
秋季南沙群岛海域浮游纤毛虫群落与环境因子关系的初步研究
2016-05-25 00:37吴风霞黄洪辉谭烨辉齐占会
中山大学学报(自然科学版)(中英文) 2016年5期
吴风霞, 黄洪辉,2, 谭烨辉, 齐占会
(1.广东省渔业生态环境重点实验室//农业部南海渔业资源开发利用重点实验室∥中国水产科学研究院南海水产研究所, 广东 广州 510300;2. 南海生物资源开发与利用协同创新中心, 广东 广州 510300;3. 中国科学院南海海洋研究所, 广东 广州510301)
秋季南沙群岛海域浮游纤毛虫群落与环境因子关系的初步研究
吴风霞1, 黄洪辉1,2, 谭烨辉3, 齐占会1
(1.广东省渔业生态环境重点实验室//农业部南海渔业资源开发利用重点实验室∥中国水产科学研究院南海水产研究所, 广东 广州 510300;2. 南海生物资源开发与利用协同创新中心, 广东 广州 510300;3. 中国科学院南海海洋研究所, 广东 广州510301)
根据2013年秋季(11月-12月)南沙群岛海域18个站位浮游纤毛虫和环境因子的调查资料,对浮游纤毛虫群落特征进行分析,并且探讨了影响浮游纤毛虫群落变动的主要环境因素。结果表明,共检出17种浮游纤毛虫,隶属于2目10属,砂壳目Tintinnida与寡毛目Oligotrichida。优势种包括三亚条纹虫Rhabdonellasanyahensis、锥形急游虫Strombidiumconicum、杆状真铃虫Eutintinnusstramentus、酒瓶类铃虫Codonellopsismorchella、球形急游虫Strombidiumglobosaneum。浮游纤毛虫平均丰度为(36.5±3.8) ind./L(10~80 ind./L),呈现斑块状分布,最大丰度出现在南沙群岛海域的中偏北部,其次在万安滩的东侧丰度值也较高,而在西北部、东北部和东南部的丰度值较低。浮游纤毛虫的水平分布可能与海洋环流有密切关系。此外,通过相关性分析和典范对应分析(canonical correspondence analysis,CCA)发现,南沙群岛海域浮游纤毛虫群落结构差异主要受水体营养盐水平与叶绿素a含量等环境因素的影响。
浮游纤毛虫;南沙群岛;群落结构;环境因子;多元分析
海洋浮游纤毛虫是海洋微型浮游动物的主要类群之一,它们以pico-级(0.2~2 μm)和nano-级(2~20 μm)浮游生物为食,然后被meso-级浮游动物和鱼类幼体摄食[1],因而是连接微食物环和经典食物链的重要中介,在海洋生态系统的物质传递和能量流动中发挥重要的作用[2-3]。
南沙群岛海域(4°-12°N,108°-118°E)位于南海南部,是典型的热带海洋性气候。由于受地形和季风驱动,中尺度涡频发,呈多涡环流结构[4-5],南沙群岛海域生境极其复杂,食物网组成亦比较复杂[6-7],因而具有独特的海洋科学研究价值。目前,有关南沙群岛海域浮游植物[6]、浮游动物[7]、水动力[8-9]和渔业资源[10]等方面已有调查和研究,但关于该海域浮游纤毛虫生态的研究还比较少。因此,研究南沙群岛海域浮游纤毛虫群落结构及空间分布并探讨浮游纤毛虫群落与环境因子间的关系,既为南海南部生态系统的研究提供基础资料,也为深入了解南沙群岛海域微食物环的能量流动提供帮助。
1 材料和方法
1.1 采样站位设置和样品采集
2013年11月8日至12月8日,搭乘“南锋号”考察船在北纬4°-12°,东经108°-118°范围内进行了调查,共设置18个站位,如图1所示。温度、盐度、水深由CTD直接测定,叶绿素a(Chla)水样首先通过200 μm筛绢过滤去除浮游动物,然后经GF/F滤膜过滤(用以测定Chla质量浓度),滤膜被吸干后保存在-20 ℃ 冰箱中。分别在每个站位5 m水层取5 L水样,加Lugol’s试剂固定(终体积分数2%),置于阴暗处保存,用以浮游纤毛虫样品的鉴定。同时,分别取上述各站位5 m水层水样200 mL,以测定分析其他水化学指标。
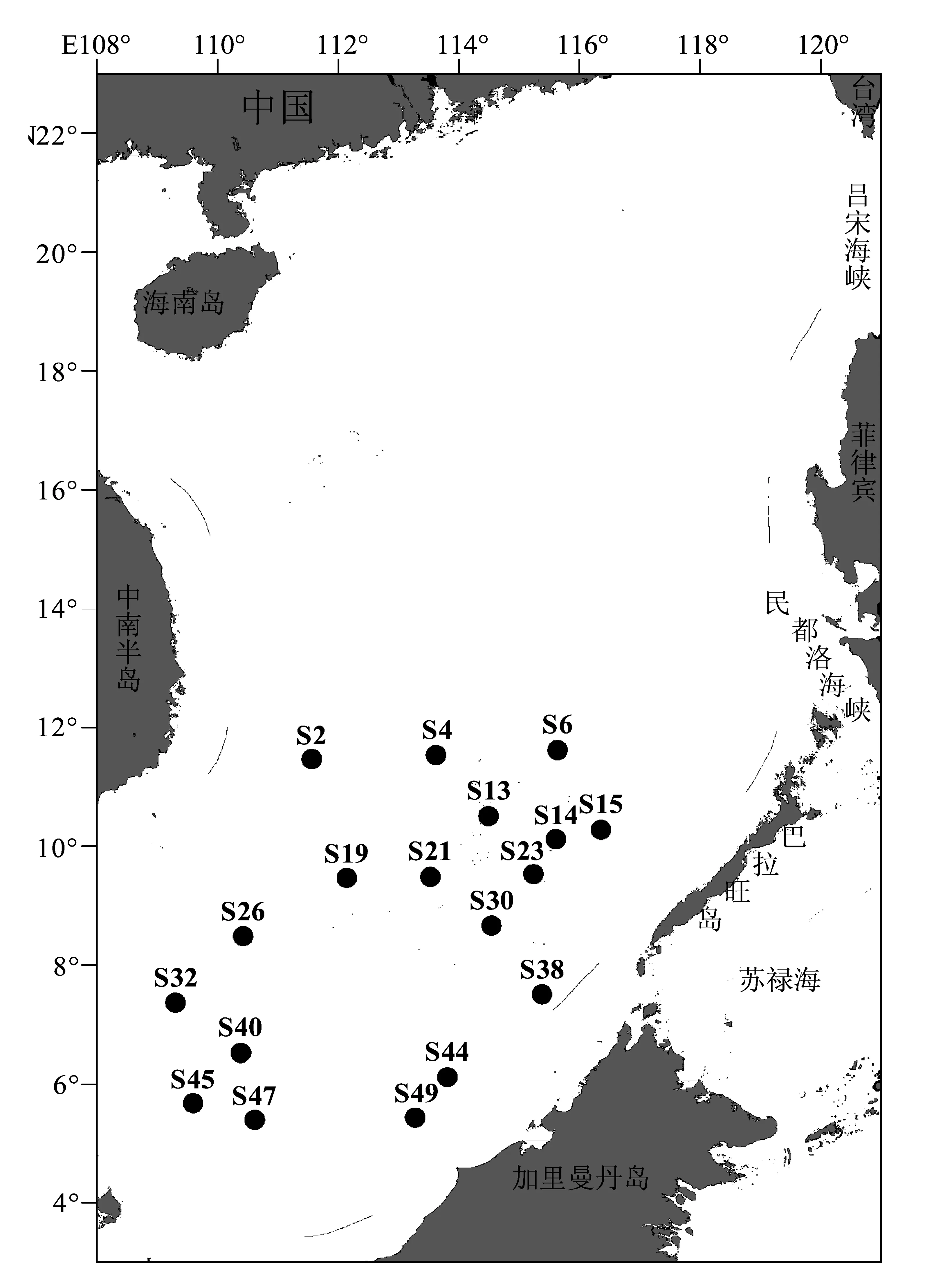
图1 南沙群岛海域采样站位图Fig.1 Sampling station in southern South China Sea
1.2 样品分析与鉴定
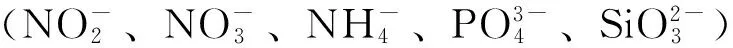
1.3 数据处理与分析
浮游纤毛虫群落指数:Shannon-Winner指数(H′),种类均匀度指数(J′),种类丰富度指数(d)(Jiang et al,2011)和优势度(Y),计算公式如下:
J′=H′/lnS
d=(S-1)/lnN
Y=(ni/N)×fi
其中Pi为样品中属于i种的丰度的比例;S为总种类数;N为总个体丰度;ni为第i种的丰度;fi为第i种的出现频率。优势种以Y≥0.02为划分标准[17]。
用大型多元统计软件PRIMER v5.0对浮游纤毛虫的群落结构进行聚类分析(CLUSTER)及多维定标分析(MDS);并使用ASOSIM模块检验各组间的相似程度;同时,采用BIOENV模块分析生物-环境相关性。生物因子相似性矩阵的建立采用Bray-Curtis相似性,浮游纤毛虫种类丰度数据转换采用4次方根转换[18];环境因素相似性矩阵的建立则采用欧氏距离(Euclidean)。
在CANOCO 4.5软件中应用典范对应分析方法(canonical correspondence analysis, CCA)来分析浮游纤毛虫与环境因子之间的关系时,首先筛选出现频率≥3个站位,且丰度比例≥1%的纤毛虫种类丰度数据,以保证进入CCA排序的种类能反映出整个浮游纤毛虫群落的特征。其次,浮游纤毛虫种类丰度数据、环境因子数据经lg(x+1)标准化转换。浮游纤毛虫与环境因素间的相关性分析使用SPSS v20.0软件。浮游纤毛虫丰度的平面分布分析采用surfer 12软件绘制。
2 结 果
2.1 环境因子


表1 2013秋末(11月8日-12月8日)南沙群岛海域各调查站位环境因素的变动Table 1 Variation of environmental variables in southern South China Sea in later autumn of 2013
2.2 浮游纤毛虫组成和丰度
南沙群岛海域18站共检出2目10属17种纤毛虫(表2),分别为寡毛目Oligotrichida和砂壳目Tintinnida。其中,砂壳目种类最多,共15种,占总种类的88.24%;寡毛目共2种,占总种类的11.76%。同样,砂壳目类纤毛虫丰度最大,占总丰度的80.65%,寡毛目类纤毛虫丰度占19.35%。
浮游纤毛虫优势种共5种(Y≥0.02,表2),依次是三亚条蚊虫Rhabdonellasanyahensis、锥形急游虫Strombidiumconicum、杆状真铃虫Eutintinnusstramentus、酒瓶类铃虫Codonellopsismorchella、球形急游虫Strombidiumglobosaneum。优势种的累积丰度占总丰度的50%,全部优势种的出现频率都在22%以上。

表2 2013秋末南沙群岛海域浮游纤毛虫名录及优势种Table 2 List of ciliate species and dominant species in southern South China Sea in later autumn of 2013
*优势种(Y≥0.02)
2.3 浮游纤毛虫的水平分布
南沙群岛海域的浮游纤毛虫丰度变化范围为10~80 ind./L,平均丰度为(36.5±3.8) ind./L,整体上呈现斑块状分布,最大丰度出现在中偏北部,其次万安滩东侧的丰度值也较高,而在西北部、东北部和东南部丰度较低(图2,图3a)。此外,不同站位之间纤毛虫种类数差异较大(1~5种之间)。在中部偏北S8、万安滩东侧的S10、S14站位种类最多,而种类低值区出现在S1、S3、S4和S15站(图3b)。

图2 2013年秋末南沙群岛海域浮游纤毛虫丰度水平分布图Fig.2 Horizontal distribution of planktonic ciliate abundance in southern South China Sea in later autumn of 2013

图3 2013年秋末南沙群岛海域浮游纤毛虫丰度(a)和种类数(b)Fig.3 Variation of ciliate abundance (a) and species number (b) in 18 sites of southern South China Sea in later autumn of 2013
2.4 浮游纤毛虫的群落结构
聚类分析的结果显示,南沙群岛海域18个站位的浮游纤毛虫样品在20%的相似水平上可聚为5组(图4a)。第1组包括S1,S6和S15站位的样品,该组样品均鉴定有球形急游虫(其他站位无此纤毛虫);第2组包括站位S4和S14站位的样品,该组样品均鉴定出根突拟铃虫Tintinnopsisradix(其他站位无此纤毛虫);第3组包括S9、S11、S13和S17站位的样品,该组样品均鉴定出变形条蚊虫Rhabdonellaamor(其他站位无此纤毛虫);第4组包括站位S16和S18站位的样品,其鉴定有简单条纹虫Protorhabdonellasimplex(其他站位无此纤毛虫);第5组包括站位S2、S3、S5、S7、S8、S10和S12站位的样品,三亚条纹虫是主要的贡献种。相似性分析(ANOSIM)(gloabR=0.763,p<0.001),表明聚类分析的各组之间有显著性差异。通过多维尺度分析(MDS)得出与聚类分析一致的结果,且分组情况同同上(图4b)。
2.5 浮游纤毛虫群落与环境因子的关系


图4 2013年秋末南沙群岛海域18个站位游纤毛虫样品的聚类分析树状图(a)和MDS分析图(b)Fig.4 Dendrogram of cluster analysis (a) and MDS plot (b) based on data of samples of 18 sites in southern South China Sea in later autumn of 2013
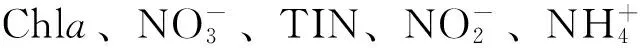
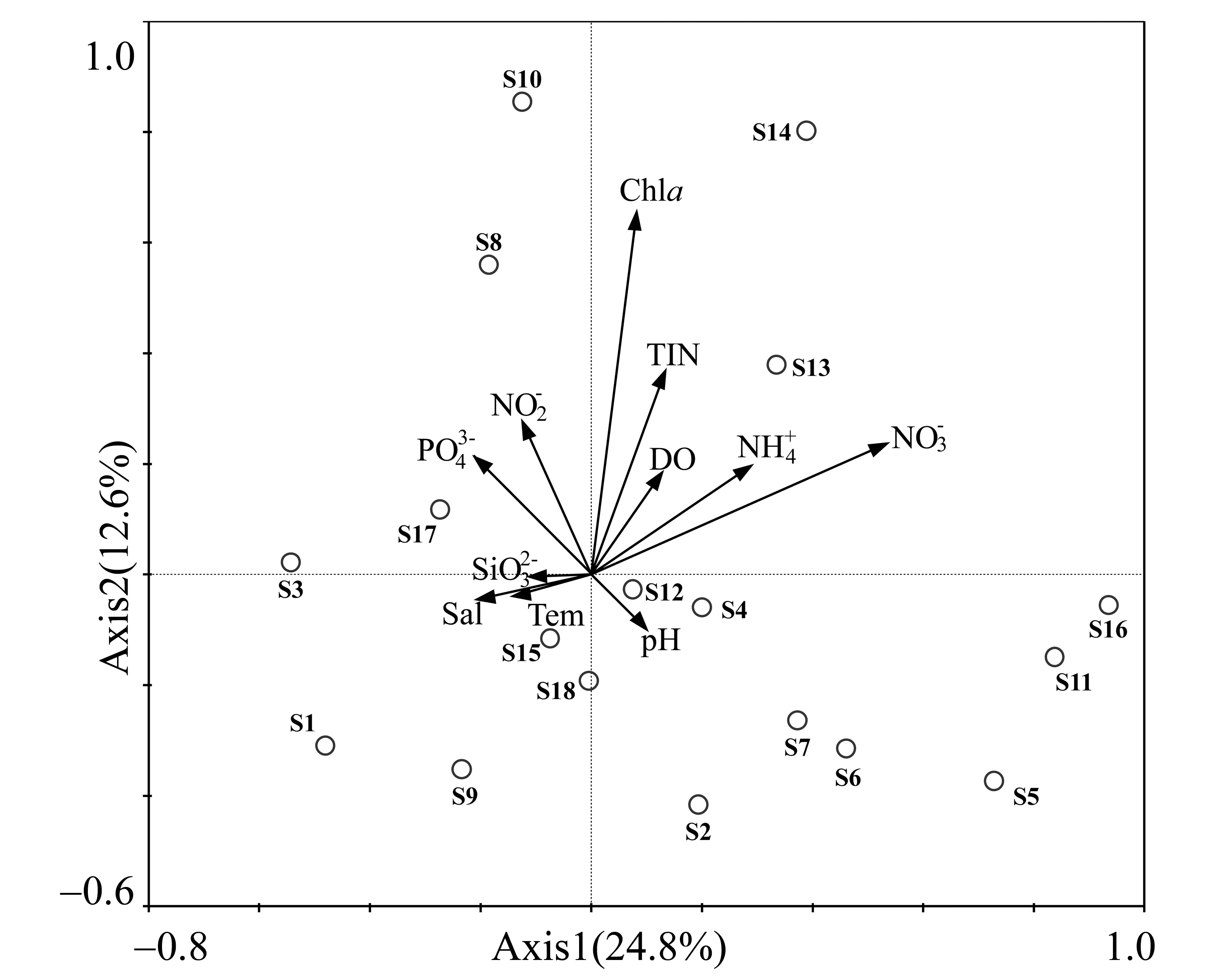
图5 2013年秋末南沙群岛海域浮游纤毛虫群落与环境因子关系的CCA二维排序图Fig.5 CCA two-dimensional plot showing the relationship between ciliate communities and environmental variables in southern South China Sea in later autumn of 2013
3 讨 论
3.1 浮游纤毛虫群落的水平分布
秋季南沙群岛海域浮游纤毛虫种类、丰度均是砂壳目纤毛虫占优势。一方面,由于南沙群岛海域受陆源输入和上升流的影响较小,营养盐水平和叶绿素a含量偏低,因此浮游纤毛虫丰度低于国内其它海域的结果[19-20]。
浮游纤毛虫群落结构受各种环境因子的影响,如盐度、温度、溶解氧、水动力特征、营养盐水平及食物来源等影响[21-24]。在东北季风期,浮游纤毛虫丰度高值区出现在南沙群岛海域中偏北部。该区域是一个复杂的多涡旋区域[25],位于北南沙反气旋环流与东南沙反气旋环流之间。气旋式冷涡可引起富营养盐底层水的涌升促发了浮游植物丰度的增高[26-27],进而浮游纤毛虫的丰度也相应较高。同样,王亮根等[28]发现浮游桡足类丰富区也位于两种南沙反气旋环流之间。另外,在西南部万安气旋涡附近出现丰度次高值。可见反式气旋环流之间及气旋环流与浮游纤毛虫群落分布有密切关系。
1) “*”表示显著正相关(p<0.05)
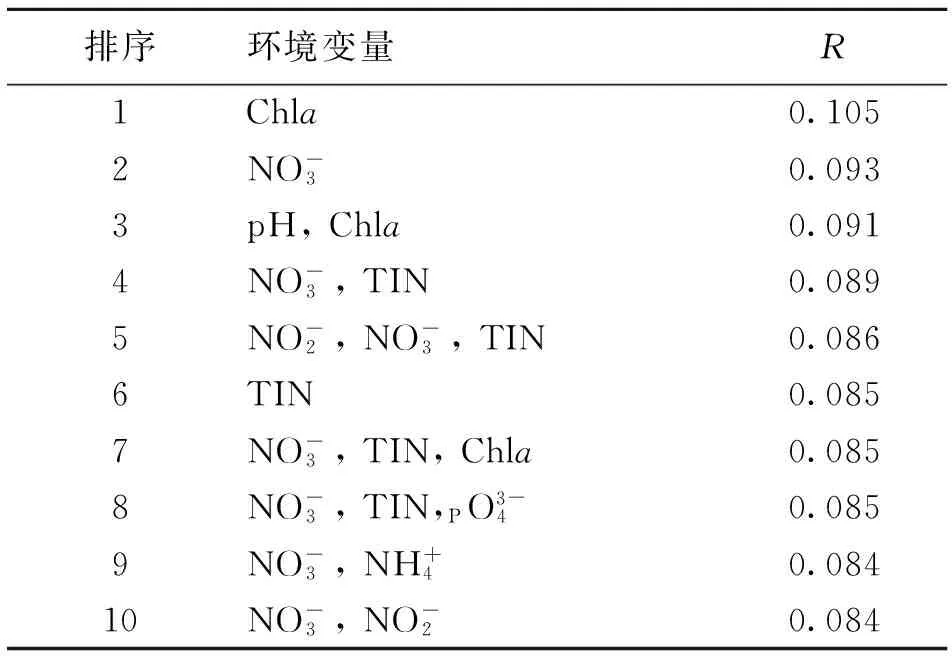
表4 2013年秋末南沙群岛海域18个采样站位浮游纤毛虫丰度与环境因子的BIOENV分析Table 4 Summary results from BIOENV for the best matches of environmental variables with spatial variations in ciliate at 18 sampling sites in southern South China Sea in later autumn of 2013
3.2 浮游纤毛虫群落与环境因子的关系

[1] PIERCE R W, TURNER J T. Global biogeography of marine tintinnids[J]. Marine Ecology-Progress Series, 1993, 94:11-26.
[2] AZAM F, FEBCHEL T, FIELD J G, et al. The ecological role of water-column microbes in the sea[J]. Marine Ecology-Progress Series, 1983, 10:257-263.
[3] PEREZ M T, DOLAN J R, VIDUSSI F, et al. Diel vertical distribution of planktonic ciliates within the surface layer of the NW Mediterranean (May 1995)[J]. Deep-Sea Research Part I-Oceanographic Research Papers, 2000, 47:479-503.
[4] 方文东, 郭忠信, 黄羽庭. 南海南部海区的环流观察研究[J]. 科学通报, 1997, 42(21):2264-2271.
[5] 管秉贤, 袁耀初. 中国近海及其附近海域若干涡旋研究综述I. 南海和台湾以东海域[J]. 海洋学报, 2006, 28(3):1-16.
[6] 宋星宇, 黄良民, 钱树本, 等.南沙群岛邻近海区春夏季浮游植物多样性研究[J]. 生物多样性, 2002, 10(3):258-268.
[7] 尹健强, 陈清潮, 张谷贤, 等.南沙群岛海区上层浮游动物种类组成与丰度的时空变化[J]. 科学通报, 2006,51(增刊II):129-138.
[8] 方文东. 南海南部海洋环流的结构与季节变化[D]. 青岛: 中国科学院海洋研究所, 2000.
[9] 刘辉, 江丽芳, 齐义泉, 等. 南沙群岛海域混合层深度季节变化特征[J]. 海洋科学进展, 2007, 25(3):268-279.
[10] 陈文河, 刘学东, 卢伙胜. 南沙群岛海域鱼类群落结构的季节性变化研究[J]. 热带海洋学报, 2010, 29(4):118-124.
[11] 宋微波.中国黄渤海的自由生纤毛虫[M]. 北京: 科学出版社, 2008.
[12] 刘瑞玉, 徐奎栋. 中国海洋生物名录-纤毛虫门[M]. 北京: 科学出版社, 2008.
[13] KAHL A. Urtitere oder protozoa[M]. Dahl ed: Die Tierwelt Deutschlands, 1935:181-805.
[14] CAREY P G. Marine interstitial ciliates[M]. London: Chapman & Hall, 1992, 14-324.
[15] KOFOID C A, CAMPBELL A S. A conspectus of the marine and freshwater Ciliata belonging to the suborder Tintinnoinea, with descriptions of new species principally from the Agassiz expedition to the eastern tropical Pacific 1904-1905[M]. Los Angeles: University of California Publications in Zoology, 1929:1-403.
[16] KOFOID C A, CAMPBELL A S. The Tintinnoinea of the eastern tropic Pacific[M]. Boston: Bulletin of the Museum of Comparative Zoology at Harvard College, 1939: 1-473.
[17] 徐兆礼, 陈亚性. 东黄海秋季浮游动物优势种聚集强度与給势渔场的关系[J]. 生态学杂志, 1989, 8(4):13-15.
[18] 廖秀丽, 陈丕茂, 马胜伟, 等.大亚湾杨梅坑海域投礁前后浮游植物群落结构及其与环境因子的关系[J]. 南方水产科学, 2013, 9(5):109-119.
[19] JIANG Y, XU H, HU X, et al. An approach to analyzing spatial patterns of planktonic ciliate communities for monitoring water quality in Jiaozhou Bay, northern China[J]. Marine Pollution Bulletin, 2011, 62:227-235.
[20] 刘华雪, 谭烨辉, 黄良民, 等. 夏季南海北部纤毛虫群落组成及其水平分布[J]. 生态学报, 2010, 30(9):2340-2346.
[21] SANDERS R W. Seasonal distributions of the photosynthesizing ciliatesLaboeastrobilaandMyrionectarubra(=Mesodiniumrubrum) in an estuary of the Gulf of Maine[J]. Aquatic Microbial Ecology, 1995, 9:237-242.
[22] MONTAGNES D J S, LESSARD E J. Population dynamics of the marine planktonic ciliateStrombidinopsismultiauris: its potential to control phytoplankton blooms[J]. Aquatic Microbial Ecology, 1999, 20:167-181.
[23] WANG Y, ZHANG W, LIN Y, et al. Spatial pattern of the planktonic ciliate community and its relationship with the environment in spring in the northern Beibu Gulf, South China Sea[J]. Oceanological and Hydrobiological Studies, 2013, 42(4):470-479.
[24] 于莹, 张武昌, 赵楠, 等. 胶州湾浮游纤毛虫丰度和生物量的周年变化[J]. 海洋与湖沼, 2011, 42(5):690-701.
[25] 黄企洲, 方文东, 陈荣裕. 南沙群岛海域西南季风变化与表层流结构和演变关系的探讨[J]. 热带海洋学报, 2001, 20(1):18-26.
[26] VAILLANCOURT R D, MARRA J, SCKI M P, et al. Impact of a cyclonic eddy on phytoplankton community structure and photosynthetic competency in the subtropical North photosynthetic competency in the subtropical North Pacific Ocean[J]. Deep-Sea Research Part I-Oceanographic Research Papers, 2003, 50:829-847.
[27] 戴明, 刘华雪, 廖秀丽, 等. 初秋南沙群岛海域网采浮游植物群落特征及其与环境因子的关系[J]. 应用生态学杂志, 2013, 24(12):3553-3560.
[28] 王亮, 杜飞雁, 李亚芳, 等.西南季风爆发前后南沙海域浮游桡足类群落特征比较研究[J]. 南方水产科学, 2015, 11(5):47-55.
[29] WANG Y, ZHANG W, LIN Y, et al. Spatial and seasonal variations of large tintinnid ciliates in Shenhu Bay of China[J]. Oceanological and Hydrobiological Studies, 2014, 43(3):292-302.
[30] UYE S, NAGANO N, TAMAKI H. Geographical and seasonal variations in abundance, biomass and estimated production rates of microzooplankton in the Inland Sea of Japan[J]. Journal of Oceanography, 1996, 52(6):689-703.
[31] REDFIELD A C. The biological control of chemical factors in the environment[J]. American Scientist, 1958, 46(3):205-221.
[32] 辛明, 王保栋, 孙霞, 等. 广西近海营养盐的时空分布特征[J]. 海洋科学, 2010, 34(9):5-9.
Planktonic ciliate community and its relationship with environmental factors in southern South China Sea during autumn
WU Fengxia1, HUANG Honghui1,2, TAN Yehui3, QI Zhanhui1
(1. Key Laboratory of Marine Fishery Ecology Environment of Guangdong Province∥Key Laboratory of South China Sea Fishery Resources Development and Utilization, Ministry of Agriculture∥South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China; 2.South China Sea Bio-Resource Exploitation and Utilization Collaborative Innovation Center, Guangzhou 510300, China; 3. South China Sea Institute of Oceanology, Guangzhou 510301, China)
Based on samples collected with CTD water sampler at a depth of 5 m from 18 sites in southern South China Sea during later autumn, 2013, species composition and abundance of planktonic ciliate and its relationship with the environment variables were investigated. A total 17 species belonging to 10 genera and two orders (Tintinnida and Oligotrichida) were identified. Among them, there were five dominant species (dominanceY≥0.02),includingRhabdonellasanyahensis,Strombidiumconicum,Eutintinnusstramentus,Codonellopsismorchella,Strombidiumglobosaneum. The mean abundance of ciliate was (36.5±3.8) ind./L (10~80 ind./L). Ciliates abundance distribution presents a patchy distribution, and the distribution of planktonic ciliates may be closely related to ocean circulation. Canonical correspondence analysis demonstrated that the nutrient level and Chlorophyllaconcentration were the most important factors affecting the spatial pattern of planktonic ciliate communities in southern South China Sea.
planktonic ciliate; southern South China Sea; community structure; environmental factors; multivariate analysis
10.13471/j.cnki.acta.snus.2016.05.016
2016-03-04
中央级公益性科研院所基本科研业务费专项资金资助项目(2014YD02);农业部财政重大资助项目(NFZX2013);广东省科技计划资助项目(2016A020222024)
吴风霞(1979年生),女;研究方向:渔业生态与环境;通讯作者:黄洪辉;E-mail:huanghh@scsfri.ac.cn
Q-9
A
0529-6579(2016)05-0089-07
猜你喜欢
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
流行色(2019年10期)2019-12-06
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
小学教学参考(语文)(2016年6期)2016-06-24
卷宗(2015年11期)2015-12-19
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
