
密度对三种莎草科植物克隆生长的影响
2016-06-05 15:01钟青龙戴文龙项世亮王天厚汪承焕
生态科学 2016年1期
钟青龙, 戴文龙, 项世亮, 王天厚, 汪承焕
华东师范大学生命科学学院, 上海城市化生态过程与生态恢复重点实验室, 上海 200241
密度对三种莎草科植物克隆生长的影响
钟青龙, 戴文龙, 项世亮, 王天厚, 汪承焕*
华东师范大学生命科学学院, 上海城市化生态过程与生态恢复重点实验室, 上海 200241
以三种莎草科克隆植物藨草(Scirpus triqueter), 海三棱藨草(Scirpus mariqueter)和糙叶苔草(Carex scabrifolia)为研究对象, 通过盆栽受控实验, 探讨三种初始种植密度处理(1株/盆、4株/盆和16株/盆)对个体表现与种群增长的影响。结果表明, 三种植物种群经过一个月左右的适应后均开始加速增长, 生长季后期增长率逐渐降低, 而海三棱藨草种群甚至出现了一定程度的衰退。生长季中前期藨草在中低密度处理下密度增长率高于高密度和整簇种植处理, 而海三棱藨草、糙叶苔草种群的整体密度增长与初始种植密度成反比。最终收获时, 藨草和海三棱藨草不同处理下的密度、冠层高度、最高高度、地上生物量和地下生物量均没有显著差异, 糙叶苔草在低种植密度下的密度最低, 中等种植密度下冠层高度最高。藨草、海三棱藨草的单株平均生物量与密度呈显著负相关, 而藨草、糙叶苔草的总生物量与密度呈显著正相关, 三种莎草科植物的根冠比都没有随密度发生显著变化。三种莎草科植物在高密度下的种内竞争效应均强于中密度。
种群增长; 密度效应; 藨草; 海三棱藨草; 糙叶苔草
1 前言
在相对较高密度下, 竞争作用不仅可以通过降低出生率, 增加死亡率的方式影响植物种群动态, 而且在个体水平上可以改变植物生长速度和形态[1]。克隆植物(clonal plant)是指在自然生境条件下, 能够通过营养生长过程, 形成多个在遗传上一致、在形态和生理上独立或者潜在独立的个体的一类植物[2]。随着种内竞争的加剧, 克隆植物构件在数量、空间排列和形态结构等方面都会发生变化, 很多研究都反映了克隆植株性状对密度的响应模式。结缕草(Zoysia japonica)无性系构件在不同密度下采取不同的适应对策: 低密度下迅速占领空间, 分蘖数量和单株产生的匍匐茎数量、匍匐茎总长度、节间长度和节数增加; 高密度下则减缓生长, 以降低种内构件间的竞争[3]。密度效应对生物量的分配也有影响,红葱(Allium cepa var. proliferum)地上部分、叶和鞘的生物量分配比例均随密度的增加而增加, 地下部分和鳞茎的分配比例随密度的增加而下降, 而根的分配比例未随密度发生显著变化[4]。然而, 我们对于克隆植物在有限空间下的种群动态是否会受到密度调控, 以及这种调控作用对个体表现及种内竞争的影响的了解仍比较有限。
藨草、海三棱藨草和糙叶苔草是崇明东滩几种重要的莎草科土著植物, 藨草和海三棱藨草属于先锋物种, 常常形成混生群落, 糙叶苔草伴生于海三棱藨草(或藨草)群落内及芦苇群落的边缘地带[5]。尽管已有研究探讨了盐度、水位对这几个物种个体生长与种间竞争的影响[6], 但我们对于密度在其种群增长以及种内竞争中的效应仍缺乏足够的了解。本文通过盆栽受控实验, 研究初始种植密度对三种莎草科植物种群增长与个体表现的影响, 以探寻克隆植物的种内调节及其适应机制, 帮助我们更好地理解其种群动态, 为崇明东滩等盐沼湿地生态系统的保护提供理论依据。
2 材料与方法
2.1 实验材料
藨草是莎草科藨草属多年生草本植物, 分布广泛, 生长于河口滩涂、沟渠、湖泊、池塘、河流等处, 同时也是一种稻田杂草[7]。藨草的繁殖能力及适应性强, 主要以球茎进行无性繁殖。海三棱藨草是莎草科藨草属多年生草本植物, 为我国特有种, 分布于长江口及杭州湾沿岸滩涂, 可以种子及根茎、球茎繁殖, 在生长季末, 地上植株全部死亡而地下部分能够存活越冬[8]。糙叶苔草是莎草科苔草属多年生草本植物, 主要生长于滩涂高潮位地带, 生长区域较干燥, 不耐水淹, 群落盖度可达50%以上[9]。
2.2 实验设计
2013年4月, 在崇明东滩挖取藨草、海三棱藨草和糙叶苔草的无性系分株带回实验室在淡水中培养。一周后, 挑选相近大小的分株(藨草: 约5 cm高,海三棱藨草: 约8 cm高, 糙叶苔草: 约12 cm高),移栽至直径20 cm、深度30 cm的塑料花盆中。花盆内盛有等量淘洗干净的河沙, 桶底置有纱网, 防止河沙从透水孔中流失。每盆添加奥绿缓释肥(Osmoeote, N/P/K14:14:14, Scotts公司)2 g, 与沙混合均匀。花盆放置在71 cm*45 cm*18 cm的周转箱中, 保持水位与基质表面平齐。三个物种单独种植, 初始种植密度有1株/盆, 4株/盆和16株/盆三种处理, 另外, 藨草增加一种处理: 将整簇16株根部相连的分株一起种植在1个花盆中。每种处理4个重复, 共40 盆。两周内对死亡的分株进行了补种。分别于5月15日、5月31日、6月15日、6月30日、7月15日、7月31日、8月15日、8月31日、9月15日、9月30日和10月15日对所有物种的密度、冠层高度进行测量, 共测量11次。10月29日收获所有植株, 测量地上部分冠层高度和最高高度, 计数每一盆内的分株数和果实数量, 将地上、地下部分分离后带回实验室, 置于80 ℃烘箱内烘至恒重后测定生物量。
2.3 数据分析
种群增长率的计算公式为: (Dn+1-Dn)/ Dn*100%, Dn为每次测量的种群密度。采用重复测量的方差分析(Repeated measurement ANOVA)检验初始种植密度对三种植物密度、高度生长的效应。采用单因素方差分析检验初始种植密度对最终收获时三种植物的冠层高度、最高高度、密度、地上生物量、地下生物量和每桶果实数量的效应, 并运用Tukey’s test进行多重比较。采用一元线性回归分析收获时三种植物总生物量、根冠比、平均单株生物量、结实率与密度的关系。采用相对邻株效应(relative neighbor effect, RNE)来度量中密度、高密度以及整簇种植处理下各物种的种内竞争强度, 计算公式为: RNE =(M-C)/max (M, C), 其中M是有相邻个体时(即分别在中密度、高密度及整簇种植处理下)植株的平均生物量, C是没有相邻个体时(即在低密度处理下)植株的平均生物量[10]。相对邻株效应(RNE)数值为负表示植株间为负相互作用关系(竞争作用), 且绝对值越大, 竞争越强; RNE的数值为正表示植株间为正相互作用关系(促进作用), 且绝对值越大, 促进作用越强[10]。所有统计分析均采用SPSS 19进行, 显著性水平为0.05。所有图形均用SigmaPlot 10.0绘制。
3 结果与分析
3.1 初始密度对三种莎草科植物密度、高度增长的影响
藨草和糙叶苔草密度经过一个月左右的迟滞期后开始逐渐加速增长, 直至生长季末达到最高(图1a,图1e)。海三棱藨草的密度增长在生长季中前期与藨草、糙叶苔草相似, 但在末期出现了一定程度的衰退(图1c)。总体而言, 三种植物种群都经过一个月左右的调整后开始加速增长, 而生长季后期增长率逐渐降低, 藨草、海三棱藨草早期的增长率高于糙叶苔草(图2)。在整个生长季中, 不同初始种植密度处理下藨草的密度增长没有显著差异, 而海三棱藨草和糙叶苔草则有极显著差异(p<0.01)(表1)
生长季中前期藨草在中低密度处理下密度增长率高于高密度和整簇种植处理, 但在生长季后期这种差异基本消失(图 2a), 不同处理下种群的密度增长趋势基本相同(图 1a)。海三棱藨草、糙叶苔草种群的整体密度增长率与初始种植密度成反比(图 2b,图2c); 到生长季末期, 海三棱藨草不同处理下的种群密度趋于一致(图 1c), 而糙叶苔草低密度处理下的种群密度始终低于其他两个处理(图1e)。
藨草的高度在生长季中前期增长较快, 随后高度趋于稳定并略有下降(图 1b)。海三棱藨草高度在生长季初期快速增长, 七月中旬后增长变缓, 甚至略有下降, 之后继续缓慢增长, 直至生长季末(图1d)。整个生长季中, 糙叶苔草不同处理下平均高度相近,并保持相似的稳定增长趋势(图 1f)。整个生长季中,三种克隆植物不同处理下的高度增长均没有显著差异(表1)。
在生长季早期, 藨草的高度增长率为低密度、中密度处理大于高密度处理; 海三棱藨草的高度增长率为低密度处理大于中密度、高密度处理; 生长季中、后期, 海三棱藨草、糙叶苔草各个处理的高度增长率相近。
3.2 初始种植密度对三种莎草科植物生长指标的影响
生长季末收获时, 藨草和海三棱藨草不同处理下的密度、冠层高度、最高高度、地上生物量和地下生物量均没有显著差异(图3)。糙叶苔草低密度处理下的密度显著低于中密度处理和高密度处理(p<0.05)(图3a), 而冠层高度则是中密度处理显著高于低密度处理和高密度处理(p<0.05)(图3b)。
3.3 三种莎草科植物生长繁殖参数与密度的相关性
生长季末收获时, 藨草的单株平均生物量与密度呈显著负相关(p<0.05), 而总生物量与密度呈显著正相关(p<0.05)(图4a, 图4b)。海三棱藨草的单株生物量与密度呈显著负相关(p<0.05), 总生物量在不同密度下基本维持恒定(图 4c, 图 4d)。糙叶苔草的单株生物量基本不随密度变化, 而总生物量随密度升高而显著增长(p<0.05)(图4e, 图4f)。随着密度的增加, 三种克隆植物的根冠比没有表现出显著的线性变化规律。生长季结束时, 海三棱藨草和糙叶苔草没有结实, 而藨草的结实率与密度之间也没有显著相关性。
3.4 不同初始密度处理下三种莎草科植物的种内竞争强度
藨草、海三棱藨草、糙叶苔草中密度、高密度处理下的相对邻株效应 (RNE) 值均小于 0, 且高密度处理下相对邻株效应(RNE)的绝对值显著大于中密度处理(p<0.05)。整簇种植下藨草的相对邻株效应(RNE)值与高密度处理没有显著差异(图5)。
4 讨论
藨草、海三棱藨草和糙叶苔草三种克隆植物都具有很强的无性繁殖能力, 但其生物学特性存在差异。藨草分株密集生长在基株周围, 间距小, 能够达到很高的密度; 海三棱藨草主要由横向生长的地下茎节形成分株, 间距较大, 其种群的密度制约效应更强, 生长季后期出现了自疏; 糙叶苔草在移栽后的调整适应较慢, 持续了近两个月时间, 生长季末期仍未达到最高密度。尽管如此, 初始种植密度对这几个物种种群动态的调控, 以及对个体表现及种内竞争的影响模式仍十分相近。

图1 三种莎草科植物的密度和高度增长趋势Fig. 1 Density and height growth of three sedge plants
4.1 克隆植物的增长规律及其密度调节
在整个生长季中, 随着密度的增加, 三种克隆植物的密度增长率均表现出先增加后降低的趋势。藨草的整体增长率较高, 中、低密度处理下很早变达到生长高峰, 并能维持较长时期的稳定增长; 海三棱藨草和糙叶苔草的种群增长率都在八月达到最大值, 之后种群增长率明显降低, 海三棱藨草在生长季末期甚至出现了负增长。尽管海三棱藨草的种群动态与张利权和雍学葵[11]的研究结果相似, 但两者的原因并不完全一致。野外条件下, 海三棱藨草在生长季中后期(八、九月间)的衰退主要是由于受到台风季潮汐侵蚀作用的影响[11]。而在我们的盆栽实验中, 海三棱藨草种群衰退的主要原因是密度制约引起的死亡率升高, 在中、高密度处理下, 分株间的竞争更为激烈, 因而生长季末期其死亡率远高于低密度处理。对其他克隆植物的研究发现, 生长于开阔空间的初始源株可以更好地生存并产生新分株,而源株拥挤会导致竞争,使新分株产生及存活率下降[12-13]。本实验中从种植到收获, 藨草和海三棱藨草在低密度处理下的种群增长率整体上高于高密度
处理, 最终收获时种群密度没有显著差异, 表明藨草和海三棱藨草较高的初始密度会限制分株增长。王永健和钟章成[14]在模拟源株密度对蝴蝶花(Iris japonica)生长和克隆繁殖影响的研究中也发现, 1个起始源株处理下蝴蝶花新分株数显著高于2个起始源株及 4个起始源株处理, 而新分株死亡率显著低于后二者; 随着起始源株数增加(种内竞争增强), 一级与二级子株数显著降低, 随着密度增加, 新分株减少, 分株死亡增加, 这些都充分体现了种群增长的密度效应。

表1 不同初始密度处理对三种莎草科植物密度和高度增长影响的重复测量的方差分析结果Tab. 1 Results of repeated measurement ANOVA testing the effects of different initial density treatment on the density and height growth of three sedge plants

图2 不同初始密度处理对三种莎草科植物种群增长率的影响Fig. 2 Effects of different initial density treatment on the population growth rate of three sedge plants
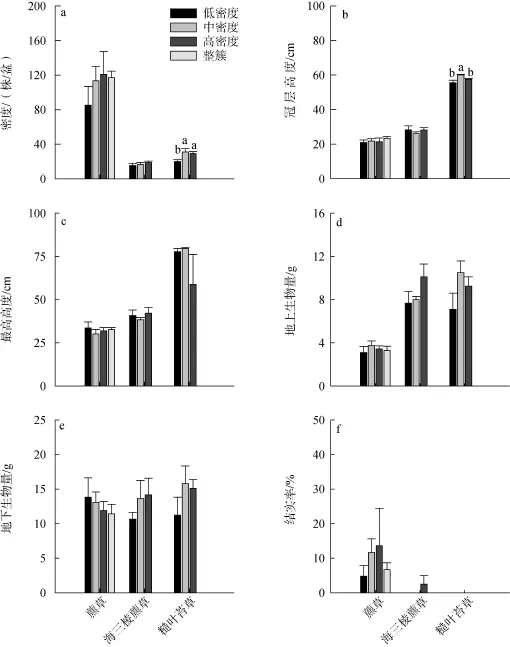
图3 收获时不同初始密度处理下三种莎草科植物的生长繁殖表现Fig. 3 Growth and reproduction performances of three sedge plants at harvest under different initial density treatments
许多研究表明, 克隆种群有很强的密度调控能力。高密度的紫茎泽兰(Eupatorium adenophorum)种群在强烈竞争下, 基株会控制根蘖株的形成和死亡,从而减少资源的消耗, 保证种群对生境的长期占领[15]。本实验中, 藨草和糙叶苔草种群经过初期的适应之后, 增长率明显增高, 有利于种群充分利用资源,快速扩张; 后期高密度下增长率降低至趋近于零的水平, 使种群能够维持在环境容纳量之下, 又避免了自疏造成的资源浪费。这表明克隆植物可以通过数量调节维持种群稳定, 促进生态平衡。

图4 三种莎草科植物收获时生物量与密度的关系Fig. 4 Relationships between biomass and density at harvest of three sedge plants
盐沼生态系统环境梯度明显, 物种组成较为简单, 因而是研究植物群落组织机制的良好系统。何彦龙研究崇明东滩中低潮滩盐沼植被分异的形成机制时发现, 互花米草(Spartina alterniflora)相对高生长速率帮助其更多地从生境中获取营养物质, 从而减少了海三棱藨草可以利用的资源, 限制了海三棱藨草种群的发展[16]。对于滩涂前沿的三种莎草科先锋植物(藨草、海三棱藨草和糙叶苔草)而言, 对盐度、淹水等环境因子耐受能力的差异是决定其分布区域的主要因素[6]。然而, 在更小的尺度上, 对生境的优先占据及种群动态的差异对混生群落的物种组成起着更重要的作用。本研究表明, 藨草在整个生长季中维持较高的增长率, 最终达到很高的分株密度, 在适宜其生长的低盐度生境中难以被另两个物种入侵; 海三棱藨草早期的增长率较高, 但生长季后期种群衰退时可能被仍处于稳定增长期的藨草及糙叶苔草入侵, 在高盐度、淹水深的小生境中占有竞争优势; 糙叶苔草在生长季早期增长缓慢, 不利于其抢先占据空白生境, 但中后期稳定持续的生长将更有利于其与演替后期物种的竞争。这些物种间拓殖、生长能力的差异会对滩涂前沿莎草科先锋植物群落的组成和结构产生重要影响。

图 5 不同初始密度处理对三种莎草科植物相对邻株效应(RNE)的影响Fig. 5 Effects of different initial density treatment on the relative neighbor effect (RNE) of three sedge plants
4.2 种群密度对克隆植物种内竞争的影响
本实验结果显示, 三种克隆植物在中密度和高密度处理下植株间竞争作用占主导, 高密度处理下相对邻株效应(RNE)的绝对值均显著高于中密度处理, 表明种植密度越高的处理整体上竞争效应越强。于国磊[17]在研究水淹对克隆植物空心莲子草(Alternanthera philoxeroides)种内关系的影响时得出了相似的结论, 相对于低密度处理(不存在种内竞争), 空心莲子草的生长在中密度和高密度下显著减弱, 其植株之间整体上为竞争关系。
很多研究表明, 克隆植物的内部竞争对种群增长及其个体生长有着同样重要的影响。如种内竞争会降低紫茎泽兰个体株高、叶片数、根蘖株数和生物量的增长[15]。在资源水平相对一致时, 羊草(Leymus chinensis)克隆构型特征, 如根茎节间长度、间隔子长度、分枝距离和分枝角度等随种群密度变化有较强的可塑性[18]。总之, 随着克隆植物种群密度的改变以及种内竞争的加剧, 植株生长策略以及种群调节等方面都会产生变化。这些变化在本实验中也有体现: 生长季早期, 藨草低、中密度处理的高度增长率大于高密度处理, 海三棱藨草的高度增长率为低密度处理大于中密度、高密度处理。
值得注意的是, 植株间的竞争强度除了受密度的影响外, 分株间的紧密联系也有利于克隆植物进行整体协作[19], 进而减弱竞争。然而, 本实验高密度处理与整簇处理的竞争强度没有显著差异, 可能是由于实验没有设置异质化的条件, 在同质环境中,分株间劳动分工及资源共享的价值丧失[20-21]。另一方面, 藨草最高的初始种植密度仍然远低于其能够达到的最大密度, 因而在生长过程中, 基株的调控效应较弱, 使整簇种植与高密度种植处理间有相似的生长表现。
4.3 种群密度与生长指标之间的关系
本研究结果表明, 不同物种的种群密度与生长指标间的关系存在差异。在现有资源供应条件下,不同密度下海三棱藨草种群的总生物量相对恒定,但密度增高后分配给单株的生物量降低; 糙叶苔草在高密度下总生物量仍没有达到环境容纳量, 分株都能获得足够的资源生长到相近的大小。藨草总生物量也可能还未达到环境容纳量, 但其策略与糙叶苔草不同, 藨草通过产生大量的小分株充分占据生境利用资源, 基株对分株形成的调控能力相对较弱,因而平均分株生物量随密度升高而下降。
收获时, 藨草、海三棱藨草的单株生物量与测定区间内的密度呈显著负相关, 这是高密度下种内竞争对个体大小的影响。实际上, 种内关系不仅有竞争还有促进。在相对不利的环境条件下, 当种群处于较低密度时, 正相互作用的效应要大于竞争,个体生物量与密度会呈正相关关系; 而密度升高到资源竞争效应占主导时, 个体生物量与密度则转变为负相关关系[22-23]。随着非生物胁迫的增加, 只考虑竞争时平均个体生物量与密度的关系呈单调递减,而在考虑了促进关系时, 平均个体生物量与密度的关系呈单峰型曲线[22]。本实验中, 由于不存在资源胁迫以及密度测量区间的限制, 单株生物量与密度之间没有体现出显著正相关(单调递增)关系或者单峰型曲线。
另一方面, 种内竞争水平会对植物体资源分配产生较大影响, 当竞争强度增加时, 植株各个部位的生长会受到不同程度的限制, 使植物的生物量分配格局在不同密度下产生变化。蝴蝶花地下部分分配比例在低密度处理下更高, 而高密度处理下地上部分分配比例更高[14], 是由于高密度下的竞争限制了植株生长, 源株间竞争的同时促进了地下根茎生长的竞争, 使根生长量降低[24]。与此相类似, 随着密度的增加, 红葱个体会增加地上营养器官的生物量分配, 并以减小地下无性繁殖器官的生物量分配为代价[4]。所以植物地上地下部分的竞争强度及植株采取的适应策略都可能导致植株生物量分配格局的改变。本实验中的三种莎草科植物在不同密度处理下根冠比未发生显著变化, 可能是由于未设置不同资源供应的处理, 导致植株地上、地下部分的竞争强度差异不大。
5 结论
本实验主要揭示了克隆植物的种内竞争关系:高密度下的竞争限制了种群生物量, 降低了单株生物量, 制约了种群的增长。藨草、海三棱藨草和糙叶苔草是长江口盐沼先锋植物群落中的优势物种,除了对盐度、淹水等环境因子耐受能力的差异之外,三种莎草科植物不同的生长速率和种内竞争强度可能也是决定其分布的重要因素。对于克隆植物在滩涂环境中的增长规律以及生长策略的深入研究, 将有助于我们预测这一区域植物的种群动态, 控制生物入侵, 维护生态系统稳定性。
[1] 段仁燕, 王孝安. 太白红杉种内和种间竞争研究[J]. 植物生态学报, 2005, 29(2): 242-250.
[2] COOK R E. Growth and development in clonal population[M]//Jackson J B C, Buss L W, Cook R E. Population Biology and Evolution of Clonal Organisms. New Haven: Yale University Press, 1985: 259-296.
[3] 代保清, 王艳. 密度对结缕草无性系构件生长的影响[J].沈阳师范大学学报(自然科学版), 2013, 31(3): 356-358.
[4] 黎磊, 周道玮, 盛连喜. 密度制约决定的植物生物量分配格局[J]. 生态学杂志, 2011, 30(8): 1579-1589.
[5] 王卿. 长江口盐沼植物群落分布动态及互花米草入侵的影响[D]. 上海: 复旦大学, 2007: 26-37.
[6] 汪承焕. 环境变异对崇明东滩优势盐沼植物生长、分布与种间竞争的影响[D]. 上海: 复旦大学, 2009: 48-62.
[7] DEEGAN B, HARRINGTON T J, DUNDON P. Effects of salinity and inundation regime on growth and distribution of Schoenoplectus triqueter[J]. Aquatic Botany, 2005, 81(3):199-211.
[8] SUN Shucun, GAO Xianming, CAI Yongli. Variations in sexual and asexual reproduction of Scirpus mariqueter along an elevational gradient[J]. Ecological Research, 2001, 16(2): 263-274.
[9] 上海科学院. 上海植物志(下卷): 经济植物[M]. 上海:上海科学技术文献出版社, 1999: 527-528.
[10] KIKVIDZE Z, KHETSURIANI L, KIKODZE D, et al. Seasonal shifts in competition and facilitation in subalpine plant communities of the central Caucasus[J]. Journal of Vegetation Science, 2006, 17(1): 77-82.
[11] 张利权, 雍学葵. 海三棱藨草种群的密度与生物量动态[J]. 植物生态学与地植物学学报, 1992, 16(4): 317-325.
[12] OBORNY B, KUN A, CZÁRÁN T, et al. The effect of clonal integration on plant competition for mosaic habitat space[J]. Ecology, 2000, 81(12): 3291-3304.
[13] VAN KLEUNEN M, FISCHER M, SCHMID B. Effects of intraspecific competition on size variation and reproductive allocation in a clonal plant[J]. Oikos, 2001, 94(3): 515-524.
[14] 王永健, 钟章成. 模拟源株密度对蝴蝶花生长和克隆繁殖的影响[J]. 植物生态学报, 2010, 34(3): 340-347.
[15] 赵春燕, 沈有信. 紫茎泽兰种内竞争试验[J]. 生态与农村环境学报, 2008, 24(2): 27-31.
[16] 何彦龙. 中低潮滩盐沼植被分异的形成机制研究—以崇明东滩盐沼为例[D]. 上海:华东师范大学, 2013: 54-73.
[17] 于国磊. 水淹对克隆植物空心莲子草种内关系的影响[J].植物生态学报, 2011, 35(9): 973-980.
[18] 何念鹏, 吴泠, 周道玮. 松嫩草地羊草克隆构型特征在不同种群密度下的可塑性[J]. 应用与环境生物学报, 2005, 11(2): 152-155.
[19] 张大勇. 植物生活史进化与繁殖生态学[M]. 北京: 科学出版社, 2004: 62-64.
[20] ALPERT P, MOONEY H A. Resource sharing among ramets in the clonal herb, Fragaria chiloensis[J]. Oecologia, 1986, 70(2): 227-233.
[21] ALPERT P. Clonal integration in Fragaria chiloensis differs between populations: ramets from grassland are selfish[J]. Oecologia, 1999, 70: 227-233.
[22] CHU Chengjin, MAESTRE F T, XIAO Sa, et al. Balance between facilitation and resource competition determines biomass-density relationships in plant populations[J]. Ecology Letters, 2008, 11(11): 1189-1197.
[23] 贾昕. 基于影响域模型的植物间相互作用沿环境梯度的变化规律及其对种群动态调控的研究[D]. 杭州:浙江大学, 2011: 56-80.
[24] CHEPLICK G P, GUTIERREZ C M. Clonal growth and storage in relation to competition in genets of the rhizomatous perennial Amphibromus scabrivalvis[J]. Canadian Journal of Botany-revue canadienne de botanique, 2000, 78(4): 537-546.
Effects of planting density on clonal growth of three sedge plants
ZHONG Qinglong, DAI Wenlong, XIANG Shiliang, WANG Tianhou, WANG Chenghuan*
Shanghai Key Laboratory of Urbanization & Ecological Restoration, School of Life Science, East China Normal University, Shanghai 200241, China
This study examined the effects of three initial planting density treatments (1, 4 and 16 ramets per pot) on individual and population growth of Scirpus triqueter, Scirpus mariqueter and Carex scabrifolia using a controlled environment experiment. Our result showed that the population size of these three species increased rapidly after an adaptation period of about one month, the growth rate decreased gradually in late growing season and S. mariqueter even declined during this period. Density growth rate of S. triqueter in early growing season was higher in low and medium initial planting density treatments than that in high density and cluster planting treatments. Population growth of S. mariqueter and C. scabrifolia had a negative correlation with initial planting density. At harvest, ramet density, canopy height, height of the tallest ramet, above- and belowground biomass of S. triqueter and S. mariqueter did not differ significantly among the three initial planting density treatments. However, for C. scabrifolia, the low initial density treatment had the lowest final density, while canopy height was the highest in the medium initial density treatment. Single ramet biomass of both S. triqueter and S. mariqueter had significantly negative correlations with their ramet density, while the total biomass of S. triqueter and C. scabrifolia was significantly and positively correlated with their ramet density. Root:shoot ratio of these species did not differ significantly with increasing ramet density at harvest, and they had more intense intraspecific competition in high density treatment than in medium density treatment.
population growth; density effect; Scirpus mariqueter; Scirpus triqueter; Carex scabrifolia
10.14108/j.cnki.1008-8873.2016.01.001
Q145
A
1008-8873(2016)01-001-09
2015-01-30;
2015-02-16
国家自然科学基金项目(31100317); 上海市科委科研计划项目(12231204703); 上海市科委科技创新行动计划(14DZ1206003)
钟青龙(1988—), 男, 硕士, 研究方向为湿地植物生态学, E-mail: 359114054@qq.com
*通信作者:汪承焕, 男, 博士, 助理研究员, 主要从事湿地生态学研究, E-mail: chwang@bio.ecnu.edu.cn
钟青龙, 戴文龙, 项世亮, 等. 密度对三种莎草科植物克隆生长的影响[J]. 生态科学, 2016, 35(1): 1-9.
ZHONG Qinglong, DAI Wenlong, XIANG Shiliang, et al. Effects of planting density on clonal growth of three sedge plants[J]. Ecological Science, 2016, 35(1): 1-9.
猜你喜欢
环球时报(2022-09-20)2022-09-20
资源信息与工程(2021年5期)2022-01-15
今日农业(2020年24期)2020-12-15
今日农业(2020年19期)2020-12-14
四川地质学报(2020年2期)2020-05-31
兽医导刊(2016年12期)2016-05-17
中国房地产业(2016年8期)2016-03-01
中国科技信息(2015年2期)2015-11-16
现代检验医学杂志(2015年4期)2015-02-06
植物营养与肥料学报(2014年1期)2014-03-11
