
饲料中铬源及铬添加水平对凡纳滨对虾幼虾生长性能、血清生化指标及非特异性免疫酶活性的影响
2016-04-19 03:42蔡海瑞谭北平杨奇慧董晓慧迟淑艳刘泓宇广东海洋大学水产学院水产动物营养与饲料实验室湛江524088
动物营养学报 2016年3期
关键词:生长性能
蔡海瑞 谭北平 杨奇慧 董晓慧 迟淑艳 刘泓宇 章 双(广东海洋大学水产学院,水产动物营养与饲料实验室,湛江524088)
饲料中铬源及铬添加水平对凡纳滨对虾幼虾生长性能、血清生化指标及非特异性免疫酶活性的影响
蔡海瑞谭北平∗杨奇慧∗董晓慧迟淑艳刘泓宇章双
(广东海洋大学水产学院,水产动物营养与饲料实验室,湛江524088)
摘要:本试验旨在研究饲料中铬源及铬添加水平对凡纳滨对虾幼虾生长性能、血清生化指标及非特异性免疫酶活性的影响,探讨3种铬源在饲料中的最适添加水平。采用双因素试验设计,铬源分别为氯化铬(CrCl3)、吡啶甲酸铬(Cr-Pic)和蛋氨酸铬(Cr-Met),铬添加水平分别为0、0.3、0.6、0.9、1.2和2.0 mg/ kg,配制成16种试验饲料,投喂凡纳滨对虾幼虾8周。挑选初始体重为(0.897±0.001)g的凡纳滨对虾幼虾1 920尾,随机分为16组,每组3个重复,每个重复40尾。结果表明:铬源、铬添加水平及两者的交互作用对凡纳滨对虾的终末体重、增重率、饲料系数和蛋白质效率有显著影响(P<0.05)。铬添加水平为0.3~2.0 mg/ kg的各组幼虾的终末体重、增重率显著高于未添加铬组(P<0.05),以Cr-Met形式添加0.9 mg/ kg铬时上述指标达到最大值。CrCl3和Cr-Met组在铬添加水平为0.9 mg/ kg时饲料系数最低,但与铬添加水平为1.2 mg/ kg时差异不显著(P>0.05);Cr-Pic组在铬添加水平为1.2 mg/ kg时饲料系数最低。3种铬源下,蛋白质效率均在铬添加水平为0.9 mg/ kg时达最大值,但与铬添加水平为1.2 mg/ kg时差异不显著(P>0.05)。铬源、铬添加水平及两者的交互作用对全虾粗脂肪和粗灰分含量有显著影响(P<0.05),但对全虾粗蛋白质和水分含量无显著影响(P>0.05)。铬源、铬添加水平及两者的交互作用对血清总蛋白、葡萄糖、胆固醇和甘油三酯含量均有显著影响(P<0.05),以Cr-Met形式添加0.9 mg/ kg铬时呈现出最高的血清总蛋白含量和最低的血清葡萄糖含量。铬源、铬添加水平及两者的交互作用对血清碱性磷酸酶、酸性磷酸酶、酚氧化物酶及总超氧化物歧化酶活性有显著影响(P<0.05),以Cr-Met形式添加0.9 mg/ kg铬时血清酚氧化物酶和总超氧化物歧化酶活性最高。以增重率为评价指标,以CrCl3、Cr-Pic和Cr-Met为铬源时,经折线模型得出饲料中铬的适宜添加水平分别为1.33、1.27、1.04 mg/ kg。通过比较可知,Cr-Met的相对生物利用率最高,Cr-Pic次之,CrCl3最低。
关键词:凡纳滨对虾;铬;生长性能;血清生化指标;非特异性免疫酶活性
铬(Cr)是动物的必需微量元素之一,是葡萄糖耐量因子(glucose tolerance factor,GTF)的重要活性组成成分,铬通过葡萄糖耐量因子协同胰岛素的作用,影响碳水化合物、脂类、蛋白质和核酸代谢,进而影响动物的生长、免疫、繁殖和胴体品质,降低应激,改善机体免疫机能,提高生产性能和繁殖力[1]。
铬作为营养素最早多在畜禽方面进行研究,近年来,铬在水产养殖上的研究也取得了一些成果,特别是在鱼类上的研究比较多。研究表明,饲料中添加铬可促进鲤鱼(Cyprinus carpio Linnaeus)[2]、草鱼(Ctenopharyngodon idellus)[3-4]、河蟹(Eriocheir sinensis)[5]、奥尼罗非鱼(Oreochoromis niloticus×O. aureus)[6]、吉富罗非鱼(Oreochromis niloticus)[7]和团头鲂(Megalobrama amblycephala)[8]等水产动物的生长和饲料利用率,但对不同品种间的研究结果差异较大。有关凡纳滨对虾(Litopenaeus vannamei)对饲料中铬需求量的研究主要集中在以吡啶甲酸铬(Cr-Pic)为铬源上。杨奇慧等[9]报道,以Cr-Pic为铬源时,凡纳滨对虾对饲料中铬的需求量为1.2~1.6 mg/ kg。
铬的吸收与其价态及化学形式有很大关系,Mertz等[10]认为有机铬较无机铬容易吸收,其吸收率为25%~30%。无机铬中的三氯化铬(CrCl3)性质很稳定,不易水解,是合成其他铬盐的重要原料,在无机合成和有机合成中有重要作用,是合成饲料添加剂的主要成分。Cr-Pic和蛋氨酸铬(Cr-Met)分别是吡啶甲酸和蛋氨酸与3价铬离子(Cr3+)的螯合物,可缓解矿物元素之间的拮抗竞争作用,有利于铬的吸收。本研究拟以我国养殖量最大的经济甲壳动物凡纳滨对虾为研究对象,比较3种不同形式的铬——无机铬、有机酸铬、氨基酸铬及其添加水平对凡纳滨对虾生长性能、血清生化指标和免疫的影响,为铬在对虾配合饲料中的应用提供理论依据。
1 材料与方法
1.1试验饲料
以酪蛋白、红鱼粉为蛋白质源,鱼油、玉米油、卵磷脂为脂肪源,配制基础饲料(表1)。在基础饲料的基础上,采用双因素试验设计,以CrCl3(由佛山市海纳化工有限公司提供)、Cr-Pic(由绵阳市新一美化工有限公司提供,Cr3+含量为0.1%)、Cr-Met(由广州天科科技有限公司提供,Cr3+含量为0.1%)为铬源,铬添加水平分别为0、0.3、0.6、0.9、1.2和2.0 mg/ kg,共配制成16种试验饲料。各种饲料原料粉碎后过80目筛,按表1的配方准确称量各饲料原料,3种铬源先分别用沸石粉作为稀释剂,稀释成Cr3+含量为0.1%,然后按各试验组的添加量进行称取,再和饲料配方中其他微量成分混合均匀(微量成分采取逐级扩大法混合均匀),各饲料原料成分用V型混合机混合均匀后,添加30%左右的水用搅拌机再次混匀后,压制成粒径分别为1.0和1.5 mm的2种颗粒饲料,经60℃熟化30 min后风干,用封口袋密封,置于-20℃冰箱中保存备用。

表1 基础饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of the basal diet(DM basis) %

表2 试验饲料中铬的测定量Table 2 The analyzed values of chromium in experimental diets mg/ kg
1.2试验动物及饲养管理
试验虾苗购自湛江市中联养殖有限公司,购回后暂养一段时间,期间投喂基础饲料,每天3次,饱食投喂。试验前,挑选大小均匀、体格健康、初重为(0.897±0.001)g的虾苗1 920尾,随机分为16组,每组设3个重复,每个重复40尾虾,以重复为单位饲养于容积为0.3 m3的玻璃纤维钢桶中。每天按体重的8%~10%投喂,分别于07:00、11:00、17:00、21:00各投喂1次,每餐以接近饱食(30 min内摄食完毕)为限。试验用水为经过沉淀、过滤的海水,试验第1~4周每2 d换水1次,第5~8周每天换水1次,换水量为总水量的30%~50%,每天观察对虾摄食、蜕壳、生长情况并记录投喂量。试验在广东海洋大学东海岛海洋生物研究基地室内养殖系统内进行。养殖过程参照杨奇慧等[9]的方法进行。试验期间连续充氧,水温为28~31℃,溶解氧浓度>7.0 mg/ L,盐度为26~28,pH 为7.8~8.2,氨氮浓度<0.03 mg/ L。试验期为8周,结束前24 h停止投喂,称重、计数。
1.3样品采集
试验结束时,对每桶虾进行称重并计数,用于计算成活率、增重率、特定生长率等生长指标。每桶随机取10尾虾,用1 mL无菌注射器从围心腔取血,将血液置于1.5 mL离心管中,于4℃冰箱中静置过夜后,在4℃、8 000 r/ min下离心10 min,取上清分装后置于-80℃冰箱中保存,用于分析血清生化指标;每桶另随机选5尾虾保存于-20℃冰箱内,用于体成分分析。
1.4指标测定
1.4.1体成分分析
参照AOAC(1995)[11]方法,分别测定饲料及全虾样品中的水分、粗蛋白质、粗脂肪和粗灰分的含量。水分含量的测定通过恒温烘箱在105℃下烘干至恒重测定;粗蛋白质含量测定采用凯氏定氮法(KjeltecTM8400,Sweden);粗脂肪含量的测定采用索氏抽提法(乙醚作为提取溶剂);粗灰分含量的测定采用550℃马弗炉灰化法。
1.4.2血清生化指标的测定
血清中总蛋白、葡萄糖、甘油三酯、胆固醇含量采用日立Hitachi 7020型全自动生化分析仪测定,所用试剂购自威特曼(南京)生物科技有限公司。
1.4.3血清非特异性免疫酶活性的测定
血清中总超氧化物歧化酶(total-surperoxide dismutase,T-SOD)、酸性磷酸酶(acid phosphatase,ACP)、碱性磷酸酶(alkaline phosphatase,AKP)活性使用南京建成生物工程研究所生产的试剂盒测定,测定方法参照试剂盒说明书进行;血清中酚氧化物酶(phenoloxidase,PO)活性的测定参照王建国等[12]和Huang等[13]的方法。
1.5生长性能指标计算公式

1.6数据处理与分析
结果用平均值±标准差表示。采用SPSS 17.0统计软件对数据进行双因素方差分析(two-way ANOVA),若存在显著性差异,再采用Duncan氏法进行多重比较,检验组间差异的显著性,P<0.05表示差异显著。饲料中铬的最适添加水平通过折线模型进行回归分析,用拐点以下的直线斜率比值比较凡纳滨对虾对CrCl3、Cr-Pic和Cr-Met的相对生物利用率。
2 结 果
2.1饲料中铬源及铬添加水平对凡纳滨对虾幼虾生长性能的影响
由表3可知,铬源及铬添加水平对凡纳滨对虾的终末体重、增重率、饲料系数和蛋白质效率均有显著影响(P<0.05),且铬源与铬添加水平之间存在交互作用(P<0.05)。铬源对凡纳滨对虾的成活率有显著影响(P<0.05),但铬添加水平对其无显著影响(P>0.05),且铬源与铬添加水平间无交互作用(P>0.05)。
终末体重:以CrCl3和Cr-Met为铬源时,0.9 和1.2 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic为铬源时,1.2 mg/ kg组显著高于其余各组(P<0.05)。添加0.3、0.6、0.9 mg/ kg铬时,Cr-Met组显著高于CrCl3和Cr-Pic组(P<0.05);添加1.2、2.0 mg/ kg铬时,CrCl3组显著低于Cr-Pic和Cr-Met组(P<0.05)。
增重率:以CrCl3和Cr-Met为铬源时,0.9和1.2 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic为铬源时,1.2 mg/ kg组与2.0 mg/ kg组无显著差异(P>0.05),而显著高于其余各组(P<0.05)。添加0. 6、0. 9 mg/ kg铬时,Cr-Met组显著高于CrCl3和Cr-Pic组(P<0.05);添加1.2、2.0 mg/ kg铬时,CrCl3组显著低于Cr-Pic和Cr-Met组(P<0.05)。
成活率:Cr-Pic组的成活率显著高于CrCl3和Cr-Met组(P<0.05)。
饲料系数:以CrCl3为铬源时,0.9 mg/ kg组最低,0、0.3 mg/ kg组显著高于0.9、1.2 mg/ kg组(P<0.05),其余各组间差异不显著(P>0.05);以Cr-Pic为铬源时,1.2 mg/ kg组最低,0 mg/ kg组最高,除0.3、0.6 mg/ kg组之间无显著差异(P>0.05)外,其余各组间差异显著(P<0.05);以Cr-Met为铬源时,0.9 mg/ kg组最低,0 mg/ kg组最高,0.9 mg/ kg组显著低于除0.6、1.2 mg/ kg组外的其余各组(P<0.05)。添加0.6、0.9 mg/ kg铬时,Cr-Met组显著低于CrCl3和Cr-Pic组(P<0.05);添加1.2、2.0 mg/ kg铬时,CrCl3组显著高于Cr-Pic和Cr-Met组(P<0.05)。
蛋白质效率:以CrCl3为铬源时,1.2 mg/ kg组最高,0 mg/ kg组最低,且2.0 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic和Cr-Met为铬源时,0.9 mg/ kg组最高,1.2 mg/ kg组次之,0 mg/ kg组最低,0. 9和1. 2 mg/ kg组显著高于0、0.3 mg/ kg组(P<0. 05)。铬的各添加水平下,CrCl3组均显著低于Cr-Pic和Cr-Met组(P<0.05)。
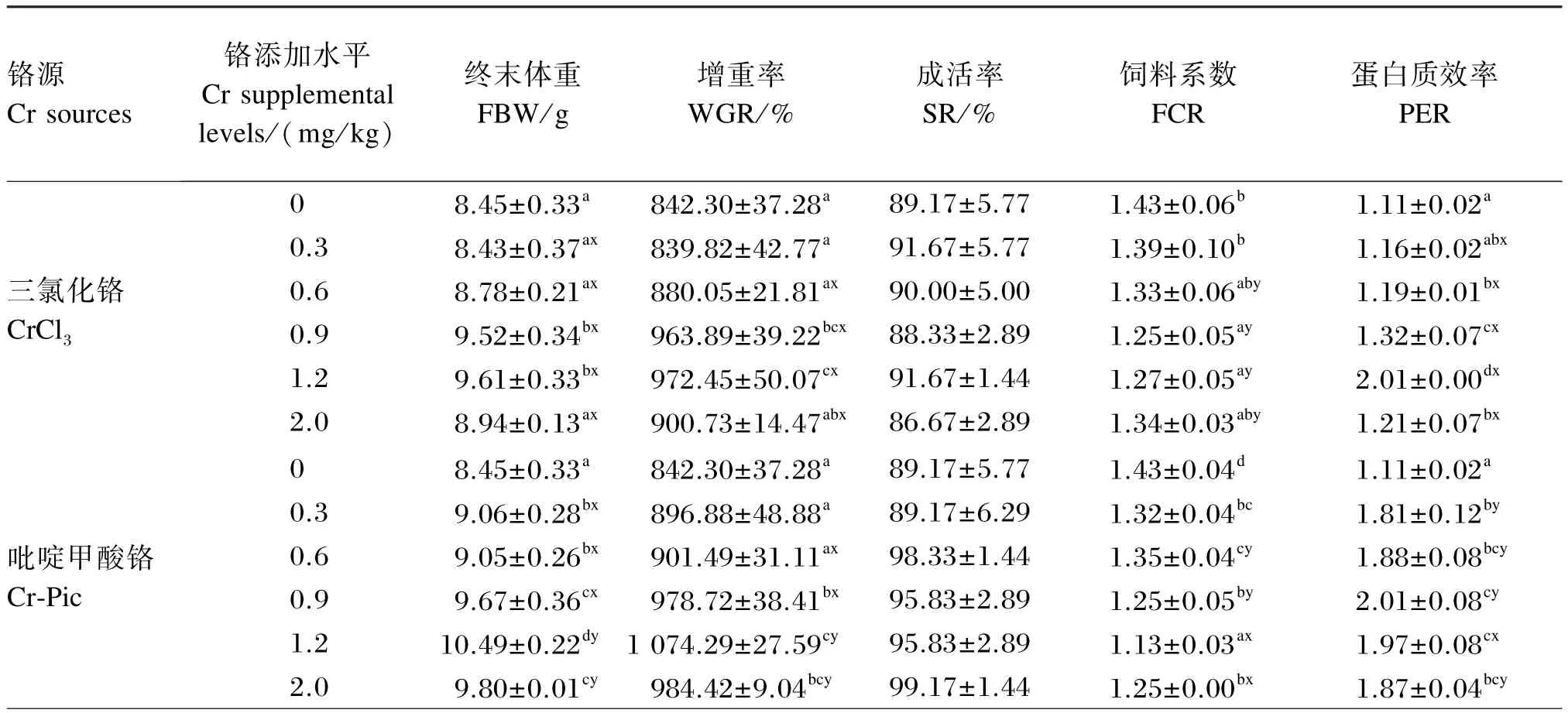
表3 饲料中铬源及铬添加水平对凡纳滨对虾幼虾生长性能的影响Table 3 Effects of dietary chromium source and supplemental level on growth performance of juvenile Litopenaeus vannamei(n=3)
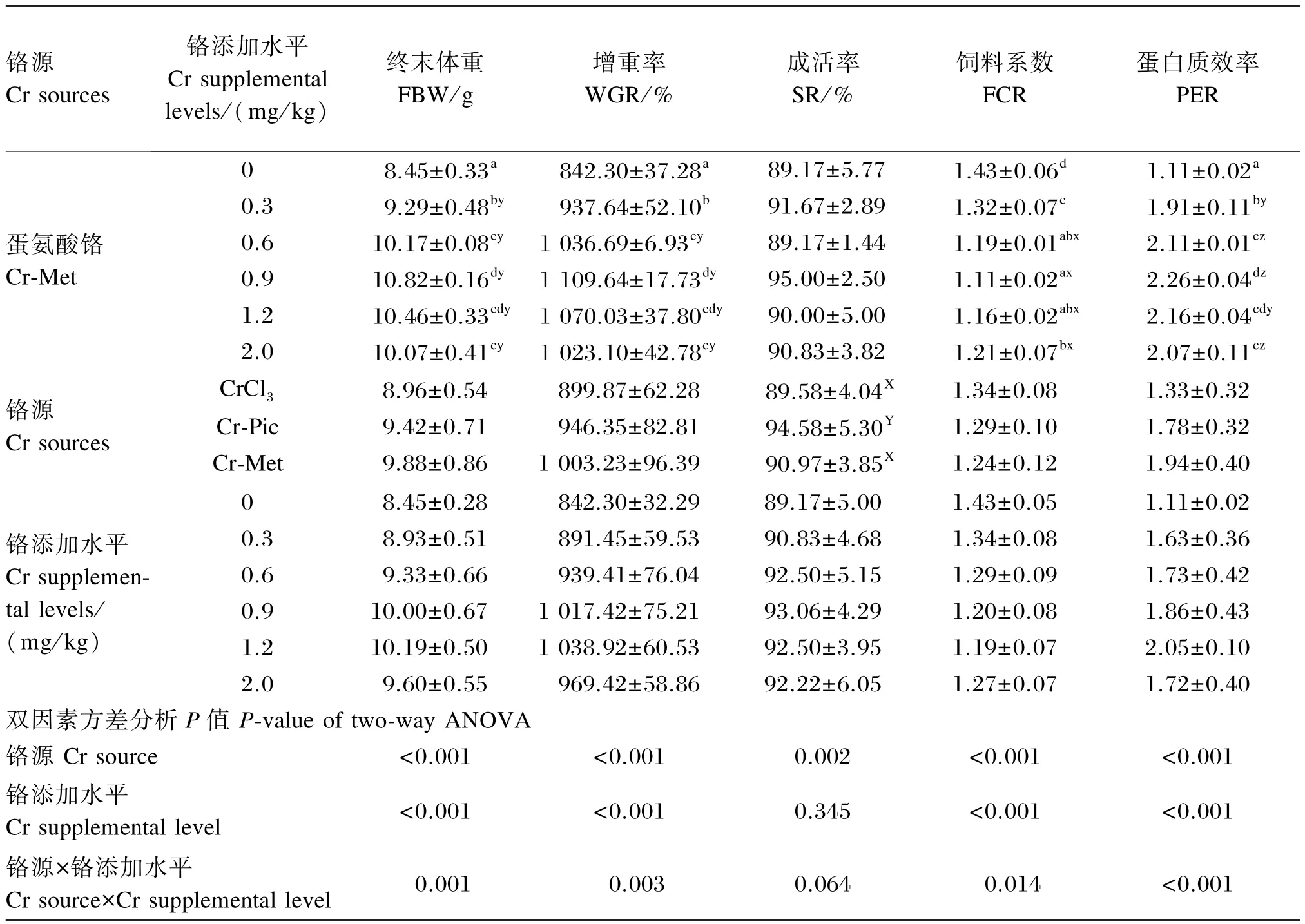
续表3
2.2饲料中铬源及铬添加水平对凡纳滨对虾幼虾体成分的影响
由表4可知,铬源及铬添加水平对凡纳滨对虾全虾的粗脂肪、粗灰分含量有显著影响(P<0.05),且铬源与铬添加水平之间存在交互作用(P <0.05);铬源及铬添加水平对凡纳滨对虾全虾的水分和粗蛋白质含量无显著影响(P>0.05),且两者间无交互作用(P>0.05)。
粗脂肪含量:以CrCl3为铬源时,1.2 mg/ kg组显著低于其余各组(P<0.05);以Cr-Pic和Cr-Met为铬源时,0.9、1.2 mg/ kg组显著低于其余各组(P<0.05)。添加0.6 mg/ kg铬时,Cr-Met组显著低于CrCl3和Cr-Pic组(P<0.05);添加0.9 mg/ kg铬时,CrCl3组显著高于Cr-Pic和Cr-Met组(P<0.05)。
粗灰分含量:以CrCl3为铬源时,0 mg/ kg组显著低于其余各组(P<0.05);以Cr-Pic为铬源时,0、0.3 mg/ kg组显著低于其余各组(P<0.05);以Cr-Met为铬源时,0.9 mg/ kg组最高,显著高于其余各组(P<0.05)。添加0.6、0.9、1.2和2.0 mg/ kg铬时,Cr-Pic组显著高于CrCl3和Cr-Met组(P<0.05)。

表4 饲料中铬源及铬添加水平对凡纳滨对虾幼虾体成分的影响(干物质基础)Table 4 Effects of dietary chromium source and supplemental level on body composition of juvenile Litopenaeus vannamei(DM basis,n=3) %
2.3饲料中铬源及铬添加水平对凡纳滨对虾幼虾血清生化指标的影响
由表5可知,铬源及铬添加水平对凡纳滨对虾血清中总蛋白、葡萄糖、胆固醇和甘油三酯含量均有显著影响(P<0.05),且铬源与铬添加水平之间存在交互作用(P<0.05)。
总蛋白含量:以CrCl3和Cr-Pic为铬源时,0.9、1.2和2.0 mg/ kg组显著高于其余各组(P<0.05);以Cr-Met为铬源时,0.9、1.2 mg/ kg组显著高于其余各组(P<0. 05)。添加0. 6、0. 9和1.2 mg/ kg铬时,Cr-Met组显著高于CrCl3和Cr-Pic组(P<0.05);添加2.0 mg/ kg铬时,CrCl3组显著低于Cr-Pic和Cr-Met组(P<0.05)。
葡萄糖含量:以CrCl3和Cr-Pic为铬源时,0、0.3 mg/ kg组显著高于其余各组(P<0.05);以Cr-Met为铬源时,0 mg/ kg组显著高于其余各组(P<0.05)。铬的各添加水平下,Cr-Met组均显著低于CrCl3和Cr-Pic组(P<0.05)。
胆固醇含量:以CrCl3为铬源时,0 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic为铬源时,0.3 mg/ kg组显著高于其余各组(P<0.05);以Cr-Met为铬源时,0、0.3 mg/ kg组显著高于其余各组(P<0.05)。添加0.3 mg/ kg铬时,Cr-Met组显著高于CrCl3和Cr-Pic组(P<0. 05);添加0. 9、2.0 mg/ kg铬时,CrCl3组显著高于Cr-Pic和Cr-Met组(P<0.05)。
甘油三酯含量:以CrCl3为铬源时,0、0.3 mg/ kg组显著低于其余各组(P<0.05);以Cr-Pic为铬源时,2.0 mg/ kg组显著高于其余各组(P<0.05);以Cr-Met为铬源时,1.2、2.0 mg/ kg组显著高于其余各组(P<0.05)。添加0.6 mg/ kg铬时,CrCl3组显著高于Cr-Pic和Cr-Met组(P<0.05);添加1.2、2.0 mg/ kg铬时,Cr-Met组显著高于CrCl3和Cr-Pic组(P<0.05)。

表5 饲料中铬源及铬添加水平对凡纳滨对虾幼虾血清生化指标的影响Table 5 Effects of dietary chromium source and supplemental level on serum biochemical indices of juvenile Litopenaeus vannamei(n=3)
2.4饲料中铬源及铬添加水平对凡纳滨对虾幼虾血清中非特异性免疫酶活性的影响
由表6可知,铬源及铬添加水平对凡纳滨对虾血清中酚氧化物酶、总超氧化物歧化酶、碱性磷酸酶、酸性磷酸酶的活性均有显著影响(P<0.05),且铬源与铬添加水平之间存在交互作用(P<0.05)。
酚氧化物酶活性:以CrCl3为铬源时,0.9、1.2 和2.0 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic为铬源时,0.9、1.2 mg/ kg组显著高于其余各组(P<0. 05);以Cr-Met为铬源时,0. 6、0.9 mg/ kg组显著高于其余各组(P<0.05)。铬的各添加水平下,Cr-Met组均显著高于CrCl3和Cr-Pic组(P<0.05)。
总超氧化物歧化酶活性:以CrCl3为铬源时,1.2 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic和Cr-Met为铬源时,0.9 mg/ kg组显著高于其余各组(P<0.05)。添加0.3、0.9 mg/ kg铬时,Cr-Met组显著高于CrCl3和Cr-Pic组(P<0.05);添加1.2 mg/ kg铬时,CrCl3组显著高于Cr-Pic和Cr-Met组(P<0.05)。
碱性磷酸酶活性:以CrCl3为铬源时,0.9、1.2 和2.0 mg/ kg组显著高于其余各组(P<0.05);以Cr-Pic为铬源时,1.2 mg/ kg组显著高于其余各组(P<0.05);以Cr-Met为铬源时,1.2、2.0 mg/ kg组显著高于除0. 9 mg/ kg组外的其余各组(P<0.05)。添加0.3、1.2 mg/ kg铬时,CrCl3组显著低于Cr-Pic和Cr-Met组(P<0.05);添加0.9 mg/ kg铬时,Cr-Pic组显著低于CrCl3和Cr-Met组(P<0.05)。
酸性磷酸酶活性:以CrCl3和Cr-Pic为铬源时,0.9、1. 2 mg/ kg组显著高于其余各组(P<0.05);以Cr-Met为铬源时,1.2 mg/ kg组显著高于其余各组(P<0.05)。添加0.3 mg/ kg铬时,Cr-Pic组显著高于CrCl3和Cr-Met组(P<0.05);添加0.6、2.0 mg/ kg铬时,Cr-Pic组显著低于CrCl3和Cr-Met组(P<0.05),添加1.2 mg/ kg铬时,CrCl3组显著高于Cr-Pic和Cr-Met组(P<0.05)。
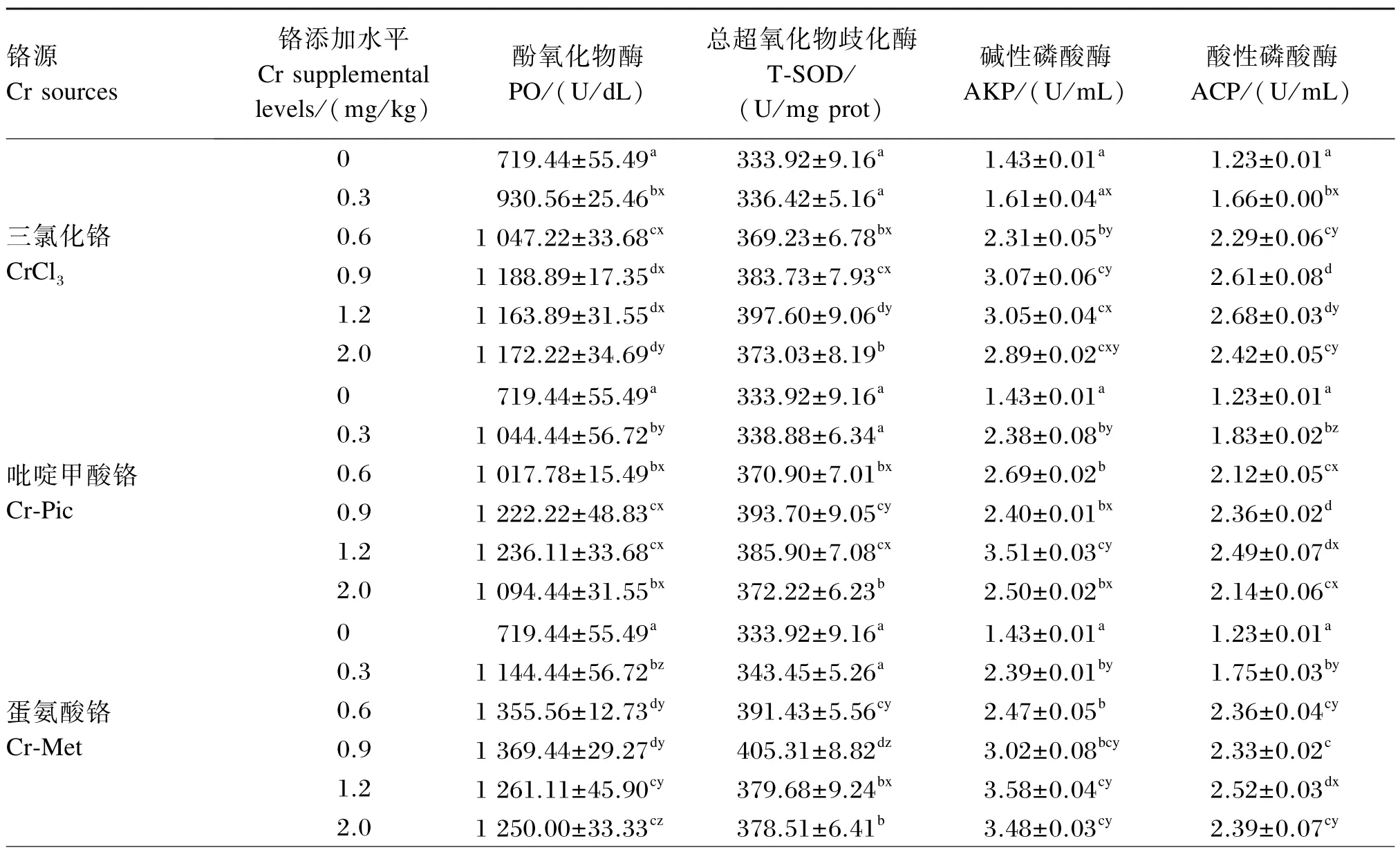
表6 饲料中铬源及铬添加水平对凡纳滨对虾幼虾血清中非特异性免疫酶活性的影响Table 6 Effects of dietary chromium source and supplemental level on serum non-specific immune enzyme activities of juvenile Litopenaeus vannamei(n=3)
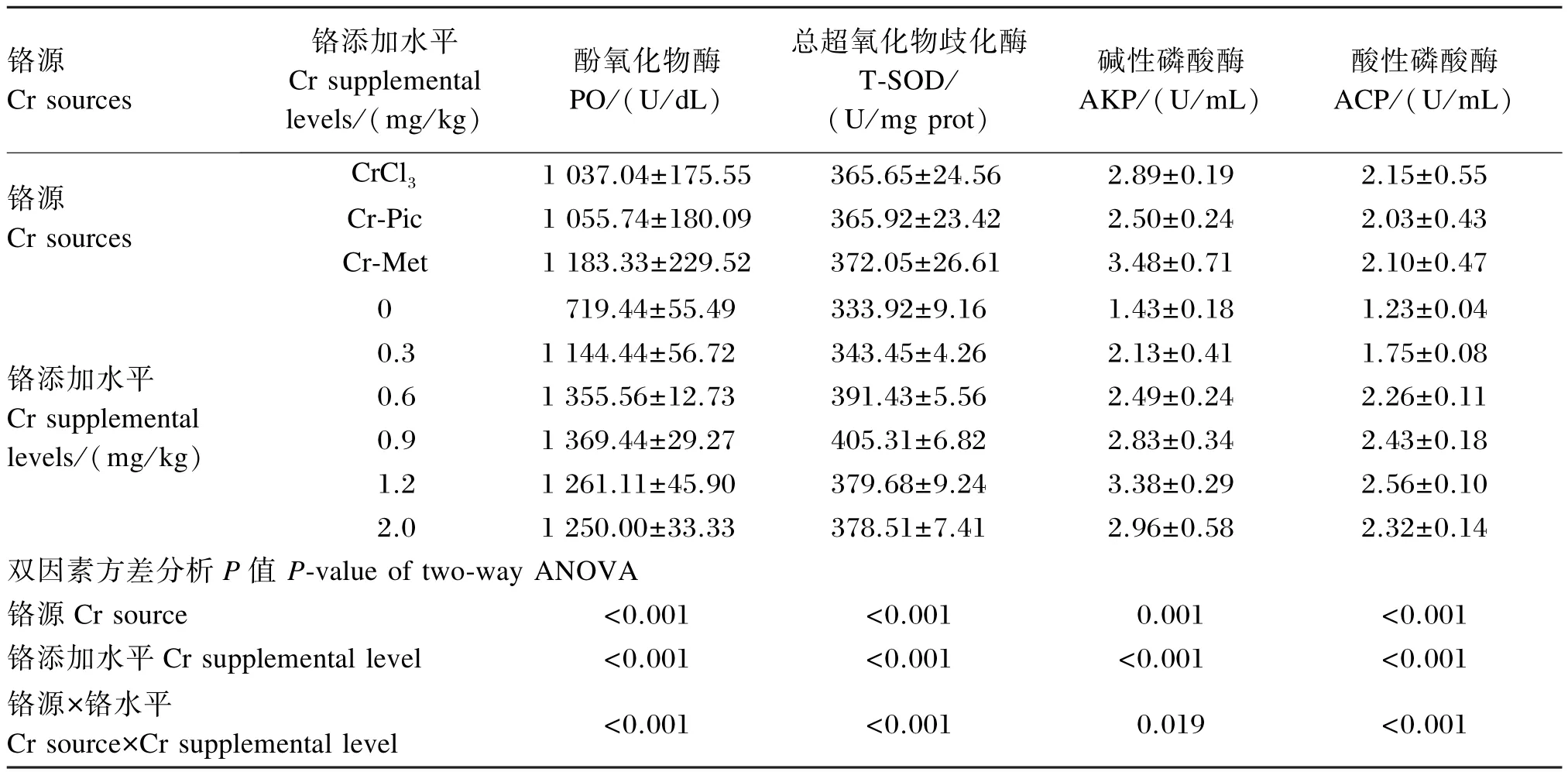
续表6
2.53种铬源相对生物利用率的比较
由表7和图1可知,以增重率为评价指标,以CrCl3、Cr-Pic、Cr-Met为铬源时,经折线模型得出饲料中铬的适宜添加水平分别为1.33、1.27、 1.04 mg/ kg。以CrCl3为标准物时,Cr-Pic和Cr-Met的相对生物利用率分别为124. 20%、184.32%。

表7 以增重率为评价指标时吡啶甲酸铬和蛋氨酸铬的相对生物利用率Table 7 Relative bioavailability of Cr-Pic and Cr-Met for WGR as an evaluation index

图1 饲料中铬添加水平与凡纳滨对虾幼虾增重率的关系Fig.1 Relationship of dietary chromium supplemental level and WGR of juvenile Litopenaeus vannamei
3 讨 论
3.1铬源及铬添加水平对凡纳滨对虾幼虾生长性能的影响
铬是动物必需的微量元素,在水产动物饲料中添加一定量的铬盐能增强水产动物的生长性能。Cr3+作为GTF的活性成分,与胰岛素之间存在协同作用[14],共同参与体内三大营养物质的代谢[15],从而促进机体的生长和增重[16]。
目前,关于饲料中添加铬对虾类生长性能影响的研究较少,但在鱼类上的报道较多。研究表明,饲料中添加1.2~1.6 mg/ kg Cr-Pic时,可提高凡纳滨对虾的生长和饲料利用率[9];饲料中添加2.0%的Cr-Pic可提高奥尼罗非鱼的生长和饲料利用率[6];添加1.7 mg/ kg的烟酸铬、甘氨酸铬和Cr-Met均可促进奥尼罗非鱼的生长,并降低饲料系数[17];饲料中以Cr-Pic的形式添加0.8 mg/ kg的铬可显著提高吉富罗非鱼的生长和饲料利用率[7];饲料中添加Cr-Pic能提高草鱼对葡萄糖的耐受量,促进草鱼生长[3];在基础饲料中添加600 μg/ kg Cr-Pic,团头鲂可获得较好的生长性能[8]。
本试验结果表明,当饲料中以CrCl3和Cr-Pic的形式添加1.2 mg/ kg铬,以Cr-Met的形式添加0.9 mg/ kg铬时,可显著提高凡纳滨对虾的增重率、蛋白质效率,并显著降低饲料系数。本试验结果与上述研究结果相似,表明饲料中添加一定量的铬可促进动物的生长和饲料利用率,且不同添加形式的铬对动物生长性能的作用效果不同。影响动物对微量元素吸收利用效果的因素相当复杂,除了受动物本身消化吸收生理特点的影响外,可能还和不同形式微量元素的分子结构、分子量、溶解度等有关。由于目前国内外对甲壳动物微量元素具体吸收机制的研究很少,对不同形式微量元素的实际应用技术比较盲目,还需要开展大量的研究工作,以期弥补有关微量元素方面的空白。
3.2铬源及铬添加水平对凡纳滨对虾幼虾体成分的影响
本试验中,全虾水分和粗蛋白质含量在各组之间无显著差异,这与在罗非鱼[18]和草鱼[3]上的研究结果一致。关于铬对动物体粗脂肪和粗灰分含量影响的研究,不同的研究所得结果有所差异。Xi等[19]报道,在生长肥育猪饲粮中添加铬,通过降低脂肪合成酶活性,从而减少了脂肪的沉积。刘太亮等[3]在草鱼饲料中添加铬后发现全鱼的粗脂肪含量并无显著变化,添加铬试验组的全鱼粗灰分含量显著高于对照组。杨奇慧等[9]研究表明,全虾粗脂肪含量随Cr-Pic添加水平的增加有下降的趋势,而粗灰分含量则有增加的趋势。崔学升[20]研究发现,添加Cr-Pic可降低幼建鲤体脂肪的含量。本研究中全虾粗脂肪含量在3种铬源下均有随着铬添加水平的增加而下降的趋势,这一结果与Xi等[19]、杨奇慧等[9]和崔学升[20]的研究结果一致。铬作用于胰岛素以及脂肪代谢有关的酶,从而调节脂肪的代谢,这可能是全虾粗脂肪含量降低的原因。本试验结果表明,全虾粗灰分含量在3种铬源下均随着铬添加量的增加而增加,这与刘太亮等[3]和杨奇慧等[9]的研究结果一致。关于铬对动物体成分影响的研究,由于动物品种、规格、饲料组成、养殖环境等因素的影响,研究结果有很大差异。因此,铬对动物体成分影响的研究还有待深入的研究。
3.3铬源及铬添加水平对凡纳滨对虾幼虾血清生化指标的影响
本试验中,在3种铬源下,随着铬添加水平的增加,血清葡萄糖含量呈递减趋势,这与杨奇慧等[7]、刘太亮等[3]和蔡春芳等[21]的报道结果一致。蔡春芳等[21]报道,在葡萄糖为糖原的饲料中添加铬显著地提高了异育银鲫的糖耐量,对淀粉组的糖耐量也有一定的促进作用;刘太亮等[3]研究发现,在饲料中添加0.8 mg/ kg Cr-Pic后草鱼的葡萄糖耐量显著提高;杨奇慧等[7]研究指出,以Cr-Pic形式在饲料中添加0.8 mg/ kg的铬可显著降低吉富罗非鱼的血清葡萄糖含量。铬是GTF的主要组成部分,GTF能增加细胞表面胰岛素受体数量以促进胰岛素与特定受体的结合,刺激组织细胞对葡萄糖的摄取,从而使血液中葡萄糖的含量降低[22]。关于铬对血糖调节作用的研究,在人类和陆生动物中比较深入,在水产动物方面还有待进一步研究。
血清总蛋白含量在一定程度上反映了饲养动物所摄食的饲料中蛋白质的营养水平以及动物对蛋白质的消化吸收程度,其与机体自身的蛋白质合成机能和氮的沉积效果息息相关[23]。Amoikon等[24]认为铬增强胰岛素的功能后,胰岛素通过促进氨基酸进入细胞,从而促进蛋白质的合成。本试验中,在3种铬源下,血清总蛋白含量随着铬添加水平的增加而增加,添加0.9、1.2 mg/ kg Cr-Met组对虾的血清总蛋白含量最高。这可能与氨基酸螯合物的添加增加了饲料中必需氨基酸的量,优化了必需氨基酸的比例以及促进了Cr3+的吸收等有关。本试验结果与吴永胜等[25]、杨奇慧等[9]的研究成果基本一致,但蔺玉华等[26]用添加铬的饲料喂养鲤,其血清中总蛋白含量低于对照组,与本试验的结论不一致,其原因可能是由于铬的添加促使氨基酸向组织蛋白转化,从而导致血清总蛋白含量下降,但具体作用机制还需进一步研究。
铬对脂肪代谢的作用主要是维持血液的正常胆固醇含量[27],影响脂肪和胆固醇在动物肝脏中的合成与清除,促进脂类重分配。在畜禽上的研究表明,饲料添加有机铬可降低血清中胆固醇含量[28],提高血清中甘油三酯含量[29]。刘太亮[30]研究表明,添加Cr-Pic和烟酸铬能显著降低草鱼血清中胆固醇含量,提高血清中甘油三酯含量。杨奇慧等[9]研究指出,添加Cr-Pic可降低凡纳滨对虾血清中胆固醇含量,而对血清中甘油三酯含量无显著影响。蔺玉华等[26]在研究中证实铬有降低血清甘油三酯含量的作用。本试验中,添加铬能够降低凡纳滨对虾血清中胆固醇含量,提高血清中甘油三酯含量,这一结果与刘太亮[30]的研究结果一致。血清中胆固醇含量的减少,可能是因为铬不但降低血清中胆固醇的累积,而且可加快主动脉内已沉积的胆固醇的迁移[31]。有关铬对血清甘油三酯含量的影响目前说法不一,有研究认为甘油三酯含量的下降可能是由于氧化反应增强,尤其是β-氧化过程,从而增强脂肪氧化供能途径,减少脂肪酸合成甘油三酯,从而导致其含量的降低[25];Mertz等[10]亦认为GTF是胰岛素的增强剂,而胰岛素促进血糖进入肝脏,骨骼肌和脂肪细胞转化为糖原和甘油三酯,并不是以脂肪的形式沉积下来,这也可能是本试验中导致血清甘油三酯含量升高的原因之一。因此,有关铬对动物血清中脂类代谢的影响仍需进一步的探讨。
3.4铬源及铬添加水平对凡纳滨对虾血清中非特异性免疫酶活性的影响
铬不仅可促进动物的生长,还可增强机体的免疫力。本试验结果表明,饲料中适量添加铬可显著提高血清碱性磷酸酶、酸性磷酸酶、酚氧化物酶以及总超氧化物歧化酶的活性。
碱性磷酸酶是一种特异性较低的膜结合酶,在动物机体中存在较为广泛,主要功能是参与体内物质(如磷酸基团)的转运及代谢、水产动物骨骼的矿化过程[32]。酸性磷酸酶是巨噬细胞溶酶体的标志酶,和碱性磷酸酶一样能够催化磷酸单脂的水解,在体内直接参与磷酸基团的转移与代谢过程[33]。本研究表明,3种铬源均显著提高凡纳滨对虾血清中碱性磷酸酶、酸性磷酸酶活性,随铬添加水平的增加逐渐趋于平稳,与周燕等[34]、杨奇慧等[9]的报道相似。
酚氧化物酶是一种重要的防御酶,具有异物识别作用,可比较敏感地反映机体的免疫状态[35-36]。超氧化物歧化酶具有清除自由基以及催化过氧化物自由基歧化为过氧化物和氧气的功能[37]。本试验中,3种铬源均显著提高血清中酚氧化物酶及总超氧化物歧化酶活性。以CrCl3和Cr-Met为铬源时,酚氧化物酶均在添加0.9 mg/ kg铬时表现出最高活性,以Cr-Pic为铬源时,酚氧化物酶在添加1.2 mg/ kg铬时表现出最高活性。以CrCl3为铬源时,总超氧化物歧化酶在添加1.2 mg/ kg铬时表现出最高活性,以Cr-Pic和Cr-Met为铬源时,总超氧化物歧化酶在添加0.9 mg/ kg铬时表现出最高活性。杨奇慧等[9]发现,以Cr-Pic的形式添加1.6 mg/ kg铬时,血清中酚氧化物酶和总超氧化物歧化酶活性达到最高值。但杨允辉等[5]研究表明,添加铬对河蟹血清中酚氧化物酶活性无显著影响。可见,铬对水产动物非特异性免疫功能影响的结果差异较大,其原因可能与试验动物的品种、大小及铬的来源、添加形式和水平等不同有关,具体原因还有待进一步的研究。
4 结 论
以增重率为评价指标,以CrCl3、Cr-Pic和Cr-Met为铬源时,经折线模型得出饲料中铬的适宜添加水平分别为1.33、1.27、1.04 mg/ kg。通过比较可知,Cr-Met的相对生物利用率最高,Cr-Pic次之,CrCl3最低。
参考文献:
[1] 阎小艳,楮秋霞,陈庆林,等.微量元素铬的营养研究进展[J].山西农业科学,2007,35(11):17-21.
[2] 邓国彬.浅谈酵母铬对鲤鱼生产性能的影响[J].饲料工业,1999,20(4):41-42.
[3] 刘太亮,蒋明,文华,等.吡啶羧酸铬对草鱼生长和糖耐量的影响[J].华中农业大学学报,2009,28(2):198-201.
[4] 刘玉林,李浪平.丙酸铬对草鱼生长性能及血液生化指标的影响[J].科学养鱼,2010(4):65-66.
[5] 杨允辉,周永奎,史合群,等.饲料中添加铬对河蟹非特异性免疫力和抗应激能力的影响[J].淡水渔业,2006,36(5):11-15.
[6] 潘庆,毕英佐,颜惜玲,等.有机铬对奥尼罗非鱼生长和糖利用的影响[J].水生生物学报,2004,26(4):393-399.
[7] 杨奇慧,谭北平,董晓慧,等.吡啶甲酸铬对吉富罗非鱼生长、饲料利用和非特异性免疫功能的影响[J].中国饲料,2012(1):26-32.
[8] 周兴华,陈建.吡啶甲酸铬对团头鲂生产性能的影响[J].兽药与饲料添加剂,2002,7(4):3-4.
[9] 杨奇慧,谭北平,董晓慧,等.铬对凡纳滨对虾生长性能、血清生化指标及非特异性免疫酶活性的影响[J].动物营养学报,2013,25(4):795-804.
[10] MERTZ W. Chromium in human nutrition:a review [J]. The Journal of Nutrition,1993,123(4):626 -633.
[11] AOAC.Official methods of analysis of AOAC International[S].16th ed.Arlington,VA:Association of Analytical Communities,1995.
[12] 王建国,陆宏达.酚氧化酶活力测定方法中关于测定时间的研究[J].上海海洋大学学报,2009,18(6):765-770.
[13] HUANG J W,YANG Y,WANG A L. Reconsideration of phenoloxidase activity determination in white shrimp Litopenaeus vannamei[J].Fish&Shellfish Immunology,2010,28(1):240-244.
[14] ANDERSON R A.Trace elements in human and animal nutrition[M].5th ed.New York:Academic Press,1987:225.
[15] NIELSO F H.Mordern nutrition in health and disease [M]. Philadelphia:Philadelphia Education Press,1994:113.
[16] MERTZ W.Chromium occurrence and function in biological systems[J]. Physiological Reviews,1969,49 (2):163-239.
[17] 蒋伟明,韩耀全,李大列.不同铬化合物对奥尼罗非鱼生长的影响[J].广西农业科学,2004,35(1):76-78.
[18] PAN Q,LIU S,TAN Y G,et al.The effect of chromium picolinate on growth and carbohydrate utilization in tilapia,Oreochromis niloticus×Oreochromis aureus [J].Aquaculture,2003,225(1/2/3/4):421-429.
[19] XI G,XU Z R,WU S H,et al.Effect of chromium picolinate on growth performance,carcass characteristics,serum metabolites and metabolism of lipid in pigs [J]. Asian-Australasian Journal of Animal Sciences,2001,14(2):258-262.
[20] 崔学升.添加吡啶羧酸铬对幼建鲤体成分和机体非特异免疫功能的影响[D].硕士学位论文.雅安:四川农业大学,2010.
[21] 蔡春芳,宋学宏,王永玲.不同糖源及铬对异育银鲫生长和糖耐量的影响[J].水产学报,1999,23(4):432-434.
[22] HERTZ Y,MADAR Z,HEPHER B,et al. Glucose metabolism in the common carp(Cyprinus carpio L.):the effects of cobalt and chromium[J].Aquaculture,1989,76(3/4):255-267.
[23] LUO X G,DOVE C R.Effect of dietary copper and fat on nutrient utilization,digestive enzyme activities,and tissue mineral levels in weanling pigs[J].Journal of Animal Science,1996,74(8):1888-1896.
[24] AMOIKON E K,FERNANDEZ J M,SOUTHERN L L,et al. Effect of chromium tripicolinate on growth,glucose tolerance,insulin sensitivity,plasma metabolites,and growth hormone in pigs[J].Journal of Animal Science,1995,73(4):1123-1130.
[25] 吴永胜,董国忠,王立常,等.饲粮中添加铬——烟酸复合物对肉鸭生产性能、胴体成分及血液生化指标的影响[J].动物营养学报,2000,12(2):20-25.
[26] 蔺玉华,卢健民,富惠光,等.不同浓度铬盐饲料对鲤生长和血浆生化指标的影响[J].大连水产学院学报,2003,18(1):48-51.
[27] PAGE T G,SOUTHERN L L,WARD T L,et al. Effect of chromium picolinate on growth and serum and carcass traits of growing-finishing pigs[J].Journal of Animal Science,1993,71(3):656-662.
[28] KIM J D,HAN I K,CHAE B J,et al.Effects of dietary chromium picolinate on performance,egg,quality,serum traits and mortality rate of brown layers[J].A-sian-Australasian Journal of Animal Sciences,1997,10(1):1-7.
[29] 李丽立,钟华宜,张彬.酵母铬对肉鸭生产性能和生化指标的影响[J].饲料研究,2001(1):30-32.
[30] 刘太亮.草鱼对饲料中铬需要量的研究[D].硕士学位论文.武汉:华中农业大学,2009.
[31] ABRAHAM A S,SONNENBLICK M,EINI M,et al. The effect of chromium on established atheroscleroticplaques in rabbits[J].The American Journal of Clinical Nutrition,1980,33(11):2294-2298.
[32] BLASCO J,PUPPO J,SARASQUETE M C.Acid and alkaline phosphatase activities in the clam Ruditapes philippinarum[J]. Marine Biology,1993,115(1):113-118.
[33] 刘树青,江晓路,牟海津,等.免疫多糖对中国对虾血清溶菌酶,磷酸酶和过氧化物酶的作用[J].海洋与湖沼,1999,30(3):278-283.
[34] 周燕,崔学升,李志琼,等.吡啶羧酸铬对幼建鲤非特异性免疫功能的影响[J].饲料研究,2011(9):67-69.
(责任编辑菅景颖)
[35] SÖDERHÄLL K,CERENIUS L.Role of the prophenoloxidase-activating system in invertebrate immunity [J].Current Opinion in Immunology,1998,10(1):23-28.
[36] CHENG W,JUANG F M,CHEN J C.The immune response of Taiwan abalone Haliotis diversicolor supertexta and its susceptibility to Vibrio parahaemolyticus at different salinity levels[J].Fish&Shellfish Immunology,2004,16(3):295-306.
[37] FATTMAN C L,SCHAEFER L M,OURY T D.Extracellular superoxide dismutase in biology and medicine[J].Free Radical Biology and Medicine,2003,35 (3):236-256.
Effects of Dietary Chromium Source and Supplemental Level on Growth Performance,Serum Biochemical Indices and Non-Specific Immune Enzyme Activities of Juvenile Litopenaeus vannamei
CAI Hairui TAN Beiping∗YANG Qihui∗DONG Xiaohui CHI Shuyan LIU Hongyu ZHANG Shuang
(Laboratory of Aquatic Animal Nutrition and Feed,College of Fisheries,Guangdong Ocean University,Zhanjiang 524088,China)
Abstract:This experiment was conducted to study the effects of dietary chromium source and supplemental level on growth performance,serum biochemical indices and non-specific immune enzyme activities of juvenile Litopenaeus vannamei,and to find the optimal supplemental levels of three chromium in the diets. Sixteen isonitrogenous and isoenergetic diets were formulated by using two-factor experimental design with three chromium sources[chromium trichloride(CrCl3),chromium picolinate(Cr-Pic)and chromium methionine (Cr-Met),respectively]and six chromium supplemental levels(0,0.3,0.6,0.9,1.2 and 2.0 mg/ kg,respectively). A total of 1 920 juvenile Litopenaeus vannamei with an initial body weight of(0.897±0.001)g were randomly distributed into 16 groups with 3 replicates per group and 40 shrimps per replicate. The experiment lasted for 8 weeks. The results showed that final body weight(FBW),weight gain rate(WGR),feed conversion rate(FCR)and protein efficiency ratio(PER)of shrimps were significantly affected by chromium source,supplemental level and the interaction of chromium source and supplemental level,respectively(P<0.05). The FBW and WGR in groups supplemented with 0.3 to 2.0 mg/ kg chromium were significantly higher than those in the group without chromium(P<0.05),and the Cr-Met group supplemented with 0.9 mg/ kg chromium had the highest FBW and WGR. The groups of CrCl3and Cr-Pic supplemented with 0.9 mg/ kg chromium had the lowest FCR,but showed no significant differences compared with 1.2 mg/ kg group(P>0.05). The highest PER of three chromium sources were founded in the groups supplemented with 0.9 mg/ kg chromium,but showed no significant differences compared with the groups supplemented with 1.2 mg/ kg chromium(P>0.05). Chromium source,supplemental level and the interaction of chromium source and sup-book=779,ebook=152plemental level had significant effects on the crude lipid and crude ash contents in whole body(P<0.05),but had no significant effects on the moisture and crude protein contents in whole body(P>0.05). Serum total protein(TP),glucose,cholesterol and triglyceride contents were significantly affected by chromium source,supplemental level and the interaction of chromium source and supplemental level,respectively(P<0.05). The highest TP content and lowest glucose content in serum were found in the Cr-Met group supplemented with 0.9 mg/ kg chromium. The activities of phenoloxidase(PO),alkaline phosphatase(AKP),acid phosphatase (ACP)and the total superoxide dismutase(T-SOD)in serum were significantly affected by chromium source,supplemental level and the interaction of chromium source and supplemental level,respectively(P<0.05). The highest serum PO and T-SOD activities were found in the Cr-Met group supplemented with 0.9 mg/ kg chromium. With the WGR as an evaluation index and the CrCl3,Cr-Pic and Cr-Met as chromium sources,brokenline model shows that the optimal supplemental levels of chromium in juvenile Litopenaeus vannamei diets are 1.33,1.27 and 1.04 mg/ kg,respectively. By comparing,the Cr-Met has the highest relative bioavailability,followed by Cr-Pic,and the lowest is CrCl3.[Chinese Journal of Animal Nutrition,2016,28(3):766-779]
Key words:Litopenaeus vannamei;chromium;growth performance;serum biochemical indices;non-specific immune enzyme activities
Corresponding author∗s:TAN Beiping,professor,E-mail:bptan@126.com;YANG Qihui,professor,E-mail:qihuiyang03@163.com
通信作者:∗谭北平,教授,博士生导师,E-mail:bptan@126.com;杨奇慧,教授,硕士生导师,E-mail:qihuiyang03@163.com
作者简介:蔡海瑞(1989—),女,河南驻马店人,硕士研究生,研究方向为水产动物营养与饲料。E-mail:caicai830520@163.com
基金项目:公益性行业(农业)科研专项(201003020);广东省自然科学基金(2015A030313621);广东省产业技术与开发专项资金项目(2013B021100017)
收稿日期:2015-10-15
doi:10.3969/ j.issn.1006-267x.2016.03.017
中图分类号:S963
文献标识码:A
文章编号:1006-267X(2016)03-0766-14
猜你喜欢
热带农业科学(2016年12期)2017-03-31
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2016年11期)2017-03-21
湖北农业科学(2016年23期)2017-03-17
湖北农业科学(2016年18期)2016-12-08
河北渔业(2016年10期)2016-11-22
湖南饲料(2016年2期)2016-05-12
江苏农业科学(2015年8期)2015-09-10
河北渔业(2015年2期)2015-03-26
