
白绒山羊PPARγ基因RNA干扰慢病毒载体的构建及对肌内脂肪细胞增殖分化的影响
2016-07-11 07:39李金泉陈秀娟
畜牧兽医学报 2016年4期
杜 琛,李金泉,陈秀娟*
(1.内蒙古医科大学附属医院,妇产科生殖医学中心,呼和浩特 010050;2.内蒙古农业大学动物科学学院,动物遗传育种与繁殖自治区重点实验室,呼和浩特 010018)
白绒山羊PPARγ基因RNA干扰慢病毒载体的构建及对肌内脂肪细胞增殖分化的影响
杜琛1,李金泉2*,陈秀娟1*
(1.内蒙古医科大学附属医院,妇产科生殖医学中心,呼和浩特 010050;2.内蒙古农业大学动物科学学院,动物遗传育种与繁殖自治区重点实验室,呼和浩特 010018)
摘要:本研究以绒山羊肌内脂肪细胞为试验材料,通过靶向阻断过氧化物酶体增殖物激活受体PPARγ基因,建立稳定干扰的肌内脂肪细胞株,探讨绒山羊PPARγ基因在肌内脂肪细胞增殖和分化过程中的功能。采用慢病毒质粒包装系统构建特异靶向白绒山羊PPARγ基因的RNA干扰(RNA interference,RNAi )慢病毒载体,用以建立PPARγ基因稳定沉默的肌内脂肪细胞株。利用实时定量和Western blot 方法检测PPARγ基因在干扰组和对照组不同时间点的表达情况,并通过MTT和油红O染色法研究绒山羊PPARγ基因对肌内脂肪细胞增殖和分化的影响。经测序证实合成的含PPARγ-shRNA慢病毒载体寡核苷酸链插入正确,对肌内脂肪细胞的感染效率为80%以上,荧光定量和Western blot检测慢病毒介导的shRNA可以有效降低PPARγ的表达,其mRNA和蛋白水平作用的时间相隔24 h。沉默PPARγ基因后,脂肪细胞内甘油三脂的浓度明显低于对照组,相反MTT检测干扰组中细胞增殖能力高于对照组。本研究构建了绒山羊shRNA-PPARγ慢病毒干扰载体,成功转染至肌内脂肪细胞后可明显抑制脂肪细胞的分化,而促进肌内脂肪细胞的增殖,为进一步研究PPARγ基因在绒山羊脂肪细胞代谢通路中的作用及脂肪沉积机制奠定基础。
关键词:肌内脂肪细胞;RNA干扰;PPARγ
肌内脂肪(Intramuscular fat,IMF)是脂肪沉积的一种主要形式,具有一般脂肪所具有的特点。肌内脂肪直接参与了肉质嫩度、风味和多汁性的形成[1]。脂肪细胞是研究脂肪形成的理想模型,体外培养前体脂肪细胞不仅能完整的认识脂肪组织发生、增殖和分化的全过程,同时可以直接观察各种因素在整个生成过程中的规律,因此调控脂肪沉积并改善动物产品质量在畜牧业生产中具有重要意义。
脂肪细胞的分化是在多种转录因子的精细调控下完成的,在此过程中逐步表达那些能量合成代谢过程中相关的功能蛋白,其中最主要的分化转录因子是过氧化物酶体增殖物激活受体(Proxisome proliferators activated receptor,PPARs),属于核内类固醇激素受体超家族,在调节脂肪分化、炎症反应和免疫应答过程中起重要作用。PPARs分为3种不同的亚型,即PPARα、β、γ,具有组织特异性。其中PPARγ在脂肪分化和发育的过程中起到分子开关的作用,与C/EBPs家族和SREBPs家族协同作用,调控其靶基因的表达[2-7]。
RNAi技术是近年来发展起来的一种基因阻断技术,可以高效特异的阻断基因表达[8]。RNA干扰的有效性主要受基因载体转移效率的制约,目前最常见的哺乳动物载体是慢病毒表达载体(Lentivirus),它可以高效转染任何一种细胞[9]。因此,本研究通过构建PPARγ基因的小发夹RNA(Short hairp in RNA,shRNA)慢病毒载体并进行包装鉴定,筛选得到稳定株,进一步研究PPARγ沉默后对脂肪细胞增殖和分化的影响。
1材料与方法
1.1材料
本实验室所构建的内蒙古白绒山羊肌内前体脂肪细胞[10]、293T细胞、慢病毒的包装细胞,为贴壁依赖型成上皮样细胞、大肠杆菌菌株DH5a、pHBLV-U6-Puro(目的片段插入BamH I,EcoR I酶切位点,由U6启动子调控表达,该慢病毒载体中含有PGK启动子调控的Puro抗性基因表达)质粒。
1.2PPARγ-shRNA载体的构建和鉴定
在GenBank中查到山羊过氧化物酶体增殖物激活受体(Peroxisome proliferator activated receptor gamma,PPARs)(登录号JQ266369.1)的mRNA序列,从起始密码子下游75 bp处开始,长度为19~25 nt,GC含量为30%~50%的且与其他基因无明显同源性的序列作为siRNA靶点序列,序列为5′-GGGAAAGACGACAGACAAATT-3′,以靶点序列为正义链,互补链为反义链。并构建到shRNA中,序列:5′-GGGGAAAGACGACAGACAAAT-TTTCAAGAGAAATTTGTCTGTCGTCTTTCC-CTTTTTT-3′;5′-AAAAAAGGGAAAGACGAC-AGACAAATTTCTCTTGAAAATTTGTCTGT-CGTCTTTCCCC-3′。把合成的上下游引物溶解于退火缓冲液中,与经BamH I、EcoR I双酶切的pHBLV-U6-Puro载体连接,形成重组质粒pLVX-puro-PPARγ-shRNA。然后用一步法转化DH5α菌,挑选阳性克隆,将菌液送上海桑尼公司测序。
1.3慢病毒的包装和浓缩
对293T细胞复苏传代,细胞长至80%~90% 即可进行转染,按顺序加入10 μg pspax,10 μg pMD2G,10 μg pLVXpuro-PPARg-shRNA。转染后12~24 h,将培养基弃去,加入10 mL 10% DMEM开始收集病毒。0.45 μm滤器过滤,7 200 r·min-1离心120 min,之后用500 mL PBS重悬病毒沉淀,置于4 ℃保存。
1.4目的细胞慢病毒感染
将细胞铺盘于6孔板,每孔为5×105个细胞,每孔加入MOI=30慢病毒pLVX-puro-PPARγ-shRNA,24 h后换液。48 h后,换以含有2 μg·mL-1的puromycin的培液继续培养,每2~3 d更换培养液,待细胞生长稳定之后,可以进行细胞传代,2代之后无需加puromycin培养,建系完成。一部分细胞用于保种,另一部分细胞用于后续验证。
1.5基因表达的检测及数据处理
1.5.1荧光定量检测PPARγ的mRNA表达水平根据GenBank中登录的山羊过氧化物增殖物激活受体(Perixusome Proliferatoractivated receptors,PPARs),用Primer Premier 5.0 软件设计引物F:5′-CCGGAGGACGATCAGATTG-3′,R:5′-CGCCCAAACCTGATGGCAT-3′ 由上海生工生物工程有限公司合成。所有试验均设置3次重复,试验所得数据以“平均值±标准差”表示,采用Graph Pad Prism 5统计分析软件中的One-way ANOVA 进行方差分析,显著性水平为P<0.05。
1.5.2Western blot检测PPARγ的表达水平将筛选得到沉默的PPARγ稳定株分入细胞培养瓶中,经过转膜、封闭、抗体孵育,在不同时间点收集细胞,检测siRNA对PPARγ蛋白表达的沉默效率。所有试验均设置3次重复,使用Genpro32分析软件检测PPARγ在不同时间点蛋白条带灰度,以β-actin为参照。
2结果
2.1载体测序结果
pHBLV-U6-Puro质粒载体使用BamH Ⅰ和EcoR Ⅰ双酶切,之后进行回收扩增,并连接形成重组质粒pLVX-puro-PPARγ-shRNA。测序结果表明,合成的 PPARγ-shRNA核苷酸序列插入正确,中间为产生针对靶基因的 shRNA寡核苷酸(图1)。
2.2PPARγ-shRNA干扰组转染293T细胞
将测序验证后的PPARγ慢病毒载体质粒系统,pspax2、pMD2G、和pLVXpuro-PPARg-shRNA共转染至293T细胞,其转染效果如图2所示,有80%以上的细胞带有绿色荧光标记,说明慢病毒载体成功转至293T细胞中,可以用于后续慢病毒的大量包装。
2.3慢病毒转染绒山羊肌内脂肪细胞
将细胞分为3组:未处理的细胞为空白对照组(WT),空载体转染的细胞为阴性对照组(NC),慢病毒载体pLVX-puro-PPARγ-shRNA转染的肌内脂肪细胞为转染组(SH)。观察转染组细胞未出现大面积漂浮的死细胞,说明慢病毒载体构建成功,得到细胞稳定株(图3)。
2.4Real-time检测干扰后PPARγ基因在脂肪细胞不同时间点的表达情况
实时定量检测结果证实,转染组PPARγmRNA表达量在干扰24~72 h显著低于空白组(WT)和阴性对照组(NC)(P<0.05),其中在24 h干扰效果最为明显。空白组和对照组间PPARγmRNA表达量无显著差异(图4)。
2.5Western blot检测干扰后PPARγ蛋白在脂肪细胞不同时间点的表达情况
Western blot检测结果证实,干扰组PPARγ在48 和72 h蛋白表达量显著低于对照组 (P<0.05)(图5)。使用Genpro32分析软件检测PPARγ在不同时间点蛋白条带灰度,以β-actin为参照(图6)。
2.6下调PPARγ基因表达量对绒山羊肌内脂肪细胞增殖的影响
利用MTT法检测PPARγ基因表达量降低后脂肪细胞的增殖情况。干扰时间点记为0点,在干扰后每隔12 h分别对干扰组和对照组的细胞增殖情况进行检测。结果如图7所示,干扰后0~24 h干扰组和阴性对照组无显著差异,干扰后36~72 h干扰组细胞增殖能力明显高于阴性对照组。
2.7下调PPARγ基因表达量对绒山羊肌内脂肪细胞分化的影响
利用油红O染色的方法,提取萃取液,OD510 nm检测PPARγ基因表达量下降后脂肪细胞中甘油三酯含量的变化。干扰时间点记为0点,干扰后每隔12 h分别对干扰组和阴性对照组的细胞分化情况进行检测。细胞形态学观察,干扰组中被染色液附着的红色脂滴与阴性对照组相比明显减少(图8)。分光光度计检测干扰12~72 h,干扰组中细胞分化能力显著低于对照组(P<0.05)(图9)。

图1 PPARγ-shRNA的测序结果Fig.1 Sequencing result of PPARγ-shRNA

A.pLVX-puro-PPARg-shRNA转染293T细胞;B.pLVX-puro-PPARg-shRNA 转染293T细胞荧光图A.pLVX-puro-PPARg-shRNA transfected 293T cell;B.pLVX-puro-PPARg-shRNA transfected 293T cell图2 慢病毒质粒转染 293T 细胞荧光图 200×Fig.2 Lentiviral vector transfected 293T cells 200×

A.未处理的肌内脂肪细胞;B.空载体转染的肌内脂肪细胞;C.慢病毒转染的肌内脂肪细胞A.Intramuscular adipocytes untreated;B.Empty vector transfected intramuscular fat cells;C.Lentivirus vector transfectd intramuscular adipocytes图3 慢病毒质粒转染肌内脂肪细胞 200×Fig.3 Lentiviral vector transfected intramuscular adipocytes 200×

不同字母表示差异显著(P<0.05)。图6同Different letters indicate significant difference (P<0.05).The same as Figure 6图4 Real-time PCR检测不同时间点PPARγ基因在脂肪细胞中的表达量Fig.4 Real-time PCR detected PPARγ gene expression levels at different time in adipocytes

1~3.空白组24、48、72 h;4~6:阴性对照组24、48、72 h;7~9.干扰组24、48、72 h1-3.Blank group 24,48,72 h;4-6.Negative conrtol group 24,48,72 h;7-9.RNAi group 24,48,72 h图5 Western bolt检测各时间点细胞蛋白PPARγ表达量Fig.5 The expression of PPARγ detected by Western blot
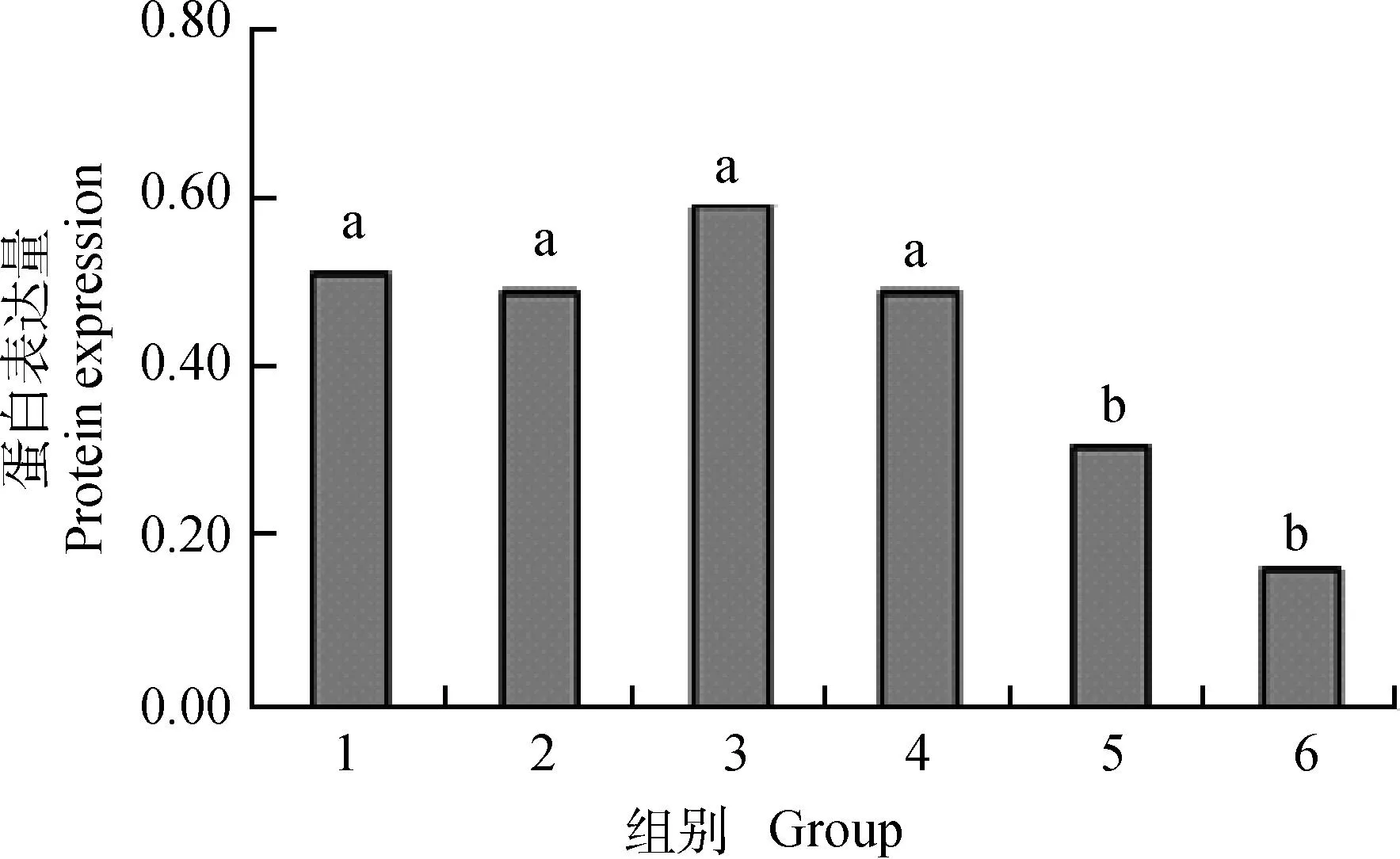
1~3.对照组24、48、72 h蛋白表达量;4~6.干扰组24、48、72 h蛋白表达量1-3.Negative control group protein expression at 24,48,72 h;4-6.RNAi group protein expression at 24,48,72 h图6 PPARγ蛋白表达量相对灰度值Fig.6 The results of the relative density of PPARγ protein expression
3讨论
基因沉默是指通过各种方式,在不影响宿主基因组DNA的情况下使特定基因不表达或是低表达的现象。一种是转录水平的沉默,另一种是转录后水平的沉默。RNAi则属于后者,在进化水平上是高度保守的,由双链RNA诱发的,特异性降解同源mRNA的现象。如果需要长期抑制基因和蛋白的表达,需要shRNA表达载体。目前实验室常用的载体分为两类,一类质粒载体,另一类是病毒载体。质粒载体会随着载体复制扩增,抑制效果可以随细胞传代传递下去,但会受到细胞类型的影响。病毒载体又分为腺病毒载体和慢病毒载体等。腺病毒可以高效干扰多种细胞株,但不能长期稳定的表达shRNA。慢病毒载体是人类免疫缺陷病毒-1(HIV-1)来源的一种病毒载体,目前广泛应用于基因功能的研究及基因沉默生物模型的制备[11-14]。与其他载体相比,其优势在于转染率高且不受细胞周期的影响;可以直接整合到染色体,很少发生炎症和毒性反应,本研究中构建的慢病毒载体pLVX-puro-PPARγ-shRNA转染肌内脂肪细胞后未出现大面积细胞漂浮死亡的现象,载体构建成功,得到细胞稳定株。研究报道,慢病毒载体可以同时兼容多个启动子,抑制多个基因的表达,转染效率可以高达80%~100%,本研究构建的白绒山羊shRNA干扰载体转染效率在80%以上,可以用于后续大规模包装使用。基于以上优点,慢病毒载体技术在RNA干扰中的应用也越来越广泛,逐渐从实验室走向了临床应用。

*.P<0.05.The same as Figure 9图7 PPARγ基因表达量下降后绒山羊肌内脂肪细胞的增殖情况Fig.7 Proliferation of intramuscular adipocytes after the PPARγ gene expression levels decreased

A.阴性对照组;B.干扰组A.Negative control group;B.RNAi group图8 油红O染色肌内脂肪细胞 200×Fig.8 Oil Red O staining result of intramuscular adipocytes 200×
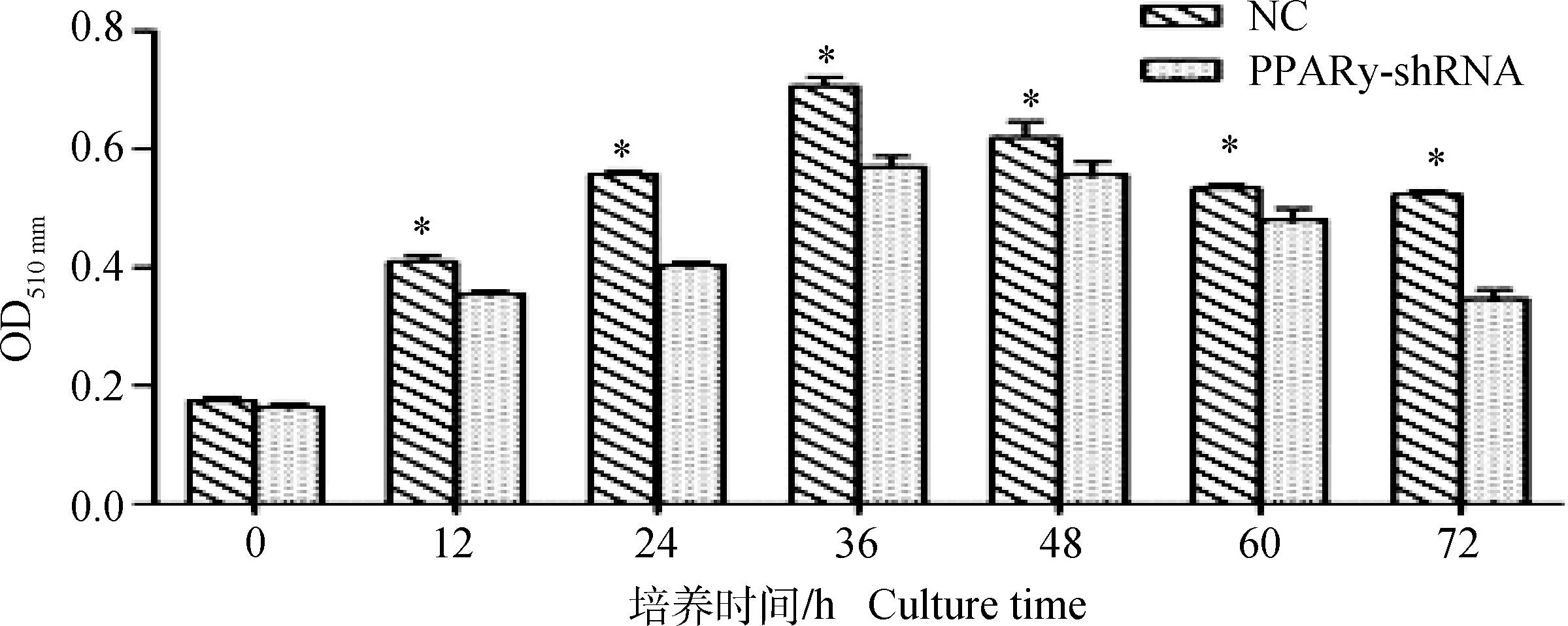
图9 PPARγ基因表达量下降后绒山羊肌内脂肪细胞的分化情况Fig.9 Differentiation of intramuscular adipocytes after the expression of PPARγ gene decreased
脂肪细胞作为糖代谢和脂代谢关键的调节器,是研究肥胖及其多种代谢并发症如糖尿病、高血压的理想模型。从育种角度来看,了解脂肪沉积机理及规律可为提高肌内脂肪含量和改善肉品品质提供理论依据。有研究证明,PPARγ是多功能的转录调控因子,参与脂肪生成和免疫调控等[15]。许多试验证明类固醇类物质可以促进骨髓间质细胞中PPARγ的表达,上调PPARγ基因的表达量可以诱导间质细胞向脂肪分化方向发展,促进脂质的积累,同时降低骨源性细胞的分化[16-18]。P.Tontonoz等[19]研究PPARγ可以诱导成纤维细胞分化为成熟脂肪细胞。另外,PPARγ作为主要的调控因子可正向调节另一转录因子C/EBPa的启动,其蛋白可以调节许多参与糖代谢和脂代谢中关键基因的表达。R.A.Hegele等[20]研究人类PPARγ基因杂合子突变出现机体脂肪分布异常,胰岛素抗性和高血脂等代谢紊乱症。所以本研究通过慢病毒介导高效率转染,构建稳定干扰PPARγ基因的细胞株,进而揭示PPARγ在肌内脂肪细胞中的确切功能,为开拓基因治疗提供了一条崭新途径。慢病毒载体本身不在宿主细胞内增殖,不会导致寄主细胞的死亡,可以根据注入慢病毒载体不同剂量,调节PPARγ的干扰效率。
实时定量和Western blot是目前对于RNAi干扰效果最为便捷的检测方法,分别从基因和蛋白角度对其进行全面分析。因为基因沉默导致的后果是多种多样,RNAi有可能导致 mRNA水平的降低,也有可能是蛋白水平的改变,从而造成表型和功能的变化。本研究在转染目的细胞0~72 h内每隔24 h收集细胞进行Real-time和Western blot检测,在干扰24~72 h后,干扰组PPARγ基因mRNA水平显著低于对照组,干扰效果显著,慢病毒体稳定抑制了基因的表达。有趣的是,在RNAi试验中有时候发现沉默的目的基因在mRNA水平没有发生显著性改变,这就需要进一步在蛋白质水平进行验证,这是因为在干扰过程中siRNA是通过microRNA的作用方式在不影响mRNA表达水平的基础上直接调控蛋白质的表达。本研究中对转染细胞24~72 h进行蛋白检测,在24 h检测PPARγ蛋白在干扰组和对照组中无明显差别,而在48和72 h干扰组中PPARγ蛋白表达量明显降低,说明PPARγ-shRNA慢病毒感染脂肪细胞后能够在蛋白水平抑制PPARγ的表达。通过Real-time检测显著抑制时间点是转染后24 h,而蛋白检测则为转染后48 h,可能是由于PPARγ在绒山羊脂肪细胞中首先表现在mRNA水平的降低,其次是蛋白水平的改变,两者之间有时间上的先后顺序。
MTT试验结果显示,PPARγ的表达量降低后,OD490 nm检测干扰组细胞从36~72 h增殖能力增强,显著高于对照组。OD510 nm检测干扰组细胞从24~72 h分化能力减弱,显著低于对照组,这与染色结果相一致。即PPARγ基因有促进绒山羊肌内前脂肪细胞分化,抑制增殖的作用,这一结果与王丽[21]等人的发现相一致。
4结论
本研究成功构建PPARγ-shRNA慢病毒载体,并将慢病毒载体转染绒山羊肌内脂肪细胞后,筛选得到稳定株。荧光定量和Western blot检测慢病毒载体介导的shRNA 可以有效的降低肌内脂肪细胞PPARγ的表达,发现其mRNA水平和蛋白水平的作用时间相隔24 h。RNAi技术沉默肌内脂肪细胞PPARγ基因后能明显降低脂肪细胞甘油三酯的合成,促进脂肪细胞的增殖。
参考文献(References):
[1]WOOD J D,ENSER M,FISHER A V,et al.Fat deposition,fatty acid composition and meat quality:A review[J].MeatSci,2008,78(4):343-358.
[2]MUELLER E.Understanding the variegation of fat:Novel regulators of adipocyte differentiation and fat tissue biology[J].BiochimBiophyActa,2014,1842(3):352-357.
[3]CRISAFULLI C,CUZZOCREA S.The role of endogenous and exogenous ligands for the peroxisome proliferator-activated receptor alpha (PPAR-alpha) in the regulation of inflammation in macrophages[J].Shock,2009,32(1):62-73.
[4]PYPER S R,VISWAKARMA N,YU S,et al.PPARalpha:energy combustion,hypolipidemia,inflammation and cancer[J].NuclReceptSignal,2010,8:e002.
[5]SHUII S,OLSON P,SEARS D D,et al.PPARγ activation in adipocytes is sufficient for systemic insulin sensitization[J].ProcNatlAcadSciUSA,2009,106(52):22504-22509.
[6]SCHUPP M,LAZAR M A.Endogenous ligands for nuclear receptors:digging deeper[J].JBiolChem,2010,285(52):40409-40415.
[7]MEI C L,HE P,CHENG B,et al.Chlamydia pneumoniae induces macrophage-derived foam cell formation via PPARα and PPARγ-dependent pathways[J].CellBiolInt,2009,33(3):301-308.
[8]MITTAL V.Improving the efficiency of RNA interference in mammals[J].NatRevGenet,2004,5(5):355-365.
[9]LOIS C,HONG E J,PEASE S,et al.Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors[J].Science,2002,295(5556):868-872.
[10]杜琛,付绍印,韩志玲,等.绒山羊肌内前体脂肪细胞的基因表达分析[J].畜牧兽医学报,2013,44(10):1532-1538.
DU C,FU S Y,HAN Z L,et al.Gene expression analysis on intramuscular preadipocytes in the cashmere goats[J].ActaVeterinariaetZootechnicaSinica,2013,44(10):1532-1538.(in Chinese)
[11]MUELLER S,GAUSSON V,VODOVAR N,et al.RNAi-mediated immunity provides strong protection against the negative-strand RNA vesicular stomatitis virus in Drosophila[J].ProcNatlAcadSciUSA, 2010,107(45):19390-19395.
[12]BRACKENY D E,SCOTT J C,SAGAWA F,et al.C6/36 aedes albopictus cells have a dysfunctional antiviral RNA interference response[J].PLoSNeglTropDis,2010,4(10):e856.
[13]LÉGER P,LARA E,JAGLA B,et al.Dicer-2-and Piwi-mediated RNA interference in Rift Valley fever virus-infected mosquito cells[J].JVirol, 2013,87(3):1631-1648.
[14]CHARLEY P A,WILUSZ J.Sponging of cellular proteins by viral RNAs[J].CurrOpinVirol,2014,9:14-18.
[15]KLIEWER S A,MANGELSDORF D J.Fibroblast growth factor 21:from pharmacology to physiology[J].AmJClinNutr,2010,91(1):254S-257S.
[16]AHMADIAN M,DUCAN R E,SUL H S.The skinny on fat:lipolysis and fatty acid utilization in adipocytes[J].TrendsEndocrinolMetab,2009,20(9):424-428.
[17]SUGII S,OLSON P,SEARS D D,et al.PPARgamma activation in adipocytes is sufficient for systemic insulin sensitization[J].ProcNatlAcadSciUSA,2009,106(52):22504-22509.
[18]王义生,王小刚,李月白,等.靶向PPARγ基因RNA干扰对激素抑制家兔骨髓基质细胞成骨分化的影响[J].郑州大学学报(医学版),2009,44(2):275-279.
WANG Y S,WANG X G,LI Y B,et al.Effects of RNAi for target on PPARγ to mainta in osteogenic differentiation of MSCs of rabbit suppressed by steroid[J].JournalofZhengzhouUniversity(MedicalSciences),2009,44(2):275-279.(in Chinese)
[19]TONTONOZ P,HU E,GRACES R A,et al.mPPAR gamma 2:tissue-specific regulator of an adipocyte enhancer[J].GenesDev,1994,8(10):1224-1234.
[20]HEGELE R A,POLLEX R L.Genetic and physiological insights into the metabolic syndrome[J].AmJPhysiolRegulIntegrCompPhysiol,2005,289(3):R663-R669.
[21]王丽,那威,王宇祥,等.鸡PPARγ 基因的表达特性及其对脂肪细胞增殖分化的影响[J].遗传,2012,34(5):454-464.
WANG L,NA W,WANG Y X,et al.Characterization of chickenPPARγ expression and its impact on adipocyte proliferation and differentiation[J].Hereditas(Beijing),2012,34(5):454-464.(in Chinese)
(编辑郭云雁)
Construction of Lentiviral RNAi Vector of PPARγ Gene in Cashmere Goat and Its Effect on Proliferation and Differentiation of Intramuscular Adipocytes
DU Chen1,LI Jin-quan2*,CHEN Xiu-juan1*
(1.DepartmentofObstetricsandGynecology,AffiliatedHospital,InnerMongoliaMedicalUniversity,Hohhot010050,China;2.KeyLaboratoryofAnimalGenetics,BreedingandReproductionofInnerMongoliaAutonomousRegion,CollegeofAnimalScience,InnerMongoliaAgriculturalUniversity,Hohhot010018,China)
Abstract:The cashmere-goat intramuscular adipocytes were used as experimental materials,and aimed to establish the stable intramuscular adipocyte line for the interference through target interrupting the peroxisome proliferation-activated receptor(PPARγ) gene,so as to explore the function of cashmere-goatPPARγgene during the proliferation and differentiation processes of the intramuscular adipocytes.Lentivirural plasmids pack-aging system was adopted to establish a lentiviral vector for RNA interference of specific targeting White Cashmere-goatPPARγgene,which was used to establish the intramuscular adipocyte line to stabilize and silence thePPARγgene.Real-time quantitative and Western blot methods were used to detect the expression ofPPARγgene at different points in time in the interference group and control group,as well as the MTT and Oil Red O Staining methods were used to research the influence of cashmere-goatPPARγgene on the proliferation and differentiation of intramuscular adipocytes.It was confirmed by sequencing that oligonucleotide chains containing PPARγ-shRNA lentivirus vector were inserted properly,its infection efficiency to intramuscular adipocytes was above 80%,lentivirus-mediated shRNA detected by fluorescence quantitative and Western blot could effectively reduce the expression ofPPARγgene,the reaction time interval between mRNA and protein level was 24 h.After silencing ofPPARγgene,the concentration of triglyceride within adipocytes was significantly lower than that in the control group;Contrarily,the cell proliferation ability in MTT test interference group was higher than that in control group.This study established cashmere-goat shRNA-PPARγ lentivirus interference vector,after successfully transfecting into the intramuscular adipocytes,it obviously inhibited the differentiation of adipocytes,and promoted the proliferation of intramuscular adipocytes,which lay foundation for further research the role ofPPARγgene in cashmere-goat metabolic pathways and in the fat deposition mechanism.
Key words:intramuscular adipocyte;RNA interference;PPARγ
doi:10.11843/j.issn.0366-6964.2016.04.005
收稿日期:2015-05-23
基金项目:国家自然科学基金(30960246);内蒙古自然科学基金(2010Zd11);国家科技支撑项目(2011BAD28B05);国家绒毛用羊现代农业产业技术体系(CARS-40-05)
作者简介:杜琛(1986-),女,内蒙古呼和浩特人,博士,助理研究员,主要从事分子遗传研究,E-mail:duchen198607@126.com *通信作者:李金泉,博导,教授,主要从事动物遗传学研究,E-mail:lijinquan_nd@126.com;陈秀娟,硕导,教授,主要从事妇产科学研究,E-mail:90098687@sina.com
中图分类号:S827;S813.3
文献标志码:A
文章编号:0366-6964(2016)04-0671-08
