
阿扎霉素F产生菌链霉菌211726基因转移系统的建立
2016-06-13 10:44马艳玲刘富来张敏孙宇辉洪葵
生物技术通报 2016年4期
马艳玲刘富来张敏孙宇辉洪葵
(1. 佛山科学技术学院食品与园艺学院,佛山 528231;2. 武汉大学药学院,武汉 430071)
阿扎霉素F产生菌链霉菌211726基因转移系统的建立
马艳玲1刘富来1张敏1孙宇辉2洪葵2
(1. 佛山科学技术学院食品与园艺学院,佛山 528231;2. 武汉大学药学院,武汉 430071)
旨在建立阿扎霉素F产生菌链霉菌211726的基因转移系统,以便基因敲除和外源基因表达等遗传操作。以整合型质粒pSET152和pIB139为出发质粒,通过接合转移构建了阿扎霉素F产生菌链霉菌211726的基因转移系统。结果显示25 μg/mL阿泊拉霉素可有效筛选接合子。经PCR验证,质粒成功整合到菌株链霉菌211726基因组中,接合子经多次传代后,导入的质粒 pSET152和pIB139仍稳定整合于接合子基因组上。
阿扎霉素F;链霉菌211726;接合转移;遗传稳定性
链霉菌(Streptomyces)在分类学上属于原核生物界、放线菌目、链霉菌科,是一类具有分枝丝状体的好氧革兰氏阳性细菌[1],是土壤中主要的微生物类群之一。链霉菌基因组平均大小为8 000 kb,约为大肠杆菌基因组的两倍[2]。同其它生物相比,其基因组最大特征之一就是其DNA具有极高的G+C mol%,可高达69%-78%,是迄今为止已知的G+C mol%含量最高的生物类群之一[3]。链霉菌是工业微生物中最具有商用价值的类群之一。人们已经了解到自然界中有近70%的抗生素是由链霉菌及其近缘放线菌产生的。自1928年Alexander Fleming发现第一个抗生素——青霉素以来,数以万计的抗生素正源源不断地被人们发现和创造出来并造福于人类[4-7]。
阿扎霉素F最早是由Arai[8]于1959年发现并从吸水链霉菌Streptomyces hygroscopicus var.中分离得到的。1995年,Krystofova[9]研究组对该抗生素的钙离子通道作用进行了初步的研究。
2010年,研究人员在海南省的沿海红树林根系土壤中分离纯化得到Streptomyces sp. 211726。经过发酵培养和产物结构鉴定,发现它可以同时产生13种结构略有差异的系列阿扎霉素,区别仅仅在于阿扎霉素C-23位和C-25上连接的基团不同[10-13],并对所发现的这13种化合物命名为阿扎霉素S4-S16,其中S13、S15和S16分别与国际上已报道的阿扎霉素F3a、F4a和F5a结构一致[14]。生物学活性检测表明,在这一系列化合物中以阿扎霉素F3a为代表的多种化合物都具有明显的抗尖孢镰刀菌(F.oxysporum)活性[10-13],表明该类抗生素在抗香蕉枯萎病病原菌具有良好防治作用。这些化合物还对白色念珠菌表现出明显的拮抗作用,其抗白色念珠菌(Candida albicans)的有效性与氟康唑(Fluconazole)和两性霉素B相当,就其毒性而言,阿扎霉素F系列化合物的毒性较氟康唑大,但是小于两性霉素B[15]。此外,毒性实验还表明在Streptomyces sp. 211726所产生的阿扎霉素F系列化合物中除阿扎霉素F5a外其余12种化合物均对人类结肠癌细胞HCT-116表现出一定的细胞毒作用,预示着该系列化合物在人类肿瘤疾病治疗方面的潜在应用[13]。由于致病真菌耐药性的日益严重和结构修饰研究的日益深入,因此开展以提高有效性、降低毒性为目的的结构改造研究越来越得到人们的广泛关注。但是目前关于阿扎霉素F的分子水平上的结构改造研究还尚未报道,这可能与其限制性修饰作用强,遗传操作困难有关。因此,有必要建立起阿扎霉素F产生菌链霉菌21172的接合转移体系,以推动其分子生物学研究。
本研究拟建立阿扎霉素F产生菌链霉菌211726的基因转移体系,探讨通过接合转移向链霉菌211726导入外源DNA片段的可能性,旨在为该菌的生物合成基因改造奠定基础。
1 材料与方法
1.1 材料
1.1.1 实验材料 阿扎霉素F产生菌链霉菌211726是洪葵教授赠与,基因克隆受体E.coli DH10B、接合转移供体菌E.coli ET12567(pUZ8002)、整合型质粒pSET152和pIB139(均携带阿泊拉霉素抗性基因aac(3)IV,接合转移位点oriT)均为笔者实验室保存。
1.1.2 试剂 硫链丝菌素和阿泊拉霉素购自美国Sigma公司;氯霉素、氨苄青霉素、卡那霉素、硫链丝菌素、Lambda/Hind III、限制性内切酶、DNA 回收试剂盒、质粒提取试剂盒等购自大连宝生物工程有限公司;其他常用试剂均购于上海国药集团。
1.1.3 培养基和抗生素 链霉菌211726的平板培养基为 SFMS,培养细菌的为LB培养基、孢子预萌发培养基为2×YT、接合转移使用的是SFMS培养基。
LB中抗生素终浓度为:阿泊拉霉素25 μg/mL,氯霉素25 μg/mL,卡那霉素25 μg/mL,氨苄青霉素100 μg/mL;接合转移培养基中抗生素终浓度为阿泊拉霉素25 μg/mL,萘啶酮酸25 μg/mL。
1.2 方法
1.2.1 抗生素敏感性实验 将已灭菌的SFMS培养基融化,温度降至45℃,加入适当质量浓度的抗生素(卡那霉素 0-100 μg/mL;氯霉素 0-100 μg/mL;阿泊拉霉素 0-100 μg/mL;硫链丝菌素 0-100 μg/ mL),将链霉菌211726菌株的孢子悬液涂布平板,28℃培养5 d,根据链霉菌211726菌株生长状态评定链霉菌211726菌株对抗生素的敏感性。
1.2.2 质粒DNA的跨属接合转移 将整合型质粒pSET152和pIB139通过CaCl2法导入接合转移供体菌E.coli ET12567(pUZ8002)中,借助于pUZ8002上tra基因,上述两种质粒以接合转移方式从大肠杆菌导入到链霉菌211726细胞中,并进行整合。接合转移方法主要依据文献[16]进行改进。
1.2.3 接合转移子的验证 以整合型质粒pSET152和pIB139上的阿泊拉霉素抗性基因aac(3)IV为模板,设计引物CP1和CP2以验证阳性接合转移子。CP1:5'-TTTATCACCACCGACTATTTGC-3';CP2:5'-TCATCTCGTTCTCCGCTCA-3'。按照文献[9]方法提取链霉菌211726基因组DNA并进行PCR验证。PCR条件反应程序为:98℃ 1 min;98℃ 30 s,58℃30 s,72℃ 30 s,30个循环;72℃ 延伸10 min。
1.2.4 质粒pSET152和pIB139在链霉菌211726接合转移子中的稳定性 将接合子接种于不含抗生素的SFMS平板上,待其孢子成熟(约15 d)后,再次接种于不含抗生素的SFMS平板上,连续转接3次后进行分离纯化,随机挑选单菌落,转接到含有25 μg/mL阿泊拉霉素的SFMS平板上,5 d后根据单菌落能否生长来确定质粒pSET152和pIB139在接合子中的遗传稳定性。
2 结果
2.1 链霉菌211726对不同抗生素抗性水平的检测
为确定抗生素抗性基因能否用于链霉菌211726菌株进行遗传操作的选择标记,检测了链霉菌211 726菌株对几种抗生素(阿泊拉霉素、氯霉素、卡那霉素、硫链丝菌素等)的抗性水平。结果(表1)显示,链霉菌211726菌株在含有12.5 μg /mL卡那霉素和阿泊拉霉素的培养基上不能生长,表明链霉菌211726菌株对卡那霉素和阿泊拉霉素非常敏感;链霉菌211726菌株在含有25 μg /mL氯霉素的培养基上不能生长,表明链霉菌211726菌株对氯霉素比较敏感;链霉菌211726菌株在含有硫链丝菌素100 μg/mL的培养基上都能生长,说明链霉菌211726对硫链丝菌素不敏感。鉴于本研究中使用的接合转移质粒携带有阿泊拉霉素抗性基因aac(3)IV,因此以25 μg /mL阿泊拉霉素筛选接合转移子。

表1 链霉菌211726在4种不同浓度抗生素平板上的生长情况
2.2 链霉菌211726与大肠杆菌之间两亲接合转移
通过两亲接合转移法分别将质粒pSET152和pIB139从供体菌E.coli ET12567(pUZ8002)导入到链霉菌211726的新鲜菌丝体和孢子中,结果表明,在链霉菌211726菌株萌发孢子作为受体菌与含有质粒pSET152和pIB139的大肠杆菌E.coli ET12567(pUZ8002)菌株进行两亲接合转移的实验中,能够获得阿泊拉霉素的抗性菌落;而在链霉菌211726菌株新鲜菌丝体作为受体菌与含有质粒pSET152和pIB139的大肠杆菌E.coli ET12567(pUZ8002)菌株进行的两亲接合转移的实验中,没有得到阿泊拉霉素的抗性菌落。
2.3 供体菌和受体菌的比例
为了确定接合转移过程中受体菌和供体菌接种最佳比例,不同比例的大肠杆菌和链霉菌211726孢子进行混合涂布平板。结果(表2)表明,当大肠杆菌与孢子的比例达到8∶1时,获得的接合转移子数最多。
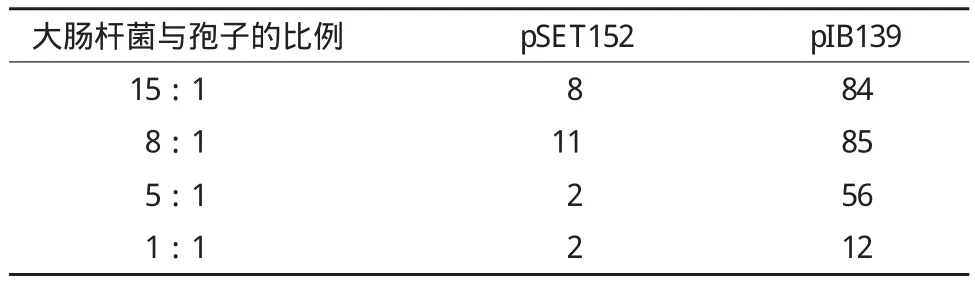
表2 基于链霉菌孢子的接合转移平板统计
2.4 接合转移子的验证
以CP1和CP2为引物进行PCR扩增验证,并以出发菌株链霉菌211726基因组DNA的扩增结果作为阴性对照,质粒pSET152和pIB139的扩增结果作为阳性对照,扩增结果(图1)显示,所获得的2株接合转移子都可以扩增出预测的882 bp条带。分别回收2 株接合转移子扩增条带,TA克隆后送测序,测序结果也证实了2株皆为阳性克隆子。

图1 来自pIB139和pSET152接合转移子的PCR验证
2.5 质粒pIB139和pSET152在链霉菌211726接合转移子中的稳定性
为了检测质粒pIB139和pSET152在链霉菌211726菌株中的稳定性,随机挑取验证正确的 200个接合转移子,发现它们在添加有阿泊拉霉素和无抗生素的SFMS培养基平板上都能生长,没有发现阿泊拉霉素抗性消失的菌落。结果表明质粒pIB139和pSET152在链霉菌211726菌株中可稳定存在。
3 讨论
将外源DNA导入宿主细胞的方式主要有:钙转化、接合转移、电转化和显微注射等。其中,接合转移因导入效率较高、高度跨种属差异、价格低廉、操作方便等优势被广泛应用于链霉菌研究领域。
质粒是一种闭合环状双链DNA[17],它不属于基因组DNA,游离于染色体DNA之外。在链霉菌遗传操作中人们常将其作为载体来介导外源DNA向宿主细胞的导入。载体可依据其是否能整合到链霉菌染色体DNA上的特点分为整合型载体和非整合型载体,例如,整合型质粒pIB139和pSET152[18,19]。而非整合型载体又可依据其是否含有链霉菌复制子分为自杀型载体和游离型载体[20],例如游离型载体pYH7[21,22]。
在接合转移过程中,含有外源载体的供体菌(大肠杆菌)和受体菌链霉菌在同一个平板上生长时,两者之间存在不可避免的相互竞争。由于链霉菌的生长速度比大肠杆菌的生长速度缓慢,过量的大肠杆菌会使链霉菌的生长受到抑制,导致接合转移率降低。因此,在建立一个新菌株的遗传操作系统时摸索供体菌与受体菌之间的最佳比例就尤为重要。
4 结论
本研究通过对链霉菌211726抗生素敏感性测定和基于孢子和菌丝体接合转移系统的摸索,成功地将位点特异性整合型载体质粒pIB139和pSET152整合到链霉菌211726染色体中,在分子水平证明所获得的接合转移子的正确性,这说明本研究建立的遗传操作系统是有效可行的。
[1] Wakesman SA, Heriei AT. The nomen culture and classification of the Actinoymcetes[J]. J Bacteriol, 1943, 46:337-341.
[2] Kieser HM, Kieser T, Hopwood DA. A combined genetic and physical map of the Streptomyces coelicolor A3(2)chromosome[J]. J Bacteriol, 1992, 174:5496-5507.
[3] Goodfellow M, Cross T. The Biology of the Actinomyces[M]. London:Academic Press, 1984.
[4] Franco CM, Coutinho LE. Detection of novel secondary metabolites[J]. Crit Rev Biotechnol, 1991, 11:193-276.
[5] Davies J. What are antibiotics? Archaic functions for modern activities[J]. Mol Microbiol, 1990, 4:1227-1232.
[6] Morin RB, Gorman M. Chemistry and biology of beta-lactam antibiotics[M]. New York:Academic Press, 1982.
[7] Strynadka NC, Adachi H, Jensen SE, et al. Molecular structure of the acyl-enzyme intermediate in beta-lactam hydrolysis at 1. 7 A resolution[J]. Nature, 1992, 359:700-705.
[8]Arai M. Azalomycins B and F, two new antibiotics. I. Production and isolation[J]. The Journal of Antibiotics, 1960, 13:46.
[9]Krystofova S, Varecka L, Betina V. The uptake by trichoderma viride Mycelium. correlation with growth and conidiation[J]. General Physiology and Biophysics, 1995, 14:323-338.
[10] Yuan G, Hong K, Lin H, et al. New azalomycin F analogs from mangrove Streptomyces sp. 211726 with activity against microbes and cancer cells[J]. Marine Drugs, 2013, 11:817-829.
[11] Yuan G, Lin H, Wang C, et al. 1H and 13C assignments of two new macrocyclic lactones isolated from Streptomyces sp. 211726 and revised assignments of azalomycins F3a, F4a and F5a[J]. Magnetic Resonance in Chemistry:MRC, 2011, 49:30-37.
[12] Yuan GJ, Hong K, Lin HP, Li J. Azalomycin F4a2-ethylpentyl ester, a new macrocyclic lactone, from mangrove actinomycete Streptomyces sp. 211726[J]. Chinese Chemical Letters, 2010, 21:947-950.
[13]Yuan GJ, Li PB, Yang J, et al. Anti-methicillin-resistant Staphylococcus aureus assay of azalomycin F5aand its derivatives[J]. Chinese Journal of Natural Medicines, 2014, 12:309-313.
[14]Fukushima K, Arai T, Iwasaki S, et al. Studies on macrocyclic lactone antibiotics. VI. Skeletal structure of copiamycin[J]. J Antibiot(Tokyo), 1982, 35:1480-1494.
[15]袁干军. 产大环内酯化合物的红树林放线菌的筛选、产物分离鉴定及活性研究[D]. 海口:海南大学, 2010:1-185.
[16]Tobias K, Bibb MJ, Mark JB, et al. Practical Streptomyces Genetics[M]. Norwich:The John Innes Foundation, 2000.
[17]Mens T, Demeyer S, Du Bois B, et al. Refactoring:Current research and future trends[J]. Electronic Notes in Theoretical Computer Science, 2003, 82(3):483-499.
[18]Wilkinson CJ, Hughes-Thomas ZA, Martin CJ, et al. Increasing the efficiency of heterologous promoters in actinomycetes[J]. J Mol Microbiol Biotechnol, 2002, 4(4):417-426.
[19]Del Vecchio F, Petkovic H, Kendrew SG, et al. Active-site residue, domain and module swaps in modular polyketide synthases[J]. J Ind Microbiol Biotechnol, 2003, 30(8):489-494.
[20]Gerdes K, Rasmussen PB, Molin S. Unique type of plasmid maintenance function:postsegregational killing of plasmid-free cells[J]. Proceedings of the National Academy of Sciences, 1986, 83(10):3116-3120.
[21] Sun Y, He X, Liang J. Analysis of function in plasmid pHZ1358 influencing its genetic and structural stability in Streptomyces lividans 1326[J]. Applied Microbiology and Biotechnoogy, 2009, 82:303-310.
[22]Sun Y, Zhou X, Liu J, et al. Streptomyces nanchangensis, a producer of the insecticidal polyether antibiotic nanchangmycin and the antiparasitic macrolide meilingmycin, contains multiple polyketide gene clusters[J]. Microbiology, 2002, 148:361-371.
(责任编辑 李楠)
The Construction of the Gene Transfer System of Strain Streptomyces sp. 211726 Producing Azalomycin F
MA Yan-ling1LIU Fu-lai1ZHANG Min1SUN Yu-hui2HONG Kui2
(1. College of Food and Horticultural Sciences,Foshan University,Foshan 528231;2. School of Pharmaceutical Sciences,Wuhan University,Wuhan 430071)
The objective of this study is to establish the gene transfer system of strain Streptomyces sp. 211726 producing azalomycin F,which can be used for genetic manipulations such as gene knock-out and expression of foreign genes. Intergeneric genetic transfer system of Streptomyces sp. 211726 producing azalomycin F was constructed by conjugating integrative plasmid pSET152 with pIB139. Results showed that 25 mg/mL apramycin may be used to efficiently screen conjugants. PCR verification revealed that exogenous plasmid was successfully integrated in the chromosomal DNA of Streptomyces sp. 211726. The continuous passage culture experiment demonstrated that transformed pSET152 and pIB139 of conjugants were stably inherited.
azalomycin F;Streptomyces sp. 211726;conjugation;genetic stability
10.13560/j.cnki.biotech.bull.1985.2016.04.026
2015-07-23
国家自然科学基金项目(31270120),佛山市科技计划项目(2014GA000425,2014AG10007)
马艳玲,女,博士,研究方向:食品生物技术;E-mail:mayanling100@163.com
猜你喜欢
当代水产(2022年1期)2022-04-26
世界最新医学信息文摘(2021年12期)2021-06-09
医学食疗与健康(2021年25期)2021-05-12
四川蚕业(2020年4期)2020-02-10
兵团工运(2017年7期)2017-01-26
画刊(2016年2期)2016-11-01
中外医疗(2015年16期)2016-01-04
少儿科学周刊·少年版(2015年3期)2015-07-07
少儿科学周刊·少年版(2015年3期)2015-07-07
食品工业科技(2014年15期)2014-03-11
