,但不同之处在于昭苏群体的POD活性峰值来自T1而其它两个群体(石河子、临夏)的POD活性峰值来自T2。CAT活性表现为昭苏群体先下降后上升而后又下降的趋势,而其它两个群体(石河子、临夏)则表现为随着PEG胁迫浓度的增加不断下降的趋势。差异显著性分析表明,不同胁迫浓度下昭苏群体的抗氧化保护酶活性均显著高于其它两个群体(石河子、临夏),而石河子群体的抗氧化保护酶活性高于临夏群体,但并非所有胁迫浓度下都具有显著差异,这表明昭苏群体的橡胶草种质资源在所有3个群体中具有较好的抗氧化保护能力,从一个侧面表现出昭苏群体具有较强水分胁迫耐受能力。
注:不同大写字母表示相同胁迫程度不同地区间有显著差异(P<0.05);不同小写字母表示地区内不同胁迫程度有显著差异(P<0.05)。下同。
Note: Values within the same stress level between different regions by the different capital letters are significantly differeence at 0.05 probability level; Values within the same region stress level between different stress levels by the different lowercase letters are significantly differeence at 0.05 probability level. The same as below.
2.4PEG模拟干旱对不同地区橡胶草渗透调节物质与丙二醛的影响
由表4可知,昭苏群体渗透调节物质(可溶性糖、脯氨酸)含量随胁迫浓度的加深呈现先上升后下降的趋势且峰值出现在T2下,T3时可溶性糖与脯氨酸含量分别较对照T0上升51%、75%;石河子群体则呈一直上升的趋势,T3时可溶性糖与脯氨酸含量分别较对照T0上升48%、73%;临夏群体与石河子群体变化趋势一致,T3时可溶性糖与脯氨酸含量分别较对照T0上升43%、39%。差异显著性分析表明,相同胁迫浓度下三个群体间渗透调节物质含量均存在显著差异,且表现为昭苏群体>石河子群体>临夏群体。随胁迫浓度的加深,三个群体的丙二醛含量均呈不断上升的趋势,相同胁迫浓度下三个群体间丙二醛含量均存在显著差异,且表现为昭苏群体<石河子群体<临夏群体。
2.5PEG模拟干旱对不同地区橡胶草生长发育的影响
由表5可知,水分胁迫下3个橡胶草群体的叶片长度、叶片宽度以及最大根长均呈现先升高后降低的趋势,表明低、中浓度的干旱胁迫可以促进橡胶草幼苗的生长发育,而在高浓度胁迫下由于生物膜系统的破坏以及有害自由基的积累等问题,造成橡胶草植株生长发育缓慢。T3胁迫下昭苏群体的叶片长度、叶片宽度以及最大根长分别较之T0下降了15.4%、29.7%和14.7%,在生长速度以及下降幅度上均优于其它两个群体(石河子 24.7%、40%和21.8%;临夏28.5%、47.1%和25.2%)且相同胁迫浓度下三个群体间生长发育指标均存在显著差异,表明昭苏群体的抗旱能力最强,石河子群体次之,临夏群体表现最差,这一结果与之前的生理指标表现一致。
3讨论与结论
叶片是植物进化过程中对环境变化比较敏感、可塑性较强的器官,也是植物进行生理代谢活动的主要场所[13]。叶片在水分胁迫下含水量的高低,一定程度上代表叶片的保水能力的强弱,水分胁迫下叶片相对含水量越高、下降速率越慢的研究材料抗旱能力越强。本研究结果表明,干旱胁迫可降低橡胶草叶片相对含水量导致叶片失水,三个供试群体中昭苏群体的叶片相对含水量下降幅度最小、且在各个胁迫浓度均显著高于其它两个群体(石河子、临夏),表明昭苏群体的橡胶草叶片保水能力更强且具有较好的抗旱能力,而石河子群体叶片相对含水量与临夏群体不存在显著差异,表明这两个群体叶片持水能力相近。
水分胁迫使植物细胞内核糖体的形成受到影响,从而抑制叶绿体片层中捕光Cha/b-Pro复合体的合成,影响叶绿素积累,降低离体叶绿体的放氧率,以及破坏叶绿体的类囊体结构,抑制光合磷酸化等[14-16],且叶绿素a对活性氧的反应比叶绿素b敏感,其破坏程度往往大于叶绿素b,所以叶绿素总量和叶绿素a/b比值会下降,其下降程度与供试材料的抗旱性呈显著负相关关系[17-18]。本研究结果表明,水分胁迫可降低橡胶草叶片内叶绿体含量,从而影响橡胶草叶片的光合效率导致植物生长发育受到抑制。供试的三个橡胶草群体中,昭苏群体的叶绿素含量下降幅度最小,且在大多数胁迫浓度下均与其它两个群体(石河子、临夏)存在显著差异,据此表明昭苏群体具有较强的抗旱能力。石河子群体与临夏群体在部分胁迫浓度下亦存在显著差异,且叶绿素含量下降幅度小于临夏群体,说明石河子群体的抗旱能力优于临夏群体。
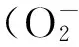
植物细胞只有在含有一定量的水分和维持一定的压力势时才能发挥正常的生理功能。植物细胞的压力势(膨压)是通过渗透调节作用来保持的[29]。渗透调节是指植物在水分胁迫下除了失水被动浓缩外,细胞主动通过代谢活动形成渗透调节物质,提高细胞内溶质浓度,降低水势,从外界水分减少的介质中继续吸水,维持一定的膨压,因而使植物能够进行正常的代谢活动和生长发育,适应逆境胁迫[30]。常见的渗透调节物质有无机离子如K+、可溶性糖、游离氨基酸、脯氨酸等[31-33],在干旱条件下增加细胞渗透调节能力的关键是细胞内渗透调节物质的主动积累,以维持较高的膨压,阻止植物脱水,增加根系生长,促进水分吸收[34-35]。可溶性糖是植物叶片细胞内的一种渗透调节物质,可溶性糖的主动积累,可以参与降低植物体内渗透势,以利于植物在干旱逆境下维持植物体正常生长所需水分,从而提高抗逆性[36]。脯氨酸有较好的水合作用,其溶解度比其它常见氨基酸都大,从而提高了原生质水溶液的渗透压。本研究结果表明,水分胁迫可以促进3个供试橡胶草群体叶片内可溶性糖以及脯氨酸含量的积累,从而提高细胞内渗透压力,减轻干旱对植物细胞内膜系统及各细胞器的伤害。3个供试橡胶草群体中,昭苏群体的渗透调节物质较其它两个群体(石河子、临夏)积累速度更快,对于水分胁迫的反应也更加积极敏感,表现出最强的抗旱能力,而临夏群体在各个胁迫浓度均低于其它两个群体(昭苏、石河子),在渗透调节能力方面表现较差,这与之前叶片相对含水量以及抗氧化保护酶活性的变化趋势相近,能够体现出临夏群体在3个供试群体中抗旱性表现是最差的。
植物器官衰老时或在逆境条件下,往往发生膜脂过氧化作用,丙二醛(MDA)是其产物之一,通常利用它作为膜脂过氧化指标,表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱。本研究显示,3个供试橡胶草群体叶片内丙二醛含量均随水分胁迫浓度的增加而不断积累,表明干旱胁迫对橡胶草的细胞膜造成了一定程度的伤害,而昭苏群体的丙二醛含量上升幅度最小,这可能与其抗氧化保护酶活性较高有关。
水分胁迫对植物生长发育的影响是鉴定该植物抗旱能力强弱的重要指标,本研究表明昭苏群体在PEG模拟干旱胁迫下的生长发育指标受影响程度优于其它两个群体,这与之前的生理指标检测结果一致,也相互印证了我们对3个群体抗旱能力强弱的判断。
综合叶片相对含水量、抗氧化保护酶活性、渗透调节物质、丙二醛含量以及生长发育指标的研究结果表明,3个供试橡胶草群体抗旱性表现为昭苏群体 >石河子群体 >临夏群体。出现这一结果的原因可能与这3个群体的原始生境有关,昭苏群体生长在降雨较少的半干旱草原上,因而伴随植物与环境的长期互作而形成了较强的抗旱能力,而石河子群体与临夏群体生长在降雨相对充沛的水源附近,因水分供给相对充足使得这两个群体的成员极少面对极端干旱环境,使得它们的抗旱性表现较差。
随着我国经济的不断发展,天然橡胶需求的供需矛盾必将日益突出,橡胶草作为具有发展前景的三叶橡胶树替代植物在我国的发展还相对滞后,本文对于来自3个不同地理群体的橡胶草种质资源水分胁迫下的生理特性研究进行研究,不仅揭示了橡胶草植株内部各生理指标的变化规律,并且确定了各供试群体的抗旱性强弱表现,发掘出抗旱性表现较好的橡胶草种质资源(昭苏群体),能为未来的橡胶草抗旱机制研究以及橡胶草抗旱育种提供相关的研究材料。
参 考 文 献:
[1]林伯煌,魏小弟.橡胶草的研究进展[J].安徽农业科学,2009,37(13):5950-5951.
[2]罗士苇,吴相钰,冯午.橡胶草的研究部分Ⅱ:新疆产橡胶草的化学分析及其橡胶含量之测定[J].中国科学,1951,2(3):381-387.
[3]van Beilen J B, Poirier Y. Establishment of new crops for the production of natural rubber[J]. Trends in Biotechnology,2007,25(11):522-529.
[4]赵平娟,安锋,林位夫,等.大力开展巴西橡胶树替代产胶植物及技术研发的建议[J].中国农学通报,2012,28(34):124-130.
[5]吴嘉雯,王庆亚.干旱胁迫对野生和栽培蒲公英抗性生理生化指标的影响[J].江苏农业学报,2010,26(2):264-271.
[6]赵英明,范文丽.光照强度对蒲公英光合特性的影响[J].辽宁农业科学,2009,(5):28-31.
[7]赵磊,杨延杰,林多.光照强度对蒲公英光合特性及品质的影响[J].园艺学报,2007,34(6):1555-1558.
[8]张新果,陈显扬,姜丹,等.耐盐药蒲公英(TaraxacumofficinaleWeber)愈伤组织筛选及生理生化特性分析[J].生物工程学报,2008,24(7):1202-1209.
[9]许明学,朴昌浩,金滢完.水稻在不同营养液中生育情况的研究[J].延边大学农学学报,1984,(1):14-19.
[10]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[11]张宪政,陈凤玉,王荣富.植物生理学实验技术[M].沈阳:辽宁科学技术出版社,1994.
[12]Minitab Inc, Minitab 15: Statistical Software[M]. Nittany Valley: State College PA,2007.
[13]宁伟,吴杰,赵婷,等.东北地区5种蒲公英叶绿体超微结构及光合特性研究[J].中国中药杂志,2012,37(10):1372-1377.
[14]卢从明,张其德,匡廷云,等.水分胁迫抑制水稻光合作用的机理[J].作物学报,1994,20(5):601-606.
[15]Lawlor D W. Limitation to photosynthesis in water-stressed leaves: stomata vs. metabolism and the role of ATP[J]. Annals of Botany, 2002,89(7):871-885.
[16]Parry M A J, Andralojc P J, Khan S, et al. Rubisco activity effects of drought stress[J]. Annals of Botany, 2002,89(7):833-839.
[17]张明生,谈锋.水分胁迫下甘薯叶绿素a/b比值的变化及其与抗旱性的关系[J].种子,2001,116(4):23-25.
[18]周海军,顾洪如,沈益新,等.干旱胁迫下两种钝叶草的生长和生理响应差异[J].江苏农业学报,2008,24(6):805-809.
[19]陈静,万佳,高晓玲,等.水稻抗旱生理及抗旱相关基因的研究进展[J].中国农学通报,2006,22(2):56-60.
[20]李成业,熊昌明,魏仙君.中国水稻抗旱研究进展[J].作物研究,2007,20(5):426-429.
[21]马余平,李道远.栽培稻抗旱性研究进展[J].广西农业科学,2007,38(4):398-403.
[22]熊正英,张志勤.POD活性与水稻抗旱性的关系[J].陕西师范大学学报(自然科学版),1995,23(4):63-66.
[23]田小磊,吴晓岚,李云,等.盐胁迫条件下γ-氨基丁酸对玉米幼苗SOD、POD及CAT活性的影响[J].实验生物学报,2005,38(1):75-79.
[24]周宝利,孟兆华,李娟,等.水分胁迫下嫁接对茄子生长及其生理生化指标的影响[J].生态学杂志,2012,31(11):2804-2809.
[25]徐建欣,杨洁,刘实忠,等.干旱胁迫对云南陆稻幼苗生理特性的影响[J].中国农学通报,2014,30(27):145-152.
[26]杨梅焕,姚顽强,曹明明,等.毛乌素沙地东南缘沙漠化共有种植物叶片抗氧化酶活性研究[J].西北农林科技大学学报(自然科学版),2012,40(7):203-208.
[27]Li Yan, Zhao Hongxia, Duan Baoli, et al. Effect of drought and ABA on growth, photosynthesis and antioxidant system ofCotinuscoggygriaseedlings under two different light conditions[J]. Environmental and Experimental Botany, 2011,71(1):107-113.
[28]杨文权,寇建村,雷忠萍,等.小冠花抗氧化保护系统对干旱胁迫及复水的动态响应[J].草地学报,2013,21(2):316-321.
[29]胡颂平,周清明.陆稻抗旱性研究进展[J].湖南农业大学学报(自然科学版),2001,27(3):240-244.
[30]陈展宇.旱稻抗旱解剖结构及其生理特性的研究[D].长春:吉林农业大学,2008.
[31]Iyer S, Caplan A. Products of proline catabolism can induce osmotically regulated genes in rice[J]. Plant Physiology, 1998,116(1):203-211.
[32]王贺正,马均,刘慧远.水稻抗旱性研究现状与展望[J].中国农学通报,2005,21(1):110-113.
[33]姚觉,于晓英,邱收,等.植物抗旱机理研究进展[J].华北农学报,2007,22(增刊):51-56.
[34]刘祖祺,张石城.植物抗性生理学[M].北京:中国农业出版社,1994.
[35]Fukai S, Cooper M. Development of drought resistant cultivars for rainfed lowland rice-experience from northeast Thailand and surrounding area[M]. Phnom Penh: Cambodian Agricultural Research & Development Institute, 2011,185-194.
[36]余玲,王彦荣,Garnett T,等.紫花苜蓿不同品种对干旱胁迫的生理响应[J].草业学报,2006,15(3):75-85.
Effects of water stress on physiological characteristics ofTaraxacumkok-saghyzRodin (TKS) populations from three different regions
XU Jian-xin, YANG Jie, GAO Yu-yao, WU Jing, SUN Zheng-mei, LIU Shi-zhong
(ZhanjiangExperimentalStation,ChineseAcademyofTropicalAgriculturalSciences/TropicalDryFarmingResearchCenter,Zhanjiang,Guangdong524013,China)
Keywords:Taraxacumkok-saghyzRodin(TKS); drought stress; polyethylene glycol; physiological response
Abstract:Three TKS populations from different regions were taken as the test materials in this paper, and the effects of simulated drought stress with different concentration of polyethylene glycol (PEG6000) were investigated on their leaf water content, chlorophyll content, activity of antioxidant protective enzyme, content of osmotic adjustment and MDA, leaf length, leaf width and maximum root length. The results showed that leaf water content, chlorophyll content, activity of antioxidant protective enzyme and growth and development indexes became decreased and content of osmotic adjustment and MDA went increased with the increase of drought stress. All of these variations suggested that drought resistance capabilities of three TKS populations from different regions are in the order of Zhaosu population>Shihezi population>Linxia population.
文章编号:1000-7601(2016)03-0153-07
doi:10.7606/j.issn.1000-7601.2016.03.24
收稿日期:2015-07-10
基金项目:中国热带农业科学院湛江实验站科研启动专项经费(zjky201303)
作者简介:徐建欣(1984—),男,助理研究员,博士,主要从事作物逆境生理研究。 E-mail:xujianxin0713@qq.com。 通信作者:刘实忠(1970—),男,研究员,博士,主要从事橡胶草种质资源收集、保护与利用研究。 E-mail:szliu@126.com。
中图分类号:S576; Q945.78
文献标志码:A
猜你喜欢
宁夏医学杂志(2020年4期)2021-01-21
化学研究(2020年4期)2020-11-06
证券市场红周刊(2020年30期)2020-08-09
安徽化工(2019年1期)2019-03-04
中国医药指南(2017年3期)2017-11-13
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
中国学术期刊文摘(2016年8期)2016-02-13
中国当代医药(2015年10期)2015-03-01
中国当代医药(2015年8期)2015-03-01
中国当代医药(2015年1期)2015-03-01
