
牛粪堆肥中厌氧氨氧化菌分子生物学检测
2016-05-05 11:59许修宏王婷婷孟庆欣孙雪微成利军张文浩任广明朱丽平王紫琪东北农业大学资源与环境学院哈尔滨150030
东北农业大学学报 2016年3期
关键词:堆肥
许修宏,王婷婷,孟庆欣,孙雪微,成利军,张文浩,孙 瑜,任广明,朱丽平,王紫琪,贺 宁(东北农业大学资源与环境学院,哈尔滨 150030)
牛粪堆肥中厌氧氨氧化菌分子生物学检测
许修宏,王婷婷,孟庆欣,孙雪微,成利军,张文浩,孙瑜,任广明,朱丽平,王紫琪,贺宁
(东北农业大学资源与环境学院,哈尔滨150030)
摘要:研究利用PCR-DGGE方法对堆肥中厌氧氨氧化菌16S rRNA基因特异扩增,检测堆肥厌氧氨氧化菌种属及多样性水平。结果表明,牛粪堆肥中厌氧氨氧化菌群组成与海洋环境不同。海洋环境中Candidatus Scalindua属厌氧氨氧化菌为优势菌群,而牛粪堆肥中厌氧氨氧化菌中优势菌属为Candidatus Brocadia、Candidatus Kuenenia和Candidatus Scalindua,多样性相对较高。可为堆肥发酵过程中厌氧氨氧化过程提供分子生物学依据。
关键词:厌氧氨氧化菌;堆肥;PCR-DGGE
许修宏,王婷婷,孟庆欣,等.牛粪堆肥中厌氧氨氧化菌分子生物学检测[J].东北农业大学学报,2016,47(3):52-58.
Xu Xiuhong,Wang Tingting,Meng Qingxin,et al.Molecular detection of anaerobic ammonium oxidation bacteria in cow manure composting[J].Journal of Northeast Agricultural University,2016,47(3):52-58.(in Chinese with English abstract)
厌氧氨氧化反应(Anammox)以亚硝酸为电子受体,铵氮为电子供体,缺氧条件下产生氮气,是氮素循环过程重要环节[1]。在反硝化工厂脱氮反应中发现厌氧氨氧化反应[2],在自然界中也发现厌氧氨氧化反应[3],并从黑海首次鉴定出厌氧氨氧化菌[4]。缺氧的海洋沉积物和水柱、淡水沉积物、陆地生态系统和一些特殊生态系统等自然环境中均有厌氧氨氧化菌存在[5]。Kuenen认为在海洋氮素循环中厌氧氨氧化菌贡献率达50%以上[6]。不同区域环境中,厌氧氨氧化菌对氮素损失贡献率在3%~ 41%之间,差异较大[7-9]。
厌氧氨氧化菌是厌氧菌,属浮霉菌门(Plancto-mycetes),目前分为5个属:Candidatus Brocadia[10];Candidatus Kuenenia[11];Candidatus Scalindua[12];Candidatus Anammoxoglobus[13]和Candidatus Jettenia[14],其中,Candidatus Scalindua主要分布在河口及海洋等水体环境中[15],其他四种主要分布在农田、湿地、水底沉积物和淡水河口等环境[16-19],Candidatus Brocadia和Candidatus Kuenenia是厌氧氨氧化菌中最常见的两个菌属。堆肥系统氮素循环中也存在厌氧氨氧化反应,但有关厌氧氨氧化菌分布及种群多样性研究鲜有报道。
堆肥是国内外对畜禽粪便无害化、资源化处理的主流技术。畜禽粪便经快速堆腐和无害化处理后生成优质有机肥料,施入农田系统,可实现废弃物农业循环再利用。氮素循环是堆肥过程关键步骤,氮素转化与堆肥产品腐熟度和氮素损失率密切相关,直接影响堆肥质量。因此,控制堆肥过程厌氧氨氧化反应造成的氮素挥发,提高堆肥质量是堆肥研究关注问题之一。传统观念认为堆肥中的氮素挥发途径主要是氨化作用和反硝化作用[20],厌氧氨氧化作用为堆肥氮素转化机理提供新理论依据。本研究利用PCR-DGGE技术对厌氧氨氧化菌16S rRNA基因扩增,证明堆肥中厌氧氨氧化菌存在,通过序列比对和进化树分析研究其多样性,为研究堆肥发酵过程中不同种属厌氧氨氧化菌消长规律,明确堆肥氮素循环过程厌氧氨氧化反应作用提供依据。
1 材料与方法
1.1堆肥材料制备
本研究所用牛粪取自哈尔滨市幸福乡奶牛养殖场,稻草秸秆来自东北农业大学香坊实验实习基地。堆肥混合物稻草秸秆截长度为5 cm,牛粪和稻草秸秆干重比例5ϑ1,含水率约65%,C/N为30ϑ1。将堆肥混合物在自制发酵罐(50 cm×50 cm×110 cm)中发酵处理,期间每隔30 min通风1次,每天测量3次温度,且按温度变化翻堆。
1.2样品采集
本研究发酵周期为30 d,分别在第0、1、3、7、13、23、29天取样。每次取样分别从上、中、下三层等量取样混合约500 g,保存温度-20℃,用于DNA提取。
1.3 DNA提取与纯化
分别取3~5 g堆肥样品用于DNA提取,提取方法参照Liu等方法[21]。提取DNA作琼脂糖凝胶电泳检测,试剂盒(Omega Bio-Tek,Inc.,Georgia,USA)纯化回收,具体操作参照说明书。
1.4 PCR与变性梯度凝胶电泳(DGGE)
以纯化DNA为模板,运用浮霉菌门通用引物Pla46f与细菌通用引物630r组合对细菌16S rRNA进行PCR扩增。以扩增得到PCR产物为模板,用来自厌氧氨氧化菌特异引物组合Amx368f(GC)/ Amx820r对Candidatus Brocadia和Candidatus Kuenenia菌属16S rRNA扩增,引物组合Pla46f/Amx368r (GC)对所有厌氧氨氧化菌属16S rRNA扩增。引物序列及来源见表1。
PCR反应体积为25 μL,10×PCR buffer(含Mg2+)2.5 μL,10 mmol·L-1dNTP 2.5 μL,Taq酶(5 U·μL-1)0.3 μL,10 μmol·L-1引物各0.2 μL,DNA模板0.5 μL(约10~100 ng),最后加无菌ddH2O 18.8 μL。第一轮PCR反应条件如下:94℃预变性5 min,94℃变性45 s,Tm值温度下退火1 min,72℃延伸40 s,30个循环,再以72℃延伸10 min。第一轮PCR产物稀释100倍作第二轮PCR模板。第二轮PCR反应条件如下:94℃预变性4 min,94℃变性45 s,Tm值温度下退火50 s,72℃延伸1 min,35个循环,72℃延伸7 min。以上3对引物PCR扩增产物用1%琼脂糖凝胶电泳检测。
DGGE分析采用DCodeTM系统(Bio-Rad公司),并用8%聚丙烯酰胺凝胶(丙烯酰胺ϑ双炳烯酰胺=37.5ϑ1 m·V-1)电泳,PCR产物加样量25 μL,其中引物组合Amx368f(GC)/Amx820r利用凝胶变性剂浓度范围为25%~55%,引物组合Pla46f/ Amx368r(GC)利用凝胶变性剂浓度范围为40%~ 60%。电泳运行条件:1×TAE电泳缓冲液,60℃恒温,恒压100 V,电泳12 h。电泳结束后,DGGE凝胶用硝酸银染色,回收目的片段条带,溶于去离子水中。12 h后以特异引物Amx368f/Amx820r和Pla46f/ Amx368r分别对回收DNA再次PCR扩增,将扩增后产物测序,所获得序列均提交GenBank数据库,基因登录号为KT003616-KT003626。
1.5 DGGE图谱分析
根据DGGE图谱及Quantity One 4.6.2软件对图谱条带分析,由公式(1)计算每个堆肥样品中微生物多样性指数(Shannon-Wiener,H),预测堆肥样品中厌氧氨氧化菌分布。

表1 试验所用PCR引物Table 1 PCR primers used in this study

式中,Pi=Ni/N,Ni-样品上各条带吸收峰面积,N-样品上所有条带吸收峰总面积。
运用Quantity one 4.6.2软件UPGAMA对DGGE图谱聚类分析,并应用SPSS 21.0软件对堆肥样品DGGE图谱进行主成分分析(PCA)。
1.6系统发育进化分析
利用Blast程序对分离到的DNA序列与GenBank数据库中已知序列比对,运用MEGA(version6.06)[24]中NJ(Neighbor-Joining)法构建系统进化树,采用Maximum Likelihood计算,Bootstrap分析值为1 000个样本。
2 结果与分析
2.1厌氧氨氧化菌DGGE图谱分析
以来自不同堆肥样品DNA为模板进行第一轮PCR,再以第一轮PCR产物为第二轮PCR模板扩增,比较PCR产物DGGE图谱发现,虽然不同样品电泳图在同一位置上有相同条带,但其条带粗细、信号强度不同,说明不同种属在堆肥发酵不同阶段数量存在一定差异(见图1)。测序结果表明,图1A检测到的DGGE条带中,条带A和K存在于整个堆肥过程,是堆肥优势类群。堆肥发酵不同阶段样品含有许多特异条带,如F和G分别是Day7和Day13特异条带,H主要存在于堆肥前期(Day0和Day1),B主要出现在高温期(Day3和Day7),C主要出现在堆肥后期(Day13、Day23和Day29)。图1B中,条带C和I存在于每个堆肥样品中,是优势类群。B和J主要出现在堆肥腐熟阶段(Day23和Day29),A、D、K和L主要出现在堆肥前期(Day0和Day1),E和M主要出现在高温期(Day3和Day7),F、B和J主要出现在堆肥后期(Day13、Day23和Day29)。可见,A、D、H、K和L在堆肥前期起作用,B、E和M在高温期起作用,F、B和J在堆肥后期起作用。
2.2 DGGE图谱多样性分析
利用以上Quantity One软件分析DGGE图谱多样性,得出不同引物组合检测堆肥不同阶段样品菌群多样性指数H。结果表明,引物组合Amx368f/ Amx820r的PCR检测中,堆肥高温期(Day7)多样性指数最高,腐熟期(Day29)多样性指数最低。引物组合Pla46f/Amx368r的PCR检测中,降温期(Day13)厌氧氨氧化菌多样性指数最高,腐熟期(Day29)多样性指数最低(见图2)。
2.3系统发育进化分析
将本研究获得DNA序列与GenBank数据库相似序列比对,MEGA软件构建树状图。结果表明,引物组合Amx368f/Amx820r检测到的厌氧氨氧化菌主要为Candidatus Brocadia、Candidatus Scalindua和Candidatus Kuenenia三个属,其中多数厌氧氨氧化菌相似于Candidatus Brocadia属。F、E、C、A 和D类似于Candidatus Brocadia属厌氧氨氧化菌,其中A和C与已知Candidatus Brocadia属厌氧氨氧化菌相似性较高,分别为95%(AF375994)和98% (KF810110);而E和D与已知Candidatus Brocadiafulgida相似性仅为90%和91%。H和G与Candidatus Scalindua属厌氧氨氧化菌相近(主要来自河口沉积物)[25],H与Candidatus Scalindua wagneri相似性为95%,G与Candidatus Scalindua marina相似性达96%。K与Candidatus Kuenenia属的Candidatus Kuenenia stuttgartiensis相似性达95%。J、I和B类似于不可培养的Planctomycetes细菌(见图3)。
引物组合Pla46f/Amx368r检测到的G、J、A、 M、D、H和B均相似于Candidatus Brocadia属厌氧氨氧化菌,但与已知菌属相似性较低,只有M与已知Candidatus Brocadia caroliniensis相似性最高,为94%。C、K和F与Candidatus Scalindua属厌氧氨氧化菌相似,其中C与Candidatus Scalindua wagneri相似性达95%。I相似于Candidatus Kuenenia属厌氧氨氧化菌。此系统发育进化树中同样存在不可培养的Planctomycetales细菌,即E和L(见图4)。

图1 堆肥中厌氧氨氧化菌16S rRNA基因DGGE图谱Fig.1 16S rRNA gene DGGE profile of anammox bacteria
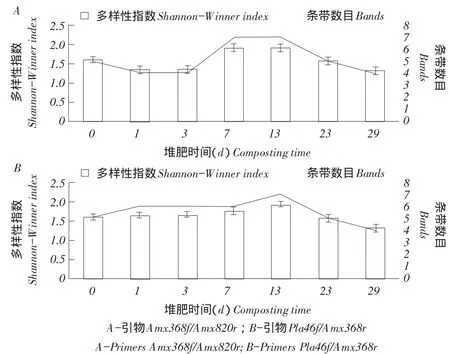
图2 厌氧氨氧化菌16S rRNA基因多样性指数Fig.2 16S rRNA gene Shannon-Winner index of anammox bacteria

图3 引物Amx368f/Amx820r扩增所得到序列系统发育进化树Fig.3 Phylogenitic tree showing the affiliations phylotypes using primer Amx368f/Amx820r
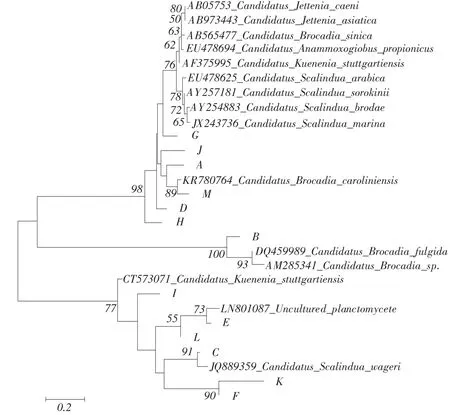
图4 引物Pla46f/Amx368r扩增所得序列系统发生进化树Fig.4 Phylogenitic tree showing the affiliations phylotypes using primer Pla46f/Amx368r
3 讨 论
在水体生态系统中,厌氧氨氧化过程所需亚硝态氮由硝化作用提供[26]。堆肥系统中,尽管高温期铵态氮大量挥发,但硝化作用和反硝化作用仍可提供充足亚硝态氮,亚硝态氮和铵态氮同时存在厌氧环境条件[27],厌氧氨氧化反应即可发生。堆肥后期,虽然硝态氮含量增多,但铵态氮越来越少[19],反硝化细菌数量增多[28],可能与厌氧氨氧化菌协同抑制[29],故厌氧氨氧化菌数量不断减少,多样性降低。本研究结果显示,牛粪堆肥过程中各时期均存在厌氧氨氧化菌,堆肥过程中厌氧氨氧化菌多样性整体呈先升后降趋势,多样性较高的样品包括Day7(高温期)和Day13(降温期),样品Day7多样性高说明厌氧氨氧化菌存在于堆肥高温环境中,主要菌属为Candidatus Brocadia。Byrne指出厌氧氨氧化菌可存在于85℃高温[30],主要菌属是Candidatus Brocadia和Candidatus Kuenenia,与本研究结果吻合。Candidatus Scalindua属主要出现在堆肥初期及后期,可能与其适合较低温度环境,如稻田有关[17]。而在降温期(Day13)厌氧氨氧化菌多样性同样较高,可能与堆体温度下降,适合多种细菌生长有关(特别是硝化细菌)。
分析细菌16S rRNA系统发育进化树发现,引物Amx368f/Amx820r经PCR得到的DNA序列与引物Pla46f/Amx368r经PCR得到序列存在一定差异,前者获得序列与已知厌氧氨氧化菌序列相似性更高。与已知菌相似性达90%~99%,而后者获得序列与本研究测得序列中仅M和C与已知厌氧氨氧化菌相似性高于90%,表明利用引物Amx368f/ Amx820r PCR更有利于扩增出与已知序列高同源性的厌氧氨氧化菌,与Sànchez-Melsió等研究结果一致[31]。
4 结 论
a.通过PCR-DGGE技术验证厌氧氨氧化菌存在于牛粪与稻草秸秆混合堆肥中,即存在于整个堆肥过程。
b.堆肥发酵过程中高温期(Day7)和降温期(Day13)厌氧氨氧化菌多样性较高。
c.堆肥发酵不同时期厌氧氨氧化菌群发生更替。
d.本研究检测到的厌氧氨氧化菌主要属于Candidatus Brocadia、Candidatus Scalindua和Candidatus Kuenenia三个属,其中Candidatus Brocadia属厌氧氨氧化菌占大多数。
[参考文献]
[1]Van d G A A,Mulder A,De Bruijn P,et al.Anaerobic oxidation of ammonium is a biologically mediated process[J].Appl Environ Microbiol,1995,61(4):1246-1251.
[2]Mulder A,van der Graaf A A,Robertson L A,et al.Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor[J].FEMS Microbiol Ecol,1995,16:177-184.
[3]Thamdrup B,Dalsgaard T.Production of N(2)through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments[J].Appl Environ Microbiol,2002,68(3):1312-1318.
[4] Kuypers M M M,Sliekers A O,Lavik G,et al.Anaerobic ammonium oxidation by Anammox bacteria in the Black Sea[J].Nature,2003,422(6932):608-611.
[5]Hu B L,Shen L D,Xu X Y,et al.Anaerobic ammonium oxidation(Anammox)in different natural ecosystems[J].Biochemical Society Transactions,2011,39(23):1811-1816.
[6]Kuenen J G.Anammox bacteria:from discovery to application[J].Nature Reviews Microbiology,2008,6(4):320-326.
[7]Naeher S,Huguet A,Roose- Amsaleg C L,et al.Molecular and geochemical constraints on anaerobic ammonium oxidation(Anammox)in a riparian zone of the Seine Estuary(France)[J].Biogeochemistry,2015,123(1/2):237-250.
[8]王衫允,祝贵兵,曲冬梅,等.白洋淀富营养化湖泊湿地厌氧氨氧化菌的分布及对氮循环的影响[J].生态学报,2012,21(21):6591-6598.
[9]Nie S,Li H,Yang X,et al.Nitrogen loss by anaerobic oxidation of ammonium in rice rhizosphere[J].IsmeJournal,2015,9(9):2059-2067.
[10]Strous M,Fuerst J A,Kramer E H,et al.Missing lithotroph identified as new planctomycete[J].Nature,1999,400(6743):446-449.
[11]Schmid M,Twachtmann U K M,Strous M,et al.Molecular evidence for genus level diversity of bacteria capable of catalyzing anaerobic ammonium oxidation[J].Systematic & Applied Microbiology,2000,23(1):309-320.
[12]Schmid M,Walsh K,Webb R I,et al.Candidatus 'Scalinduabrodaea',spec.nov.,Candidatus 'Scalindua wagneri',spec.nov.,two new species of anaerobic ammonium oxidizing bacteria[J].Systematic & Applied Microbiology,2003,26(4):529-538.
[13]Kartal B,Rattray J,Niftrik L A V,et al.Candidatus "Anammoxoglobus propionicus" a new propionate oxidizing species of anaerobic ammonium oxidizing bacteria[J].Systematic & Applied Microbiology,2007,30(1):39-49.
[14]Quan Z X,Rhee S K,Zuo J E,et al.Diversity of ammoniumoxidizing bacteria in a granular sludge anaerobic ammoniumoxidizing(Anammox)reactor[J].Environmental Microbiology,2008,10(11):3130-3139.
[15]Schmid M C,Nils R P,Jack V D V,et al.Anaerobic ammoniumoxidizing bacteria in marine environments:widespread occurrence but low diversity[J].Environmental Microbiology,2007,9(6):1476-1484.
[16]Humbert S,Tarnawski S,Fromin N,et al.Molecular detection of Anammox bacteria in terrestrial ecosystems:distribution and diversity[J].Isme Journal,2010(4):450-454.
[17]Wang J,Gu J D.Dominance of candidatus scalindua species in Anammox community revealed in soils with different duration of rice paddy cultivation in northeast china[J].Applied Microbiology & Biotechnology,2013,97(4):1785-1798.
[18]Zhao Y,Xia Y,Kana T M,et al.Seasonal variation and controlling factors of anaerobic ammonium oxidation in freshwater river sediments in the Taihu Lake region of China[J].Chemosphere,2013,93(9):2124-2131.
[19]Shen L D,Liu S,Lou L P,et al.Broad distribution of diverse anaerobic ammonium-oxidizing bacteria in Chinese agricultural soils[J].Appl Environ Microbiol,2013,79:6167-6172.
[20]张雪辰,王旭东.畜禽粪便快速发酵过程中的氮素转化及损失[J].农业环境科学学报,2014,3(3):458-464.
[21]Liu J,Xu X H,Li H T,et al.Effect of microbiological inocula on chemical and physical properties and microbial community of cow manure compost[J].Biomass and Bioenergy,2011,35:3433-3439.
[22]Neef A,Amann R,Schlesner H,Schleifer,K H.Monitoring a widespread bacterial group:in situ detection of planctomycetes with 16S rRNA- targeted probes[J].Microbiology,1998,144:3257-3266.
[23]Juretschko S,Timmermann G,Schmid M,et al.Combined molecular and conventional analyses of nitrifying bacterium diversity in activated sludge:Nitrosococcus mobilis and nitrospira-like bacteriaas dominant populations[J].Applied & Environmental Microbiology,1998,64(8):3042-3051.
[24]Tamura K,Stecher G,Peterson D,et al.MEGA6:Molecular evolutionary genetics analysis version 6.0[J].Molecular Biology & Evolution,2013,30(4):2725-2729.
[25]Hou L,Zheng Y,Liu M,et al.Anaerobic ammonium oxidation and its contribution to nitrogen removal in China's coastal wetlands[J].Scientific Reports,2015(5):15621.
[26]Kuypers M M,Lavik G,Woebken D,et al.Massive nitrogen loss from the Benguela upwelling system through anaerobic ammonium oxidation[J].Proceedings of the National Academy of Sciences of the United States of America,2005,102(18):6478-6483.
[27]Francis C A,Beman J M,Kuypers M M.New processes and players in the nitrogen cycle:the microbial ecology of anaerobic and archaeal ammonia oxidation[J].Isme Journal,2007,1(1):19-27.
[28]马丽红,黄懿梅,李学章,等.牛粪堆肥化中氮素形态与微生物生理群的动态变化和耦合关系[J].农业环境科学学报,2009,12(12):2674-2679.
[29]贾方旭,彭永臻,杨庆.厌氧氨氧化菌与其他细菌之间的协同竞争关系[J].环境科学学报,2014,34(6):1351-1361.
[30]Byrne N,Strous M,Crépeau V,et al.Presence and activity of anaerobic ammonium-oxidizing bacteria at deep-sea hydrothermal vents[J].Isme Journal,2009,3(1):117-123.
[31]Sànchez-Melsió A,Cáliz J,Balaguer M D,et al.Development of batch-culture enrichment coupled to molecular detection for screening of natural and man-made environments in search of anammox bacteria for N- removal bioreactors systems[J].Chemosphere,2009,75(2):169-179.
Molecular detection of anaerobic ammonium oxidation bacteria in cow manure composting
XU Xiuhong,WANG Tingting,MENG Qingxin,SUN Xuewei,CHENG Lijun,ZHANG Wenhao,SUN Yu,REN Guangming,ZHU Liping,WANG Ziqi,HE Ning(School of Resources and Environmental Sciences,Northeast Agricultural University,Harbin 150030,China)
Abstract:In this study,the species and diversity of anammox bacteria 16S rRNA gene in composting process were detected by PCR- DGGE technology.The results showed that anammox bacterial genera in cow manure composting were different from the marine environment.Candidatus Scalindua was dominat anammox bacteria in marine environment,while Candidatus Brocadia,Candidatus Kuenenia and Candidatus Scalindua were dominant anammox bacteria in cow manure compost,and anammox bacteria diversity was relatively high.This study provided the molecular evidence for the existence of anammox during the composting process.
Key words:anammox bacteria; compost; PCR-DGGE
作者简介:许修宏(1968-),男,教授,博士,博士生导师,研究方向为应用微生物学。E-mail:xuxiuhong@neau.edu.cn
基金项目:国家自然科学基金项目(31272484,31372351)
收稿日期:2015-12-06
中图分类号:S141.4
文献标志码:A
文章编号:1005-9369(2016)03-0052-07
猜你喜欢
绿色科技(2016年22期)2017-03-15
吉林农业(2016年12期)2017-01-06
江苏农业科学(2016年2期)2016-04-11
江苏农业科学(2015年11期)2016-01-27
