
玉米陈化关键酶基因Zmlox-2的RNA干扰载体的构建
2016-06-17 07:23韩颖赵寿经杨瑜孙尧王乐
华南理工大学学报(自然科学版) 2016年3期
韩颖 赵寿经 杨瑜 孙尧 王乐
(1.吉林大学 生物与农业工程学院, 吉林 长春130022; 2.长春工业大学 化学与生命科学学院, 吉林 长春130012)
玉米陈化关键酶基因Zmlox-2的RNA干扰载体的构建
韩颖1赵寿经1杨瑜1孙尧2王乐1
(1.吉林大学 生物与农业工程学院, 吉林 长春130022; 2.长春工业大学 化学与生命科学学院, 吉林 长春130012)
摘要:脂肪酸氧化酶(LOX)催化不饱和脂肪酸过氧化反应,导致玉米(Zea mays L.)种子储藏品质的下降.为抑制玉米陈化关键酶基因Zmlox-2的表达,依据RNA干扰原理,针对玉米Zmlox-2基因进行克隆,采用重组PCR及酶切连接技术成功构建了Zmlox-2基因的RNA干扰植物表达载体,并将重组载体pRI101-on-Zmlox-2-RNAi通过热激转化法转化到根癌农杆菌中.实验结果表明Zmlox-2基因的RNA干扰表达载体构建正确,这为下一步诱导遗传转化、探索陈化酶基因的干扰表达效果奠定了理论和实验基础.
关键词:生物工程;陈化;RNA干扰表达载体;重组PCR;脂肪酸氧化酶
玉米是脂质含量较高的粮种,其胚芽中的油脂约占总重的3.1%~7.0%.近年来的研究表明,玉米贮藏期间脂质的降解是导致其品质下降并产生令人不快气味的主要原因之一.脂肪酸氧化酶(LOX,EC 1.13.11.12)是脂质降解的关键酶,其催化多元不饱和脂肪酸过氧化反应,然后经过裂解酶分解形成短链的丙二醛(MDA)等挥发性物质,导致玉米储藏过程中品质下降[1- 4].脂肪酸氧化酶是一个多基因家族,玉米种子中共发现3种LOX酶,其中LOX-2酶可能是导致储藏品质下降的关键酶.因此从理论上讲,玉米LOX-2酶的缺失可以明显地阻止脂质过氧化作用,减缓贮藏玉米氧化变质的速度,保持清新气味,提高耐贮性.上述推测己经在玉米脂肪酸氧化酶LOX-2 (玉米种子中主要的LOX同工酶)缺失体与耐贮性关系的研究中得到了初步证实[5-7].针对玉米、水稻、大豆等植物种子,通过外源性减少或缺失LOX酶活性以提高储藏品质,已经做了一些研究工作,例如微波辐射、低温气调储存等[8].但是这些方法消耗大量的物质资源和能量,还导致种子蛋白质的变性.因此通过RNA干扰技术(RNAi)抑制玉米lox-2基因的表达量有可能成为改善玉米储藏品质、延缓种子陈化衰老的有效手段[9].
本研究利用PCR重组技术,克隆玉米lox-2基因(Zmlox-2)的特异序列,构建成含lox基因反向重复结构(正反义片段之间间隔一段含有多克隆位点的序列,便利转录后形成具有“发夹结构”的dsRNA)的RNA干扰表达载体(pRI101-on-Zmlox-2-RNAi),将其转导入根癌农杆菌中,获得RNAi表达载体基因工程菌.Zmlox-2基因的RNAi表达载体的构建为后续的遗传转化及探讨Zmlox-2基因的沉默表达对延缓玉米陈化的作用提供了研究素材,奠定了理论和实验基础.
1材料
1.1植物材料
玉米(ZeamaysL.)自交系种子WM03由长春农业科学研究所提供.
1.2菌株、质粒、引物
植物转化高效表达载体pRI101-on购自宝生物工程大连有限公司;根癌农杆菌(Agrobacteriumtumefaciens)LBA 4404菌株、感受态大肠杆菌(Escherichiacoli)JM109菌株均为吉林大学生物与农业工程学院生物工程实验室保存;引物由宝生物工程大连有限公司合成.
1.3酶和试剂
RNA提取试剂盒、反转录试剂盒、质粒提取试剂盒、凝胶回收试剂盒、T4 DNA连接酶、PrimeSTAR HS DNA聚合酶、DNA电泳标准物等均购自宝生物工程大连有限公司,卡那霉素购自北京鼎国生物技术有限公司,其余为国产分析纯.
2方法
2.1玉米总RNA提取
取玉米种子(WM03)2 g放入研钵中,加入液氮研磨.研磨后的样品中加入20 mL RNA 提取试剂,提取总RNA,并以此为模板反转录成cDNA.
2.2引物设计
根据Genebank上获得的Zmlox-2(ABC59686)、Zmlox-1(DAA56391)、Zmlox-3(NP_001105515)基因序列,通过Clustal软件,找出Zmlox-2、Zmlox-1和Zmlox-3编码区中的非同源序列区,并在此区域内设计引物.目的使RNA干扰元件特异性抑制Zmlox-2的表达,不能抑制Zmlox-1和Zmlox-3的表达.使用Premier 5.0设计特异目的基因序列的正向和反向引物,所设计的引物含载体构建所需的酶切位点NdeⅠ和KpnⅠ.引物由宝生物工程大连有限公司合成,用于后续的载体构建[10-11].
设计的引物核苷酸序列为:
正义片段5′ATCGTCGTCCACGACATATGCATGTTCGGAAACATCGGAAA3′和5′GAATTCGAGCTCG-GGCCCGTCGACTACTTGCTCGGATCAAGG3’.
反义片段5′GTCGACGGGCCCGAGCTCGAATTCTACTTGCTCGGATCAAGG3′和5′GACGCCCTTA-TTTTAGGTACCATGTTCGGAAACATCGGAAA3′.
使用正义片段和反义片段拼接上游引物5′ATCGTCGTCCACGACATATGC3′(带有NdeⅠ酶切位点),下游引物5′GACGCCCTTATTTTAGGTACC3′(带有KpnⅠ酶切位点).
2.3融合Zmlox-2 的RNA干扰元件的扩增
以反转录的玉米cDNA为模板,以正义片段引物和反义片段引物分别为反应引物,按下面的反应体系和扩增程序进行PCR扩增反应,进行正义片段和反义片段的扩增[12-14].之后以正义片段和反义片段为扩增模板,利用带酶切位点的拼接引物PCR扩增Zmlox-2 的RNA干扰元件
正义片段扩增反应体系:5×PrimeSTAR PCR 缓冲液10 μL,dNTP 混合物(2.5 mmol/L) 4 μL,正义片段上游引物1 μL,正义片段下游引物1 μL,PrimeSTAR HS DNA聚合酶(2.5 U/μL) 2 μL,cDNA模板2 μL,ddH2O 30 μL,总体系50 μL.PCR仪扩增程序为:94℃,3 min;94℃ 30 s,62-1℃ 45 s, 72℃ 2 min,5次循环;94℃ 30 s,57℃ 45 s,72℃ 2 min,30次循环;72℃ 10 min.
反义片段扩增反应体系:5×PrimeSTAR PCR 缓冲液10 μL,dNTP混合物 (2.5mmol/L) 4 μL,反义片段上游引物1 μL,反义片段下游引物1 μL,PrimeSTAR HS DNA聚合酶(2.5 U/μL) 2 μL,cDNA模板2 μL,ddH2O 30 μL,总体系50 μL.PCR 仪扩增程序为:94℃,3 min;94℃ 30 s,65-1℃ 45 s,72℃ 2 min,5次循环;94℃ 30 s,60℃ 45 s,72℃ 2 min,30次循环;72℃ 10 min.
正义片段、反义片段拼接扩增反应体系:5×PrimeSTAR PCR 缓冲液10 μL,dNTP 混合物(2.5 mmol/L) 4 μL,上游拼接片段引物1 μL,下游拼接片段引物1 μL,PrimeSTAR HS DNA聚合酶(2.5 U/μL) 2 μL,正义片段和反义片段混合模板2 μL,ddH2O 30 μL,总体系50 μL.PCR仪扩增程序为:94℃,3 min ;94℃ 30 s,70-1℃ 45 s, 72℃ 2 min,5 次循环;94℃ 30 s,65℃ 45 s,72℃ 2 min,30 次循环;72℃ 10 min.
重组PCR扩增得到Zmlox-2 RNA干扰元件,图1为其构建示意图.用凝胶回收试剂盒切胶回收目的产物.

图1 Zmlox-2 RNAi元件Fig.1 Zmlox-2 RNAi component
2.4融合Zmlox-2的RNA干扰载体的构建
将扩增得到的Zmlox-2 RNA干扰元件插入单子叶植物转化表达载体pRI101-on(构建原理如图2所示).pRI101-on载体长度约10.4 kb,含有一个组成型启动子(CaMV 35S)和外来基因表达增强子(OsADH 5′UTR),增强子的下游即为外源基因插入的多克隆位点.pRI101-on载体上有NPT Ⅲ卡那霉素抗性基因,可用于筛选重组质粒的标记.
利用限制酶NdeⅠ和KpnⅠ分别对Zmlox-2 基因RNAi元件和植物表达载体pRI101-on双酶切,之后利用T4 DNA连接酶连接,连接后的产物即为Zmlox-2基因的RNAi重组载体.采用热激转化法[15]将其直接转化至大肠杆菌E.coliJM109感受态细胞中,涂布含50 mg/L卡那霉素的LB平板过夜培养,挑选单菌落进行菌落PCR初步鉴定.阳性菌落继续在LB液体培养基中进行培养,提取重组质粒进行双酶切鉴定.连接成功的重组载体命名为pRI101-on-Zmlox-2-RNAi[16-17].
2.5融合Zmlox-2的RNA干扰载体的转化
将重组质粒pRI101-on-Zmlox-2-RNAi利用热激转化法[15]转化至根癌农杆菌LBA4404感受态细胞中.涂布YEP平板过夜培养菌体,以菌落为模板进行PCR鉴定,将含有Zmlox-2基因RNAi重组质粒的阳性克隆菌命名为LBA4404-pRI101-on-Zmlox-2-RNAi工程菌,将其加入15%的甘油后放-70℃冰箱保存[18].
3结果与分析
3.1玉米总RNA提取
将提取的玉米总RNA在1%的琼脂糖凝胶上进行电泳,电泳结果如图3所示.由电泳图谱可见:28S rRNA和18S rRNA条带完整清晰,28S/18S亮度比大约为2∶1,因此所提取的RNA是比较完整的,没有蛋白污染,适用于反转录合成cDNA.
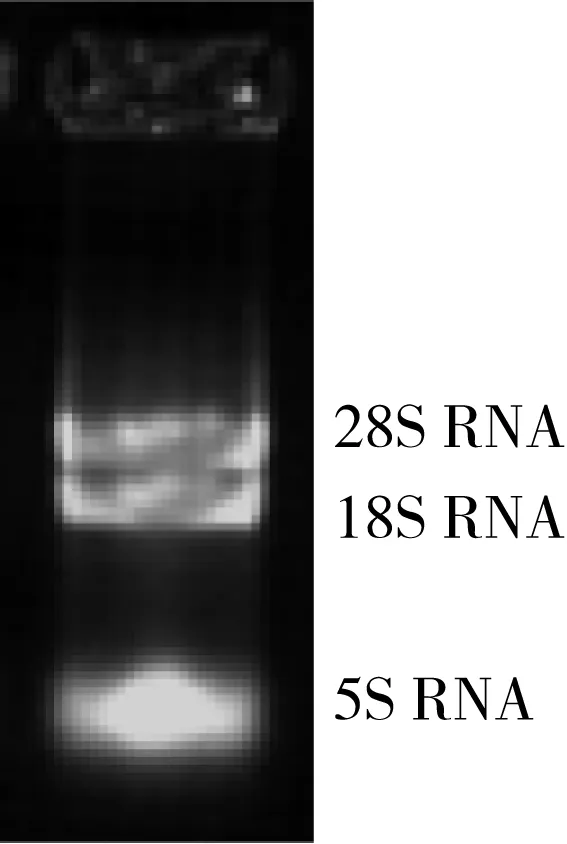
图3 玉米总RNA电泳图Fig.3 Electrophoregram of total RNA of maize
3.2融合Zmlox-2 的RNA干扰元件的扩增
通过玉米Zmlox-2、Zmlox-1和Zmlox-3的基因序列比对,从起始密码子开始在相似度低的非同源编码区(约500 bp左右)设计引物,如图4所示.使得构建的干扰元件能够特异性地干扰Zmlox-2的表达,不能抑制Zmlox-1和Zmlox-3的表达.

图4 Zmlox-2、Zmlox-1和Zmlox-3的非同源编码区比对Fig.4 Alignment analysis of nonhomologous coding region of Zmlox-2、Zmlox-1 and Zmlox-3氨基酸序列号:Zmlox-1 (DAA56391),Zmlox-2 (ABC59686),Zmlox-3 (NP_001105515)
然后以玉米总RNA反转录的cDNA产物为模板,以正义片段引物和反义片段引物进行第1轮PCR反应,分别构建正义片段和反义片段.PCR反应程序的设置直接影响着引物和模板的特异性结合效果.退火温度低会产生一些非特异性产物,退火温度高会干扰引物和模板特异性结合.本实验对退火温度、循环数等进行实验条件优化,确定了最终的反应程序.PCR产物经琼脂糖凝胶电泳鉴定,结果显示:在目的条带440 bp处各有一清晰条带(见图5).

图5 正义片段和反义片段PCR扩增电泳图谱Fig.5 Electrophoregram of PCR amplification of sense fragment and antisense fragmentM—DNA 标准物DL2000;1—正义片段PCR产物;2—反义片段PCR产物
以第1轮正义片段和反义片段的PCR产物为模板,利用带酶切位点的拼接引物进行第2轮PCR扩增.优化PCR扩增实验条件后的产物经琼脂糖凝胶电泳鉴定.结果显示:目的产物在860 bp处条带清晰(见图6).结果与预期相同,证明成功地构建出Zmlox-2-RNA干扰元件,即Zmlox-2-RNAi.

图6 Zmlox-2 RNA干扰元件PCR产物电泳图谱Fig.6 Electrophoregram of PCR product of Zmlox-2-RNAi componentM—DNA 标准物 DL2000;1—RNAi干扰元件
3.3融合Zmlox-2的RNA干扰载体的构建
对上述重组PCR扩增得到的产物Zmlox-2-RNAi元件和植物表达载体pRI101-on同时用NdeI和KpnI双酶切,使用T4 DNA连接酶连接干扰元件和表达载体pRI101-on,得到重组干扰载体pRI101-on-Zmlox-2-RNAi.然后热转化至大肠杆菌JM109感受态细胞中,涂布平板过夜培养菌体.挑选单菌落,分别进行摇瓶培养.提取RNAi载体质粒,用NdeI、KpnI对其进行酶切鉴定.由电泳图谱可见:2泳道得到860 bp左右的目的条带(见图7).产物经核苷酸序列测序,结果与预期相同,证明Zmlox-2基因的RNAi载体构建成功.

图7 pRI101-on-Zmlox-2-RNAi重组质粒的酶切鉴定电泳图谱Fig.7 Electrophoregram of double enzyme digestion of pRI101-on-Zmlox-2-RNAi recombinant plasmidM—DNA 标准物 DL2000; 1—pRI101-on;2—pRI101-on-Zmlox-2-RNAi重组质粒酶切产物
3.4融合Zmlox-2的RNA干扰载体的转化
采用热激转化法将pRI101-on-Zmlox-2-RNAi重组质粒转化至根癌农杆菌LBA4404细胞中,得到Zmlox-2基因RNAi表达载体工程菌.涂布于含有50 mg/L卡那霉素的YEB平板过夜培养菌体,挑取单菌落进行菌落PCR鉴定(正义片段引物),结果见图8.从图中可以看到, 2-5泳道中的条带位置与预期位置一致.PCR鉴定结果表明,获得了LBA4404-pRI101-on-Zmlox-2-RNAi植物表达载体工程菌.
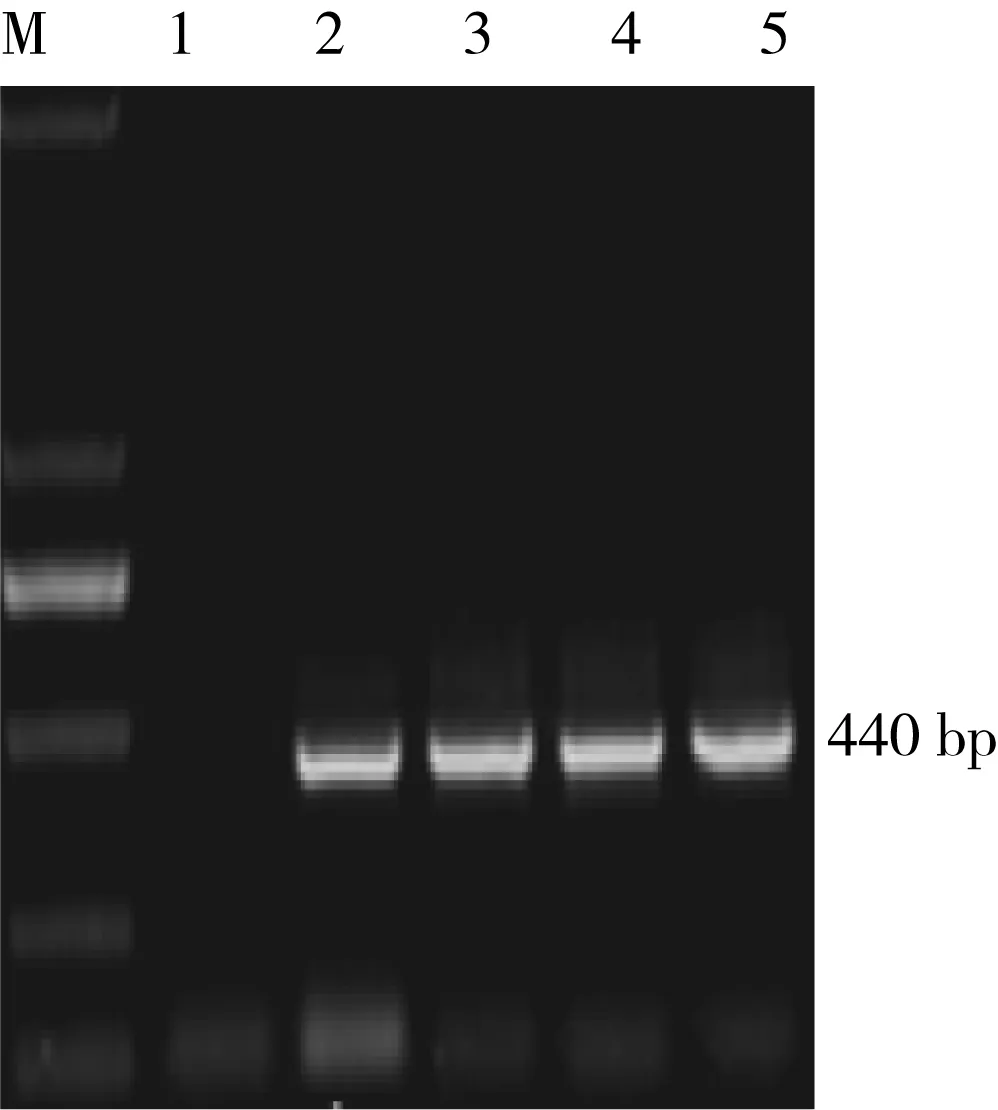
图8 菌落PCR鉴定电泳图Fig.8 Electrophoregram of bacterial colony PCR identificationM—DNA 标准物 DL2000;1—阴性对照;2-5—LBA4404-pRI101-on-Zmlox-2-RNAi菌落PCR产物
4讨论
RNAi是由双链RNA(dsRNA)诱导同源mRNA高效特异性降解致使转录后基因沉默的现象.RNAi作为一种新的技术方法,在植物基因组功能鉴定、品种改良以及植物抗病性研究等方面展现出极大的优势[19].
RNAi表达载体的构建过程中,正义片段与反义片段的连接较为困难.本研究采用重组PCR技术克服传统限制性内切酶连接基因片段的缺点,在引物设计时加入相应的酶切位点序列,再与传统的酶切连接法相结合,成功地构建了Zmlox-2基因的RNAi表达载体.
引物的设计是重组PCR技术中最重要的环节,直接影响最终的PCR结果.研究中须保证不影响玉米基因组功能的前提下降解陈化关键酶基因,如果在玉米基因组内有某个基因与RNAi干扰元件具有同源性,这个基因可能会受到影响.在实验中笔者选取的Zmlox-2目的基因片段与玉米种子Zmlox-1、Zmlox-3基因不存在30个碱基以上的同源序列,保证了RNA干扰的特异性.本研究通过非同源编码区比对设计引物,构建Zmlox-2基因的干扰载体,使Zmlox-2基因沉默的同时,不影响其他玉米LOX酶行使功能.
5结论
文中采用重组PCR技术及酶切连接技术构建了融合Zmlox-2基因的干扰表达载体,并将重组载体pRI101-on-Zmlox-2-RNAi转化到根癌农杆菌中,验证了所构建载体的正确性.实验操作简便快捷,为植物RNA干扰表达载体的构建提供了一个高效、便捷、适应性广的方法.
参考文献:
[1]胡廷章,胡宗利,屈霄霄,等.植物脂肪氧化酶的研究进展 [J].生物工程学报,2009,25(1):1-9.
HUTing-zhang,HUZong-li,QUXiao-xiao,etal.Advancesinplantlipoxygenasesresearch[J].ChineseJournalofBiotechnology,2009,25(1):1-9.
[2]WANG Fa-wei,WANG Rong,JING Wen,et al.Quantitative dissection of lipid degradation in rice seeds during accelerated aging [J].Plant Growth Regulation,2012,66(1):49-58.
[3]GOMBOEVA S B,SHUMAEV K B,GESSLER N N,et al.The mechanism of oxidation ofβ-carotene and polyunsaturated fatty acids [J]. Doklady Biochemistry and Biophy-sics,2001,377(1):98-101.
[4]周显青,张玉荣,张勇.储藏玉米陈化机理及挥发物与品质变化的关系[J].农业工程学报,2008,24(7):242-246.ZHOU Xian-qing,ZHANG Yu-rong,ZHANG Yong.Aging mechanism of stored maize and the relationship between its volatile components and quality [J].Transactions of the Chinese Society of Agricultural Engineering,2008,24(7):242-246.
[5]CHRISTOPHE Bailly,ABDELILAH Benamar,FRANCOISE Corbineau,et al.Changes in malondialdehyde content and in superoxide dismutase catalase and glutathione reductase activities in sunflower seeds as related to dete-rioration during accelerated aging [J]. Physiologia Plantarum,1996,97(1):104-110.
[6]STEWART R R,BEWLEY J D.Lipid peroxidation asso-ciated with accelerated aging of soybean axes [J].Plant Physiology,1980,65(2):245-248.
[7]李金库,王玉娟,张瑛,等.玉米脂肪氧化酶缺失种质耐储藏特性的研究 [J].中国粮油学报,2006,21(6):143-146.
LI Jin-ku,WANG Yu-juan,ZHANG Ying,et al. Lipoxygenase deficiency vs grain storage durability for maize [J].Journal of the Chinese Cereals and Oils Association,2006,21(6):143-146.
[8]曲春阳,刘鹏,屠康.大米储藏保鲜技术现状及研究进展 [J].粮食储藏,2009,38(3):22-25.
QU Chun-yang,LIU Peng,TU Kang.Research progress and present situation on rice storage technology [J]. Grain Storage,2009,38(3):22-25.
[9]赵寿经,侯春喜,徐立新,等.抑制齐墩果烷型人参皂苷合成支路对达玛烷型人参皂苷生产能力的影响 [J].吉林大学学报(工学版),2011,41(3):865-868.
ZHAO Shou-jing,HOU Chun-xi,XU Li-xin,et al.Effects of suppressing oleanane-type ginsenoside biosynthesis on dammarane-type ginsenoside production [J].Journal of Jilin University(Engineering and Technology Edition),2011,41(3):865-868.
[10]马建,厉志,刘艺苓,等.应用重组PCR技术构建大豆脂肪氧化酶基因RNA干扰表达载体 [J].大豆科学,2008,27(4):564-568.
MA Jian,LI Zhi,LIU Yi-ling,et al.Use recombiant PCR to construct RNAi expression vector of soyabean lipoxygenase [J].Soyabean Science,2008,27(4):564-568.
[11]吕品,柴晓杰,王丕武,等.大豆胰蛋白酶抑制剂KSTI3基因的克隆及其植物表达载体的构建[J].吉林农业大学学报,2007,29(3):275-278.
Lü Pin,CHAI Xiao-jie,WANG Pi-wu,et al.Cloning and construction of expression vector on KSTI3 gene of soybean Kunitz-type trypsin inhibitor [J].Journal of Jilin Agricultural University,2007,29(3):275-278.
[12]HAN J Y,KWON Y S,YANG D C,et al.Expression and RNA interference-induced silencing of the dammarenediol synthase gene in Panax ginseng [J].Plant Cell Physiology,2006,47(12):1653-1662.
[13]SHIRASAWA K,TAKEUCHI Y,SUZUKI Y.Identification of gene for rice (Oryzasativa) seed lipoxygenase-3 involved in the generation of stale flavor and development of SNP markers for lipoxygenase-3 deficiency [J].Breed Science,2008,58(2):169-176.
[14]SUN Yao,ZHAO Shou-jing,LIANG Yan-long,et al.Re-gulation and differential expression of protopanaxadiol synthase in Asian and American ginseng ginsenoside biosynthesis by RNA interferences [J].Plant Growth Re-gulation,2013,71(3):207-217.
[15]崔武,刘炜,吴光耀.高效、快速地将外源DNA导人根癌土壤杆菌[J].生物工程学报,1995,11(4):350-355.CUI Wu,LIU Wei,WU Guang-yao.Rapid,high effieient transformation of foreign DNA toAgrobacteriumtumefaciens[J].Chinese Journal of Biotechnology,1995,11(4):350-355.
[16]柴晓杰,王丕武,关淑艳,等.应用RNA干扰技术降低玉米支链淀粉含量 [J].植物生理与分子生物学学报,2005,31(6):625-630.
CHAI Xiao-jie,WANG Pi-wu,GUAN Shu-yan,et al.Reducing the maize amylopectin content through RNA interference manipulation [J].Journal of Plant Physiology and Molecular Biology,2005,31(6):625-630.
[17]赵寿经,曹豪杰,何沐阳,等.人参β-AS基因RNAi表达载体工程菌的构建 [J].吉林大学学报(工学版),2012,42(6):865-868.
ZHAO Shou-jing,CAO Hao-jie,HE Mu-yang,et al.Engineering bacteria contruction of ginsengβ-AS gene RNA interference plant expression vector [J].Journal of Jilin University(Engineering and Technology Edition),2012,42(6):865-868.
[18]朱见明,严学兵,史莹华,等.紫花苜蓿光敏色素B基因片段克隆及RNA干扰表达载体的构建 [J].作物学报,2011,37(2):374-379.
ZHU Jian-ming,YAN Xue-bing,SHI Ying-hua,et al.Cloning of Medicago sativa phychrome B cDNA and establishment of its RNA interference expression vector [J].Acta Agronomica Sinica,2011,37(2):374-379.
[19]黄冰艳,吉万全,郭蔼光,等.转录后基因沉默(PTGS)及其在作物遗传改良中的应用 [J].中国生物工程杂志,2005,25(5):1-5.
HUANG Bing-yan,JI Wan-quan,GUO Ai-guang,et al.Post-transcriptional gene silencing (PTGS) and its application to crop genetic improvement [J].China Biotechnology,2005,25(5):1-5.
Construction of RNA Interference Expression Vector Containing Key Aging EnzymeZmlox-2 in Zea Mays
HANYing1ZHAOShou-jing1YANGYu1SUNYao2WANGLe1
(1.College of Biological and Agricultural Engineering, Jilin University, Changchun 130022, Jilin, China;2.College of Chemistry and Life Science, Changchun University of Technology, Changchun 130012, Jilin, China)
Abstract:The peroxidation of unsaturated fatty acid caused by lipoxygenase (LOX) is known to be responsible for the deterioration of storage quality in maize(Zea mays L.) seed.In order to inhibit the expression of key aging enzyme Zmlox-2 gene in maize, Zmlox-2 gene in maize seed was cloned according to the principle of RNA interference (RNAi), and the RNAi plant expression vector containing Zmlox-2 was constructed by means of recombinant PCR and enzyme connection technology.Then, the recombinant vector pRI101-on-Zmlox-2-RNAi was transformed into Agrobacterium tumefaciens via heat shock method.Experimental results show that the RNAi expression vector containing Zmlox-2 gene has been successfully constructed.This research lays a theoretical and experimental foundation for the induction of genetic transformation and the exploration of interference expression effect of aging enzyme gene.
Key words:bioengineering; aging; RNA interference expression vector; recombinant PCR; lipoxygenase
收稿日期:2015- 04-15
*基金项目:吉林省科技发展计划项目(20130102041JC);吉林省教育厅科学技术研究基金资助项目(吉教科合字[2012]376)Foundation item: Supported by the Jilin Scientific and Technological Development Program(20130102041JC)
作者简介:韩颖(1971-),女,博士生,副教授,主要从事农业生物环境与能源工程研究.E-mail:jlgshy@163.com † 通信作者: 赵寿经(1961-),男,教授,主要从事农业生物环境与能源工程研究.E-mail:swgc@jlu.edu.cn.com
中图分类号:Q81
doi:10.3969/j.issn.1000-565X.2016.03.019
猜你喜欢
农家参谋(2021年5期)2021-06-20
中国茶叶加工(2020年3期)2020-10-21
食品安全导刊(2018年24期)2018-01-17
课程教育研究·中(2016年11期)2017-01-04
大学教育(2016年11期)2016-11-16
大学教育(2016年11期)2016-11-16
考试周刊(2016年84期)2016-11-11
大学教育(2016年9期)2016-10-09
中国烟草学报(2012年4期)2012-04-09
