
太白山森林群落和林下草本物种变化的环境解释
2016-06-17 05:47:02许金石张丽霞郭垚鑫柴永福张晨光
西北植物学报 2016年4期
陈 煜,许金石,张丽霞,郭垚鑫,柴永福,王 茂,张晨光,岳 明
(西北大学 生命科学学院,西安 710069)
太白山森林群落和林下草本物种变化的环境解释
陈煜,许金石,张丽霞,郭垚鑫,柴永福,王茂,张晨光,岳明*
(西北大学 生命科学学院,西安 710069)
摘要:采用典范对应分析(CCA)、Pearson相关分析、冗余分析(RDA)分别研究了不同环境因子对太白山森林群落及林下草本物种组成、多样性、功能性状等方面的影响。结果表明:(1)9种环境因子对于群落及林下草本物种的组成变化的解释程度不高,分别为23.9%和21.1%,但与海拔和土壤含水量紧密相关的CCA 1轴能大致区分高、中、低海拔样方的群落所有物种和草本植物的组成差异。(2)沿海拔梯度,群落所有物种的α多样性先增加后降低,林下草本物种α多样性沿海拔梯度有下降趋势,但二者都与坡度(Slope)及木本层盖度(WCD)无关。(3)海拔是决定森林群落及林下草本层物种功能性状值变化的主要因子,与海拔密切相关的RDA1轴能解释森林群落和林下草本物种性状值变化的71.2%和54.7%,林下草本植物与群落的最大株高(Hmax)变化模式不一致。
关键词:林下草本;物种组成;多样性;功能性状;典范对应分析(CCA);冗余分析(RDA)
森林群落的物种多样性及维持机制是当今生态学领域的研究热点。林下草本层也是森林群落不可或缺的一部分[1]。草本植物虽然生物量所占整个森林群落的比重不高,但其物种多样性却是森林生态系统中最高的,并有着最高的物种丰富度[2]。在温带森林中,草本植物可占所有维管植物多样性的90%[3-4]。与群落中的个体较大的木本植物不同,草本植物对于光、热和营养元素水平等环境因素反应展示出不同于大个体树木的多样性特点[5-6]。在物种的结构和组成、物候周期、生活史对策、生物量更新速度、与环境相互作用强度、面对特定干扰的响应方式等方面,林下草本植物也都有其自身的特点[7-10]。前人的研究表明,林下草本植物在增加物种多样性、涵养水源、保持水土、矿山复垦、改变林区小气候等方面具有重要的生态功能[11-12]。温带森林下的草本层对于森林生态系统的能量流动和养分循环也有重要的作用[13],它能够增加土层中的营养元素和有机质,维持和提高土壤肥力[14]。此外,林下草本植物对于乔木幼苗的更新也具有重要意义,在一定程度上影响种苗的发育、存活和生长[4,15],从而可以促进群落的演替[16]。森林下层植物还具有拦截和过滤地表径流的作用,能够有效减少林地水土流失[17]。这些研究表明,林下草本植物在森林群落中具有相当重要的生态学意义[18]。
不过,目前对森林群落的研究往往仍然侧重于群落中的木本植物,对林下草本植物的研究还不是很充分。有关林下草本植物在整个森林生态系统物种多样性的维持中所起的作用和贡献程度的系统研究仍没有充分开展[19]。目前,对于林下草本植物,研究比较多的是生境与草本物种组成的关系。利用典范对应分析(CCA)等排序手段,前人对此已有了初步的研究[20-21]。然而,在木本植物群落构建中经常用到的多样性和功能性状的梯度分析,在林下草本植物中却鲜有研究报道。分析地形、土壤养分,甚至林冠郁闭度等生物和非生物的因素对草本层物种分布、多样性和功能性状变化的作用,定量地揭示它们之间的相互作用,无疑具有重要的生态学意义[22]。本研究基于太白山地区林下草本及群落所有物种的群落学信息以及叶面积(LA)、比叶面积(SLA)、叶片干物质含量(LDMC)、最大株高(Hmax)等常用的功能性状的信息,结合获取的环境因子,在不同梯度下对林下草本植物及群落内所有植物物种的组成、物种多样性及功能性状变化进行了分析和比较,试图通过更多方面阐述森林植被群落的发展状况,在对森林群落物种共存机制的验证和完善,以及不同生活型植物间经济谱和分布模式的对比方面都有十分重要的意义。
1研究区域和方法
1.1研究区域概况
研究区域位于秦岭主峰太白山北麓。秦岭位于中国东部湿润平原以西,青藏高原以东,是暖温带和亚热带的天然分界线,也是南北气候的交接过渡区域,由此成为华北、华中、横断山脉植物区系的交汇点[23],是中国北方植物种类最丰富的地区[24]。秦岭主峰太白山位于陕西省境内,地理位置约为N33°40′~34°10′,E107°19′~107°50′,最高处拔仙台海拔3 767 m,为暖温带第一高峰。太白山地区多年平均降水量约1 000 mm,年均温1~5 ℃[25]。种子植物有125科657属1 782种[26],其中草本植物1 094种,隶属69科 410 属[27]。太白山山体高大,水热、土壤条件随海拔升高呈现有规律的变化。山体北麓属暖温带气候,上部寒冷、雾大、多风。北坡自下而上分别为暖温带、温带、寒温带、亚寒带、寒带气候类型,且土壤垂直分布明显,自下而上依次为褐色土、棕色森林土、高山草甸土、高山石质土等[26]。生境的垂直变化导致植被景观也表现出明显的垂直带状分布,植被类型复杂[21]。区域内有各级保护植物、濒危或渐危物种30余种,本研究涉及到的也有独叶草(Kindoniauniflora)、秦岭米面翁(Buckleyagraebneriana)、陕西报春(Primulahandeliana)、反萼银莲花(Anemonereflexa)等数种。
1.2取样方法和样品处理
在2014年6月间,沿蒿坪寺-小文公一线进行采样,选取人为干扰小、林相较好的森林、灌丛进行样方取样。由于在本区相近的海拔范围内相似的小生境有着相似的群落,为尽可能地扩展环境梯度,相邻样方间地理距离至少为100 m,且保证坡向和坡度不一致。在1 113 m~3 513 m的海拔范围内共设置了41个样方,样方间最大海拔差异为2 400 m。对于木本植物,设置的样方大小为20 m×30 m,记录样方的经纬度、海拔、草本层盖度、坡度和坡向。对样方中出现的木本物种进行每木调查,记录每一株出现在样方中木本植物的学名,用测高仪和皮尺测量每株木本植物的株高、冠幅,进而得出样方中每一种木本植物的多度、平均株高、最大株高(Hmax)。每一种木本植物在样方中的盖度用冠幅根据椭圆面积公式求得树冠投影面积后进行计算。木本层植物的总盖度即各木本植物物种盖度之和。每个样方中的每一木本植物物种,选取3个植株上不同方向的成熟展开叶片至少15片,用浸湿的滤纸包裹后带回实验室。样方内植株数量不足3个的物种则按采集出现的所有植株的不同方向的叶片。对于草本植物,则在木本植物样方的四个角设置4个2 m×2 m的小样方,记录每个出现的草本物种的多度、盖度及该物种样方内出现的最大植株高度。每一样方中每种草本植物随机选取3株采集整株,量取其株高,进行平均计算后得到该物种的平均株高。草本植物取不同小样方内的4株个体的20片成熟叶片,仍用湿滤纸包裹后带回实验室。
采集得到的木本和草本植物叶片在实验室称得每片叶片的鲜重并拍照,照片利用ImageJ软件测算每片参与分析的叶片的叶面积(LA)。对于条形叶片及针叶,则分别采用叶长×叶宽及针叶圆柱体表面积的一半来表示其叶面积。每个样方中每个物种的叶面积为该物种所有采集到叶片单叶面积的平均值。随后将叶片置于烘箱,75 ℃烘干2 d,测其干重,并计算每个样方中每一物种的叶片干物质含量(LDMC)、比叶面积(SLA)。
叶片干物质含量=单枚叶片干重/单枚叶片鲜重
比叶面积=单枚叶片面积/单枚叶片干重
样方中每一物种的叶片干物质含量和比叶面积为该物种所有采集到叶片的干物质含量或比叶面积的平均值。
每个样方沿坡向方向的左上-右下角的对角线分别在对角及对角线的中心处取得土样,每个取土点取2层土壤,每层深度为10 cm,称得每份土壤样本的湿重。土壤样本带回实验室后自然风干1个月,测得土壤的干重,以此计算土壤水含量(SWC)。风干后的土壤过筛后,用Cleverchem间断化学分析仪分析土壤中的铵态氮(AN)、硝态氮(NN)、总氮(TN)及速效磷(RAP)含量,用Eppendoff pH计测定1 g/100 mL土壤水溶液的pH值。
1.3数据处理
研究共获得141种木本植物、190种草本植物的群落信息和性状信息,以及15种环境因子信息。
根据群落学信息,分别计算每个样方中所有植物物种以及草本层中林下草本植物的重要值,计算公式为:IV=(Hr+Dr+Gr)/3
式中:Hr为相对高度,即某一物种平均高度/所有物种的总平均高度;Dr为相对多度,即某一物种总株数/所有物种总株数;Gr为相对盖度,即某一物种盖度/所有物种盖度总和[28]。重要值反映的是物种在群落中或草本层中的相对重要程度。它既可以代表群落中物种的组成,也可以用来加权各物种的功能性状值以求得群落加权性状值。
同时,以各样方海拔、坡度、木本层盖度、土壤含水量(包含2层)、土壤pH(包含2层)、土壤铵态氮含量(包含2层)、土壤硝态氮含量(包含2层)、土壤全氮含量(包含2层)、土壤速效磷含量(包含2层)等环境因子作为自变量,以群落的全物种及草本层物种的重要值作为因变量进行分析。分别对全物种及草本层物种的数据进行除趋势对应分析(DCA),根据其1轴的长度(1轴长均>9)从而确定使用典范对应分析(CCA)进行生境-物种关联分析。CCA是群落生态学中常用的降维排序方法,它可以直观地反映出不同环境梯度下样方变化的趋势[29-30]。在进行分析之前,先检验了各环境因子之间的方差膨胀因子,发现土壤1、2层含水量,土壤1、2层pH以及土壤1、2层铵态氮含量的多重共线性非常严重(VIF>100),土壤1、2层全氮含量,土壤1、2层硝态氮含量,土壤1、2层速效磷含量也有较严重的多重共线性(VIF>50)。为避免多重共线对模型的干扰,选用与植物根系关系更加密切的第二层土壤指标(pH-2、AN-2、TN-2、NN-2、RAP-2)作为进行CCA的土壤因子。为避免冗余变量的共辄效应,在CCA分析中向前选择,并进行Monte Carlo检验,挑选出对物种分布显著的环境因子[20]。DCA和CCA均在软件Canoco 4.5中完成。
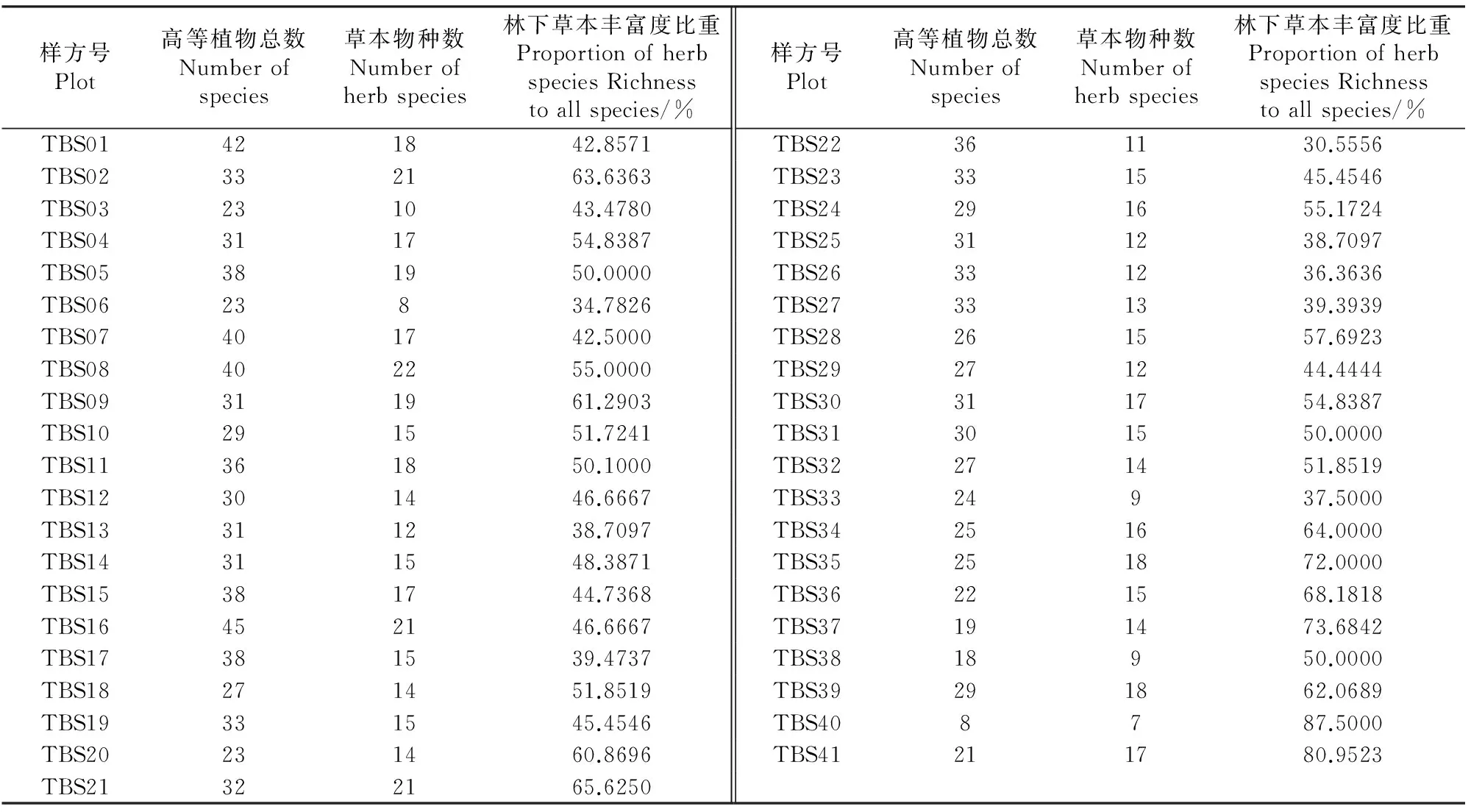
对于包含所有物种的群落物种信息以及草本层的物种信息,分别以其物种丰富度(richness)和计算得出的香农-威纳指数(Shannon-Wiener)作为该群落物种的α多样性度量。为验证生境因子与群落全物种及草本物种α多样性的关系,又对各生境因子进行了Pearson相关分析,选出最能够反映全部生境影响的海拔作为环境梯度,探讨海拔梯度上群落全物种及草本物种多样性的变化。物种多样性用R 3.1.1下的vegan包以及基于Excel 2003的Biotools组件进行计算,生境因子间的Pearson相关用SPSS 19.0进行分析,海拔与多样性间关系的拟合与作图用Origin 8.0软件进行。此外,还统计了各样方所有物种及林下草本物种的丰富度,以及林下草本物种所占全部物种丰富度的比例。
对于获得的植物的最大株高、叶面积、比叶面积、叶片干物质含量等功能性状信息,分别用群落内全物种及草本层物种的重要值进行加权,计算出群落内全物种的群落加权性状值以及林下草本物种层的加权性状值。各样方群落和草本物种层的加权性状值为样方中每一物种或草本层每一物种的性状值乘以其在群落或草本层中重要值之和。
为探究生境因子对于性状值变化的贡献程度,与研究样方物种组成的方法类似,首先仍对样方的群落性状值、草本层性状值及各生境因子进行DCA,由于DCA 1轴长度<9,因此选择冗余分析(RDA)作为量化生境因子和性状值变化关系的分析方法。RDA也是群落生态学常用的降维排序方法,只是当DCA 1轴长度较小时,其比CCA更可靠。以每个样方的15种生境因子(土壤1、2层pH,土壤1、2层含水量,土壤1、2层硝氮含量,土壤1、2层铵氮含量,土壤1、2层总氮含量,土壤1、2层速效磷含量,海拔,木本层盖度,坡度)作为自变量,以群落和草本植物层的加权性状值分别作为因变量进入模型进行RDA。RDA及作图在Canoco 4.5下进行。
为探究群落所有物种和林下草本层物种的多样性及功能性状变化模式的异同,又对各样方所有物种及草本植物间的α多样性值(以Shannon-Wiener指数代替)和不同性状的群落加权性状值进行了Pearson相关分析,该分析仍用SPSS19.0 软件完成。
2结果与分析
2.1群落和草本层物种的重要值
各样方群落优势种及草本物种重要值最高的物种见表2。
由表2可以看出,样方基本涵盖了太白山的主要森林类型(太白山自下而上的垂直带森林类型分别为锐齿栎林、辽东栎林、红桦林、糙皮桦林、巴山冷杉林、太白红杉林、杜鹃灌草丛)。在低海拔地区,草本层重要值较高的物种为荩草、铃兰、糙苏、升麻、穿龙薯蓣等物种;在中海拔地区为崖棕、淫羊藿、点叶薹草;高海拔地区则为圆齿狗哇花、以及薹草属的几个物种。各个样方的草本层优势物种分属15科,其中莎草科为草本层优势物种的第一大科,41个样方中有15个样方草本优势物种属莎草科。其他草本层优势较多的科还有小檗科、薯蓣科、毛茛科、禾本科及菊科(各有3个样方的草本优势种属之)。这种模式与调查样方中出现的各科的物种数有关,如表1。
2.2环境因子与物种组成的关系
基于物种重要值信息以及筛选出的9个环境因子,利用CCA分析二者的关系(表3、4,图1、2)。
图1、2中的数字编号为样方号,箭头的长短代表了该环境因子能解释的物种组成变化的大小,两两箭头的夹角代表了环境因子间两两的关系,锐角夹角越小,二者的相关程度越高;夹角为直角表明二者互相独立,夹角为钝角时,角度越大则表明二者的负关联程度越大。由表4知,CCA的前4轴占到群落所有物种分析下总特征根值的68.9%以及林下草本物种分析下总特征根值的66.0%,能够反映大部分的排序信息。对于样方内群落的所有物种而言,CCA的1轴可解释物种变化的8.9%,前4轴一共可解释物种变化的23.9%:其中第1轴与海拔、土壤含水量、铵态氮、总氮、速效磷等显著相关;2轴与土壤pH及硝态氮含量显著相关;3轴则与坡度、木本层盖度显著相关;4轴则与土壤总氮显著相关。对于草本层物种而言,CCA的1轴仅能解释物种变化的7.2%,前4轴可解释物种变化的21.1%:其中,1轴与海拔、土壤含水量显著相关;2轴与土壤pH、海拔、硝态氮含量显著相关;3轴与坡度、木本层盖度显著相关;4轴与土壤总氮含量显著相关。CCA分析中向前选择的结果表明,对于群落所有物种而言,环境变量进入模型的顺序依次为土壤含水量、海拔、木本层盖度、坡度、总氮含量、速效磷含量、土壤pH、硝态氮含量、铵态氮含量;对于林下草本层植物而言,环境变量进入模型的顺序依次为土壤含水量、海拔、木本层盖度、总氮含量、坡度、速效磷含量、土壤pH、硝态氮含量、铵态氮含量,变量进入模型的顺序反映了不同环境因子对于物种组成变化贡献度的大小。因此,影响群落中所有物种和林下草本层植物物种组成变化的主要环境因子都是土壤含水量、海拔和木本层盖度。此外,虽然环境因子能解释的群落及草本层物种组成变化的程度都不高(表3、4),但CCA的1轴仍能大体区分各样方的高、中、低海拔范围(图1、2,表1)。CCA的1轴与海拔和土壤含水量密切相关(表3、4),因此,海拔和土壤含水量仍能大致区分和反映高、中、低海拔下群落物种及草本层物种的组成趋势。
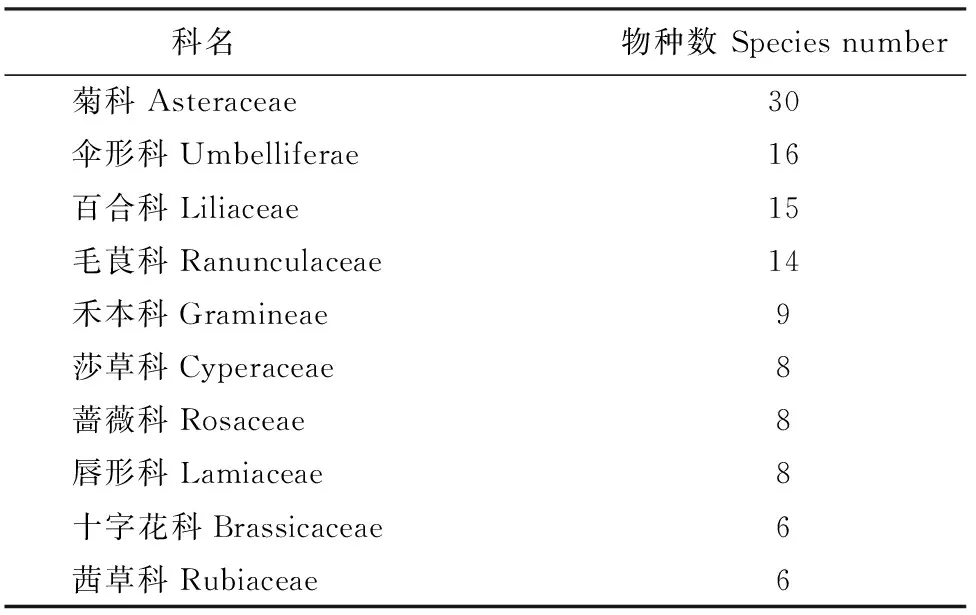
表1 样方中草本物种数最多的科及物种数
2.3环境因子与物种多样性的关系
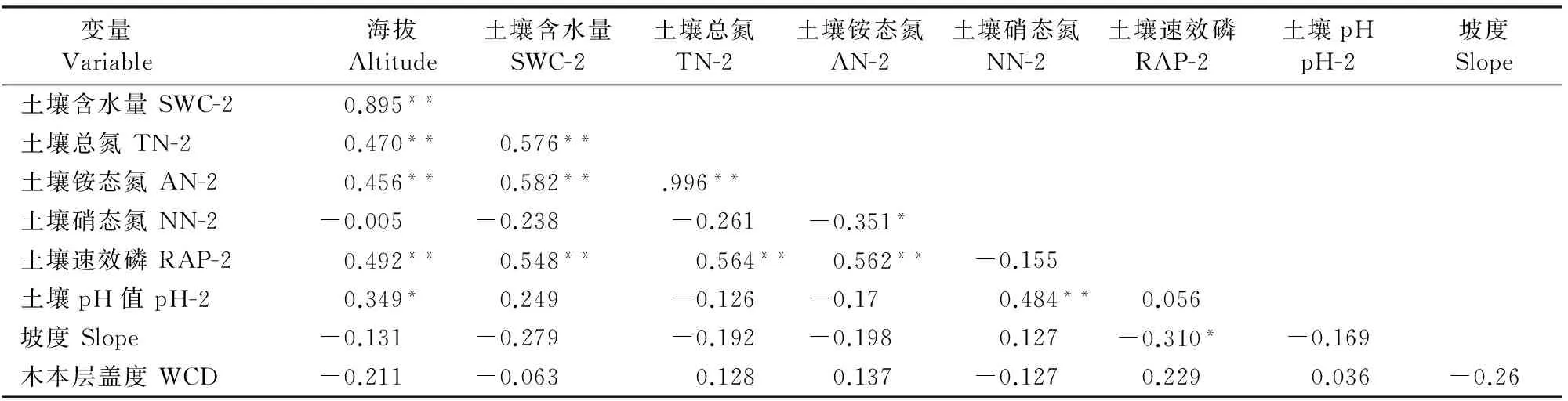
在分析环境因子对群落及林下草本物种的多样性变化的影响之前,首先对各环境因子做了Pearson相关分析。相关分析的结果见表5。
环境因子间的Pearson相关分析结果表明,海拔与土壤含水量、土壤总氮含量、土壤铵态氮含量、土壤速效磷含量呈极显著正相关关系,与土壤pH呈显著正相关关系。由于海拔差异是导致水热环境不同的主要因素,并进一步影响着土壤因子的理化性质,因此可以认为,海拔差异是导致土壤因子变化的原因,故而可以只探讨海拔与物种多样性间的关系。进一步对群落所有物种和草本层物种的α多样性(以Shannon-Wiener指数代表)与坡度、木本层盖度做Pearson分析发现,群落α多样性和草本层植物α多样性与坡度、木本层盖度均无明显相关关系(全物种多样性-坡度Pearson=-0.115,P=0.473;全物种多样性-木本层盖度Pearson=-0.001,P=0.995;草本层物种多样性-坡度Pearson=0.265,P=0.094;草本层物种多样性-木本层盖度Pearson=0.137,P=0.394),因此本研究只对海拔梯度上群落所有物种及草本层物种的多样性进行分析,结果见图3、4。
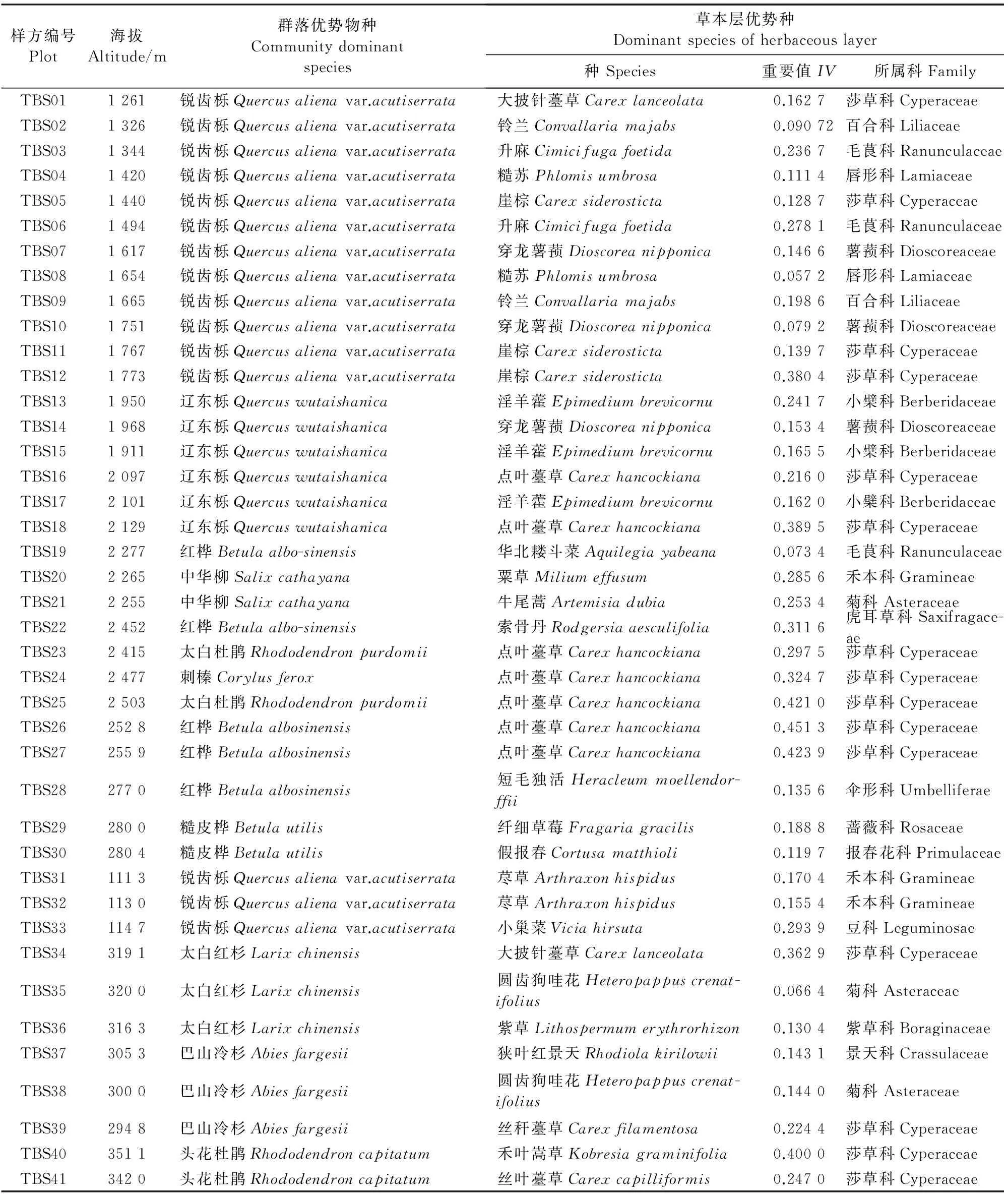
表2 样方群落优势种及草本层优势种重要值表
注:**P<0.01;*0.01 Note:In the table,the value of soil means the second layer soil which took from 10-20 cm depth soil;The same as below. 表4 草本层物种典范对应分析(CCA)排序及蒙特卡罗检验结果 横纵坐标分别为各变量在CCA轴1和轴2上的得分,1~41为样方编号。各环境变量:Alt为海拔,Slope为坡度,WCD为木本层盖度,pH-2为第2层土壤酸碱度,SWC-2为第2层土壤含水量,AN-2为第2层土壤铵态氮含量,NN-2为第2层土壤硝态氮含量,TN-2为第2层土壤总氮含量,RAP-2为第2层土壤速效磷含量;图2同。 Abscissa represents CCA 1 axis and ordinate represents CCA 2 axis,the number of the axis is the score.Number 1-41 represent the plot 1-41.Alt equals altitude,Slope equals slope of every plot,WCD equals the cover of woody plant layer,pH-2 equals the second layer soil's pH value,SWC-2 equals the second layer soil's water content,AN-2 equals the second layer soil's ammonium nitrogen content,NN-2 equals the second layer soil's nitrate nitrogen content,TN-2 equals the second layer soil's total nitrogen content and RAP-2 equals the second layer soil's rapidly available phosphorus content;The same as Fig.2. Fig.1The graph of CCA with variables and plots in community 由图3可知,包含群落所有物种的群落Shannon-Wiener指数与海拔的关系呈单峰模式,即随海拔的升高,群落的α多样性呈现先升高后降低的模式,且最高点出现在海拔2 200 m左右。林下草本层植物的α多样性与海拔的关系模式与群落近似但又不甚相同。通过拟合后发现,林下草本植物α多样性与海拔的关系较弱,随海拔的升高,有小幅下降的趋势(图4)。对群落α多样性与林下草本植物α多样性的Pearson相关分析表明,二者的的变化模式的关联程度较显著(Pearson=0.458,P=0.003)。 物种丰富度也是生物多样性的一种度量。各样方的群落物种丰富度、草本层物种丰富度及草本层物种丰富度占比的统计结果如表6所示。 图2 不同环境因子与林下草本物种组成的CCA排序图 变量 Variable 海拔Altitude土壤含水量SWC-2土壤总氮TN-2土壤铵态氮AN-2土壤硝态氮NN-2土壤速效磷RAP-2土壤pHpH-2坡度Slope土壤含水量SWC-20.895**土壤总氮TN-20.470**0.576**土壤铵态氮AN-20.456**0.582**.996**土壤硝态氮NN-2-0.005-0.238-0.261-0.351*土壤速效磷RAP-20.492**0.548**0.564**0.562**-0.155土壤pH值pH-20.349*0.249-0.126-0.170.484**0.056坡度Slope-0.131-0.279-0.192-0.1980.127-0.310*-0.169木本层盖度WCD-0.211-0.0630.1280.137-0.1270.2290.036-0.26 可以发现,在各个样方中,林下草本植物物种数能占到群落物种丰富度的37.5%~87.5%,比例较高,尤其在高海拔的灌草丛群落样方中(TBS40、TBS41)所占的比例更大。 2.4环境因子与功能性状的关系 对所有15个环境因子与群落及草本层的4个加权的功能性状值(叶面积、比叶面积、叶干物质含量、最大株高)进行冗余分析(RDA,因变量的DCA轴1长度小于2),所得结果见图5、6。 图5、6中数字与箭头代表的含义与图1、2相同。对于群落所有物种和林下草本物种,RDA前4轴均能解释环境-性状关系的100%,故而能反映全部的排序信息。在群落所有物种中,RDA1轴能解释性状变化的71.2%(P=0.001),而前4轴共可解释79.5%(P=0.001),因此,RDA1轴即可大致反映生境与性状的变化规律:RDA的1轴与海拔呈极显著正相关(相关系数0.9526,P<0.001)也与土壤含水量、总氮含量呈显著正相关。在林下草本植物中,RDA1轴对于性状变化的解释程度为54.7% (P=0.01),前4轴共可解释形状变化的58.8% (P=0.012),故而RDA1轴仍是反映林下草本性状变化规律的最好度量。林下草本植物的RDA1轴仍与海拔关系最紧密(相关系数0.873 5,P<0.01)也与土壤含水量、总氮含量、铵态氮含量、速效磷含量显著相关。 图3 海拔与群落α多样性的关系 图4 海拔与草本层α多样性的关系 样方号Plot高等植物总数Numberofspecies草本物种数Numberofherbspecies林下草本丰富度比重ProportionofherbspeciesRichnesstoallspecies/%样方号Plot高等植物总数Numberofspecies草本物种数Numberofherbspecies林下草本丰富度比重ProportionofherbspeciesRichnesstoallspecies/%TBS01421842.8571TBS22361130.5556TBS02332163.6363TBS23331545.4546TBS03231043.4780TBS24291655.1724TBS04311754.8387TBS25311238.7097TBS05381950.0000TBS26331236.3636TBS0623834.7826TBS27331339.3939TBS07401742.5000TBS28261557.6923TBS08402255.0000TBS29271244.4444TBS09311961.2903TBS30311754.8387TBS10291551.7241TBS31301550.0000TBS11361850.1000TBS32271451.8519TBS12301446.6667TBS3324937.5000TBS13311238.7097TBS34251664.0000TBS14311548.3871TBS35251872.0000TBS15381744.7368TBS36221568.1818TBS16452146.6667TBS37191473.6842TBS17381539.4737TBS3818950.0000TBS18271451.8519TBS39291862.0689TBS19331545.4546TBS408787.5000TBS20231460.8696TBS41211780.9523TBS21322165.6250 横纵坐标分别为各变量在RDA1轴和2轴上的得分,1-41为样方编号。各环境变量:Alt为海拔,Slope为坡度,WCD为木本层盖度,pH-1/pH-2为第1、2层土壤酸碱度,SWC-1/SWC-2为第1、2层土壤含水量,AN-1/AN-2为第1、2层土壤铵态氮含量,NN-1/NN-2为第1、2层土壤硝态氮含量,TN-1/TN-2为第1、2层土壤总氮含量,RAP-1/RAP-2为第1、2层土壤速效磷含量;图6同。 Abscissa represents RDA 1 axis and ordinate represents RDA 2 axis,the number of the axis is the score.Number 1-41 represent the plot 1-41.Alt equals altitude,Slope equals slope of every plot,WCD equals the cover of woody plant layer,pH-1/pH-2 equal two layers soil's pH value,SWC-1/SWC-2 equal two layers soil's water content,AN-1/AN-2 equal two layers soil's ammonium nitrogen content,NN-1/NN-2 equal two layers soil's nitrate nitrogen content,TN-1/TN-2 equal two layer soil's total nitrogen content and RAP-1/RAP-2 equal two layers soil's rapidly available phosphorus content;The same as Fig.6. Fig.5The graph of RDA with variables and community trait value 图6 不同环境因子与草本层性状值的RDA排序图 为探究群落所有物种与草本物种功能性状的变化模式的异同,又对群落和草本层的4个性状值分别对应做了Pearson相关分析。结果显示,群落所有物种的群落加权性状值与草本层物种的加权性状值存在一定的关联。其中,二者的比叶面积(SLA)的关联程度最高(Pearson=0.828,P<0.01),其次为叶片干物质含量(LDMC,Pearson=0.733,P<0.01)、叶面积(LA,Pearson=0.705,P<0.01),但群落与草本层最大株高(Hmax)的加权性状值之间没有显著关联(Pearson=0.262,P>0.1)。 3讨论 对于草本层物种的重要值信息分析表明,在某些样方(特别是高海拔地区的样方)中,某些草本植物的重要值较高,如果忽略草本植物而只研究其中木本植物的多样性、结构和性状,显然是不妥当的,森林群落中的木本植物研究虽然能从一个方面反映群落的生态功能和多样性维持机制[31],但它毕竟不能够完整地反映整个群落的特征。因此,结合林下草本植物研究群落是非常必要的。 典范对应分析(CCA)对于环境因子与物种组成的关系表明,不论对于整个群落还是对于林下草本层植物,环境因子仅能解释很少的物种组成变化,这也与前人研究的结果近似[20]。例如,秦晓威[27]发现,不同的森林群落中,仅27.0%~36.3%的草本物种分布格局与生境显著相关。 不过,对于群落和林下草本,与海拔和土壤含水量相关的CCA分析的1轴却可以大致区分出样方的海拔范围(高、中、低,图1、2,表1),说明环境因素仍然影响着较大海拔范围的群落和草本层物种组成。前人已经发现林下草本植物与土壤含水量关系密切[32]。而对于海拔因素,在较小的海拔范围内,物种的组成模式可能是无序的。这可能是由于除参与分析的环境因子之外其他微环境的不同导致的,也可能是由于稀有种的干扰导致的。在进行CCA时,稀有物种有时可占到样方内全部物种的60%,这会导致对物种组成数据排序的失真。前人的研究发现,在山区森林中,海拔是影响物种分布格局的主要因子[12,33-35],较小海拔范围内物种组成的无序性造成了CCA中环境对于物种组成变化的较低解释程度。在较大的海拔范围内,物种组成的变化可以由CCA进行大致的区分,这表明在较大的尺度上,植物物种的组成在相似的生境中具有相似的组成模式,也从侧面证明林下草本植物在较大的空间尺度上也表现出一定的地带性。 针对生境梯度与物种多样性变化的关系,前人的研究结果已经表明其多样性与气候和土壤养分条件相关[36-37]。本研究发现,在海拔梯度上,群落所有物种和草本层物种的多样性有着弱的关联关系,其总趋势都是沿海拔多样性降低,这与岳明等在本区研究的结果相一致[38]。但是,与林下草本植物相比,群落所有物种的多样性与海拔的关系更显著,拟合优度更好。群落物种的Shannon-Wiener指数沿海拔呈现先升后降的单峰模式,这也与前人在本区研究的结果类似[39]。通常认为,在中等海拔处,水热条件最好,分化的生态位多,因此有最高的物种多样性[27]。而低海拔地区受人为干扰较严重,多样性水平较中海拔为低。林下草本植物的多样性变化没有随海拔表现出非常显著的关系,但有随海拔下降的趋势。这与Wang等[32]的研究结果吻合,但与马晓勇在太岳山的研究结果不一致。太岳山地区植被物种多样性虽然受光照、温度、湿度、人为干扰等因子的影响会出现波动性,但总体趋势为随着海拔高度的升高而增加[40],这可能是因为太岳山的海拔梯度明显小于太白山,在某一海拔范围内,草本物种多样性可能沿海拔升高而增加,但在更大的海拔范围下,其多样性变化模式则会发生变化。而秦晓威对太白山林下草本植物研究发现,草本植物丰富度的海拔梯度格局呈单峰分布,且丰富度峰值在海拔 1200~1300 m[27]。不过,秦晓威在本区的研究梯度也较小,例如,其海拔梯度差只有约900 m。 林下草本植物与群落不甚相同的多样性模式表明,海拔可能并不是决定林下草本植物多样性的最主要的环境因子。与李双喜和马晓勇等的研究不同,太白山地区的林下草本物种多样性也与林分郁闭度即木本层盖度没有明显的关联[40-41]。虽然前人的研究认为林隙面积会对林下草本物种的多样性产生影响,但林隙面积并没有反映在木本层植物的总盖度上[42]。而本区域草本物种多样性与坡度无关的模式也与有的研究者的发现不一致[43-44],这可能是由于本研究所选取的样方坡度变化不大,坡度因素导致的林下草本多样性变化相对其他因子为小。此外,在本研究区域内,林下草本植物物种数能够占群落物种丰富度水平的37.5%~87.5%,尤其在高海拔地区所占比例很高,其多样性水平是很高的。这表明林下草本植物对维持太白山地区物种多样性起到了重要的作用[38]。 植物的功能性状是反映反映生境过滤对群落植物物种组成的重要表征,也是当前研究群落构建模式的重要手段。对于群落所有物种4个群落性状值与环境因子的RDA表明,环境对于群落物种的性状具有明显的筛选和修饰作用。海拔梯度对草本物种的影响,往往是通过水分、热量、光照等因子起作用[45]。在太白山地区,由于多地形雨,海拔能够影响降水量,进而影响土壤含水量。因此,海拔是影响太白山地区物种功能性状变化的主要因子。对于林下草本植物来说,环境因子能够解释的草本层性状值变化要小于群落内全物种,但其RDA轴1仍能解释55%左右的性状变化。这表明,林下草本物种的功能性状依然受到环境因子的过滤。林下草本物种的RDA1轴除与海拔和土壤含水量相关外,还与土壤总氮含量、铵态氮含量及速效磷含量有较高的相关性,说明草本物种的性状变化受到更多环境因子的影响。 此外,群落所有物种和林下草本物种4个功能性状的相关性分析表明,草本植物在SLA、LDMC、LA等叶经济性状上与群落的性状值共变,也说明在叶性状方面,林下草本植物与群落其他物种对相似的生境做出了相似的反应,林下草本植物与木本植物的性状一样同样可以反映群落物种对于生境的响应。不过,在最大株高这一性状上,林下草本植物与群落所有植物对于环境的反应不一致,没有明显的关联关系。在海拔梯度上,群落内重要值较高的木本高度往往随海拔的升高而有降低的趋势,但草本植物这一趋势不明显。例如,在中低海拔的某些林分郁闭度较高的区域,由于下层草本大都是耐阴植物,不需要进行光竞争,其草本层物种的株高往往较小;而在高海拔地区尤其是灌草丛区域,由于木本植物植株较为矮小,反而会有一些较高的阳生草本物种出现。鉴于林下草本物种在某些性状上与木本植物相比沿梯度具有不同的变化模式,因此,包含林下草本植物的群落功能性状研究是科学的、必要的。 总之,在本研究区域,林下草本物种的物种组成与能够通过土壤含水量与海拔有关的CCA轴1区分,而物种的多样性变化则随海拔的升高而有下降的趋势。对于草本物种的功能性状,也与海拔及土壤理化因子有关。与此同时,在上述这些方面,林下草本物种与群落所有物种的一些变化模式又有所不同。由此可见,林下草本物种研究对于了解不同梯度下森林群落的物种组成、多样性变化、功能性状变化、预测群落冠型[45]等方面都有着十分重要的作用,对于森林群落构建机制、森林物种多样性的维持、森林物质循环和能量流动以及森林的保护和管理等研究提供了又一可靠方向。 参考文献: [1]BHAT D M,MURALI K S.Phenology of understory species of tropical moist forest of Western Ghatsregion of Uttara Kannada district in South India[J].CurrentScience,2001,81:799-805.[2]AUGUSTO L,DUPOUEY J L,RANGER J.Effects of tree species on understory vegetation and environmental conditions in temperate forests[J].AnnualsofForestScience,2003,60(8):823-831. [3]WHIGHAM D F.Ecology of woodland herbs in temperate deciduous forests[J].AnnualReviewofEcology,Evolution,andSystematics,2004,35:583-621. [4]GILLIAM F S.The ecological significance of the herbaceous layer in temperate forest ecosystems[J].BioScience,2007,57(10):845-858. [5]SIEBERT S.From shade to sun-grown perennial crops in Sulawesi,Indonesia:implications for biodiversity conservation and soil fertility[J].BiodiversityandConservation,2002,11:1 889-1 902. [6]RAMADHANIL R,TJITROSOEDIRDJO S S,SETIADI D.Structure and composition of understory plant assemblages of six land use types in the Lore Lindau National Park,Central Sulawesi,Indonesia[J].BangladeshJournalofPlantTaxon,2008,15:1-12. [7]AKPO L.Phenological interactions between tree and understory herbaceous vegetation of a sahelian semi-arid savanna[J].PlantEcology,1997,131:241-248. [8]Michael AJ,George RP.The response of herbaceous-layer vegetation to anthropogenic disturbance in intermittent stream bottomland forests of southern Indiana,USA[J].PlantEcology,2000,151:223-237. [9]LIIRA J,ZOBEL K,MAGI R,etal.Vertical structure of herbaceous canopies:the importance of plant growth-form and species-specific traits[J].PlantEcology,2002,163:123-134. [10]杨昆,管东生.林下植被的生物量分布特征及其作用[J].生态学杂志,2006,25(10):1 252-1 256. YANG K,GUAN D S.Biomass distribution and its funtioning of forest understory vegetation[J].ChineseJournalofEcology,2006,25(10):1 252-1 256. [11]郝占庆,郭水良.长白山北坡草本植物分布与环境关系的典范对应分析[J].生态学报,2003,23(10):2 000-2 008. HAO Z Q,GUO S L.Canonical correspondence analysis on relationship of herbs with their environments on northern slope of Changbai Mountain[J].ActaEcologicaSinica,2003,23(10):2 000-2 008. [12]范玉龙,胡楠,丁圣彦,等.伏牛山自然保护区森林生态系统草本植物功能群的分类[J].生态学报,2008,28(7):3 092-3 101.FAN Y L,HU N,DING S Y,etal.The classification of plant functional types based on dominant herbaceous species in forest ecosystem at Funiu Mountain National Natural Reserve[J].ActaEcologicaSinica,2008,28(7):3 092-3 101. [13]夏富才,潘春芳,赵秀海,等.长白山原始阔叶红松林林下草本植物多样性格局及其影响因素[J].西北植物学报,2012,32(2):370-376. XIA F C,PAN C F,ZHAO X H,etal.Effect of overstory on the seasonal variability of understory herbs in primary broad-leaved Korean pine forest of Changbai Mountain[J].ActaBot.Boreal.-Occident.Sin,2012,32(2):370-376. [14]杨再鸿.海南岛桉树人工林林下植物多样性的比较研究[D].海南儋州:华南热带农业大学,2001. [15]Muller RN.Nutrient relations of the herbaceous layer in deciduous forest ecosystems.//Gilliam FS,Roberts MR.(Eds.).The Herbaceous Layer in Forests of Eastern North America[M].Oxford University Press,New York,2003:15-37.[16]奇凯,张春雨,侯继华,等.赤峰市沙地油松林草本植物多样性及种间关联动态[J].生态学报,2010,30(18):5 106-5 112. QI K,ZHANG C Y,HOU J H,etal.Dynamics of species diversity and interspecific associations of herbaceous plants in aPinusTabulaeformisforest on a sandy site in Chifeng,China[J].ActaEcologicaSinica,2010,30(18):5 106-5 112. [17]姚茂和,盛炜彤,熊有强.杉木林林下植被及其生物量的研究[J].林业科学,1991,27(6):644-648. YAO M H,SHENG W T,XIONG Y Q.Studies on understory and its biomass in Chinese Fir stands[J].ScientiaSilvaeSinicae,1991,27(6):644-648. [18]茹文明,张金屯,毕润成,等.山西霍山森林群落林下物种多样性研究[J].生态学杂志,2005,24(10):1 139-1 142. RU W M,ZHANG J T,BI R C,etal.Species diversity of undergrowths in Huoshan Mountains of Shanxi Province[J].ChineseJournalofEcology,2005,24(10):1 139-1 142. [19]田锴.古田山亚热带常绿阔叶林草本植物群落结构和多样性格局研究[D].浙江金华:浙江师范大学,2011. [20]余敏.山西灵空山林分冠层结构与草本植物群落分析[D].北京:北京林业大学,2013. [21]YE J,HAO Z Q,XIE P,etal.Habitat associations of saplings and adults in an old-growth temperate forest in the Changbai mountains in northeastern China[J].ForestryStudiesinChina,2011,13(1):13-22. [22]冯云,马克明,张育新,等.辽东栎林不同层植物沿海拔梯度分布的DCCA分析[J].植物生态学报,2008,32(3):568-573. FENG Y,MA K M,ZHANG Y X,etal.DCCA analysis of plant species distributions in different strata of oak(Quercusliaotungensis) forest along an altitudinal gradient in Dongling Mountain,China[J].JournalofPlantEcology(Chinese Version),2008,32(3):568-573. [23]傅志军,郭俊理.秦岭太白山植被的群落特征[J].宝鸡师范学院学报(自然科学版),1992,(1):70-75. FU ZH J,GUO J L.The characters of community on the vegetation of the Taibai Mountain in the Qinling[J].JournalofBaojiTeacherCollege(Natural Science),1992,(1):70-75. [24]李景侠,张文辉,杨赵洁.陕西省地方重点保护植物及保护对策[J].西北林学院学报,1999,14(3):6-12. LI J X,ZHANG W H,YANG Z J.On the important native conservated plants in Shaanxi Province and strategies of conservation[J].JournalofNorthwestForestryUniversity.1999,14(3):6-12. [25]高贤明,黄建辉,万师强,等.秦岭太白山弃耕地植物群落演替的生态学研究[J].生态学报,1997,17(6):619-625. GAO X M,HUANG J H,WAN S Q,etal.Ecological studies on the plant community succession on the abandoned farm land in Taibaishan,Qinling Mountains[J].ActaEcologicaSinica,1997,17(6):619-625. [26]朱志诚.秦岭太白山森林主要类型特征及其分布规律[J].陕西林业科技,1981,(5):29-39. ZHU Z C.The regulation and characteristic of dominant type of forest in Taibai Mountain,Qinling Mountains[J].ShaanxiForestScienceandTechnology,1981,(5):29-39. [27]秦晓威.秦岭太白山草本植物多样性格局及其对林隙干扰的响应[D].陕西杨凌:西北农林科技大学,2012. [28]CURTIS J T,MCINTOSH R P.An upland forest continuum in the prairie forest border region of Wisconsin[J].Ecology,1951,32:476-496. [29]邱扬,张金屯.DCCA排序轴分类及其在关帝山八水沟植物群落生态梯度分析中的应用[J].生态学报,2000,20(2):199-206.QIU Y,ZHANG J T.The ordination axes clustering based on detrended canonical correspondence analysis ordination and its application to the analysis of the ecological gradients of plant communities[J].ActaEcologicaSinica,2000,20(2):199-206.[30]许彬,张金屯,杨洪晓,等.京西百花山植物群落数量分析[J].北京师范大学学报(自然科学版),2006,42(1):90-94. XU B,ZHANG J T,YANG H X,etal.Quantitative analysis of plant communities in Baihua Mountains,Western Beijing[J].JournalofBeijingNormalUniversity(Natural Science Edition),2006,42(1):90-94. [31]MCGILL B J,ETIENNE R S,GRAY J S.Species abundance distributions:moving beyond single prediction theories to integration within an ecological framework[J].EcologyLetters,2007,10:995-1 015. [32]WANG H F,LENCINAS M V,FRIEDMAN C R,etal.Understory plant diversity assessment of Eucalyptus plantations over three vegetation types in Yunnan,China[J].NewForests,2011,42:101-116. [33]娄安如.天山中段山地植被的生态梯度分析及环境解释[J].植物生态学报,1998,22(4):364-372. LOU A R.Ecological gradient analysis and environmental interpretation of mountain vegetation in the middle stretch of Tianshan Mountain[J].ActaPhytoecologicaSinica,1998,22(4):364-372. [34]沈泽昊,张新时.三峡大老岭地区森林植被的空间格局分析及其地形解释[J].植物学报,2000,42(10):1 089-1 095. SHEN Z H,ZHANG X S.The spatial pattern and topographic interpretation of the forest vegetation at Dalaoling region in the Three Gorges[J].ActaBotanicaSinica, 2000,42(10):1 089-1 095. [35]刘秋锋,康慕谊,刘全儒.中条山混沟地区森林乔木种的数量分类与环境解释[J].植物生态学报,2006,30(3):383-391. LIU Q F,KANG M Y,LIU Q R.Quantitative classification and environmental interpretation of forest tree species in Hungou,Zhongtiao Mountain[J].JournalofPlantEcology(Chinese Version),2006,30(3):383-391. [36]FLAVIA R C.Mesoscale gradients of herb richness and abundance in central Amazonia,Biotropica,2006,38(6):711-717. [37]NORTH M,OAKLEY B,FIEGENER R.Influence of light and soil moisture on Sierran mixed-conifer understory communities[J].PlantEcology,2005,177:13-24. [38]岳明,任毅,党高弟,等.佛坪国家级自然保护区植物群落物种多样性特征[J].生物多样性,1999,7(4):263-269. YUE M,REN Y,DANG G D,etal.Species diviersity of higher plant communities in Foping National Reserve[J].ChineseBiodiversity,1999,7(4):263-269. [39]岳明,张林静,党高弟,等.佛坪自然保护区植物群落物种多样性与海拔梯度的关系[J].地理科学,2002,22(3):349-354. YUE M,ZHANG L J,etal.The relationships of higher plants diversity and elevation gradient in Foping National Reserve[J].ScientiaGeographicaSinica,2002,22(3):349-354.[40]马晓勇,上官铁梁.太岳山森林群落物种多样性[J].山地学报,2004,22(5):606-612. MA X Y,SHANGGUAN T L.Species diversity of the forest communities in Taiyue Mountain,Shanxi[J].JournalofMountainScience,2004,22(5):606-612. [41]李双喜,朱建军,张银龙,等.人工马褂木林下草本植物物种多样性与林分郁闭度的关系[J].生态与农村环境学报,2009,25(2):20-24. LI S X,ZHU J J,ZHANG Y L,etal.Diversity of understory herbaceous species and canopy density ofLiriodendronchinenseStand[J].JournalofEcologyandRuralEnvironment.2009,25(2):20-24. [42]HADLEY A S,ROBINSON W D,FREY S J K,etal.Tropical forest fragmentation limits pollination of a keystone understory herb[J].Ecology,2014,95(8):2 202-2 212. [43]SMALL C J,MCCARTHY B C.Spatial and temporal variation in the response of understory vegetation to disturbance in a central Appalachian oak forest[J].JournaloftheTorreyBotanicalSociety,2002,129(2):136-153. [44]ROBERT J W.Mechanisms driving understory evergreen herb distributions across slope aspects:as derived from landscape position[J].PlantEcology,2008,198(2):297-308. [45]HODGSON J Y S,BUNKER G,HODGSON J R,etal.Predictive relationships between the canopy overstory and the herbaceous understory in a northeastern Wisconsin forest[M].MichiganBotanist,2005,139-147. (编辑:潘新社) Environmental Interpretation of Variations on Forest Community and Understory Herbaceous in Taibai Mountain,Shaanxi,China CHEN Yu,XU Jinshi,ZHANG Lixia,GUO Yaoxin,CHAI Yongfu,WANG Mao, ZHANG Chenguang,YUE Ming* (College of Life Sciences,Northwest University,Xi’an 710069,China) Abstract:Our study aimed to determine the relationship between the environment factors and these variation patterns.Also,we examined the difference of variation patterns between forest community and understory herb layer only.The canonical correspondence analysis (CCA) was used for testing the relationship between 9 environment factors and species composition data.A correlation test was determined the relationship between environment factors and species α diversity.Redundancy analysis (RDA) was used to analyze the importance of environment factors on plant functional traits.We also tested the Pearson correlation coefficient of community α diversity and functional traits between forest community and understory herb layer.We found that:1) 9 environment factors had a poor power to explain the change of both forest community and understory herb layer.However,the axis 1 of CCA could distinguish the altitude range of each plot in both modes.2) Along the elevation,the Shannon-Wiener index of forest community at first increased and then decreased while the Shannon-Wiener index of understory herb layer decreased monotonously.3)The Shannon-Wiener index of both community and herb layer was independence to slope and woody species layer cover degree(WCD).The axis 1 of RDA which related to altitude could explain 71.2% of total trait variation for forest community and 54.7% for understory herb layer.The variation pattern of plant max height value (Hmax) of forest community and herb layer was different. Key words:understory herbaceous species;species composition;diversity;functional traits;canonical correspondence analysis (CCA);redundancy analysis (RDA) 文章编号:1000-4025(2016)04-0784-12 doi:10.7606/j.issn.1000-4025.2016.04.0784 收稿日期:2016-01-05;修改稿收到日期:2016-03-10 基金项目:国家自然科学基金(41571500);陕西高校省级重点实验室科研项目(13JS095);陕西省中医药管理局专项(13-ZY052) 作者简介:陈煜(1990-),男,在读硕士研究生,主要从事群落生态学研究。 *通信作者:岳明,博士,教授。主要从事群落生态学和全球变化生态学研究。 E-mail:yueming@nwu.edu.cn 中图分类号:Q948.11 文献标志码:A






猜你喜欢
发明与创新·中学生(2024年6期)2024-05-29 00:00:00
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
林业勘查设计(2022年1期)2022-02-15 05:35:12
散文诗(2021年22期)2022-01-12 06:14:04
安顺学院学报(2021年4期)2021-09-16 08:11:08
少儿美术(2020年8期)2020-11-06 05:21:38
林业调查规划(2020年3期)2020-06-03 07:02:36
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
