
3种不同养殖模式对精养水体尼罗罗非鱼生长性能及养殖水质的影响
2017-03-21 08:29方彰胜刘邦辉王广军
江苏农业科学 2016年11期
方彰胜+刘邦辉+王广军
摘要:以尼罗罗非鱼为研究对象,采用投放光合細菌、安装生物浮床单养和罗非鱼、鲫鱼、鳙鱼混养等3种不同模式对尼罗罗非鱼精准养殖水体水质和生长等方面进行研究。结果显示,在70 d养殖周期内,除混养组合和对照组间的平均增质量率无显著差异(P>0.05)外,光合细菌和生物浮床使罗非鱼的生长和成活率有显著提高(P<0.05);3种养殖模式对试验周期内罗非鱼精养水体中磷酸盐和中亚硝酸盐浓度均具有不同程度降低作用,光合细菌组的作用尤其显著(P<0.05),其后依次是生物浮床组、混养组和对照组;3种养殖模式对硝酸盐浓度无显著影响(P>0.05),3种养殖模式氨氮含量在养殖周期内均呈现先升高后降低的规律,整个养殖周期内,光合细菌组水体中氨氮含量均显著低于其他组(P<0.05),对照组水体中氨氮含量始终高于其他组(P<0.05)。结果表明,3种不同养殖模式在改善罗非鱼养殖水质的前提下,对罗非鱼的生长和成活率有显著提高(P<0.05)作用。同时,此几种模式可为推动我国罗非鱼发展模式由资源高耗费型向环境友好型转变提供新的途径和思路,不失为罗非鱼生态健康高效的养殖模式。
关键词:尼罗罗非鱼;养殖模式;生长性能;水质
中图分类号: S965.125 文献标志码: A
文章编号:1002-1302(2016)11-0252-04
罗非鱼(Oreochromis)是原产于热带亚热带暖水性鱼类,有尼罗、莫桑比克、奥利亚等10多个品种,因其具有繁殖力强、生长速度快、耐粗食、抗病力强等优点,是我国特别是华南地区重要的淡水养殖和出口创汇品种[1]。其中,养殖模式的发展和完善对我国罗非鱼养殖发展起着积极的促进作用。目前,罗非鱼的养殖模式主要有池塘混养、网箱养殖、高密度集约化池塘养殖等模式[2-4]。随着养殖产量、经济效益逐渐提高,出现无整体规划,养殖品种结构单一,水源交叉感染,水质恶化、密度较高致过度使用鱼药等问题,使罗非鱼养殖水质、环境、鱼病以及品质等方面陷入瓶颈[5]。王武等研究表明水体水质单因子变化对养殖动物有显著影响[6-7],李如忠等研究表明水质参数变化较大,且对养殖动物影响较大的主要是氮、磷等有机污染指标[8-10]。因此,我国水产科技工作者通过运用光合细菌、生物浮岛和混养等多种方法对南美白对虾、中国对虾、鲫鱼等生物的养殖水体的水质改善、促生长、抗病等方面做了部分研究[11-18],并取得了一定的成效,但有关不同养殖模式对精准养殖水体水质及鱼类生长性能的影响仍需进一步研究。
本研究以尼罗罗非鱼为研究对象,通过采用投放光合细菌、安装生物浮床单养和罗非鱼、鲫鱼、鳙鱼混养等3种不同模式对尼罗罗非鱼精准养殖水体水质和生长等方面进行研究,旨在探索尼罗罗非鱼生态高效健康养殖模式,以期为推动我国罗非鱼养殖模式由资源高耗费型向资源节约与环境友好型转变提供新的途径和思路。
1 材料与方法
1.1 试验材料
1.1.1 试验用饲料
试验所用饲料为浮水料,由中国水产科学研究院珠江水产饲料厂提供。其饲料配方为:鱼粉1(国产) 9.00%,消化能11.7 MJ/kg,大豆粕15.56%,粗蛋白33.28%,棉仁粕25.00%,粗脂肪2.81%,菜籽粕 20.00%,钙1.26%,次粉19.86%,总磷1.14%,小麦麸6.72%,有效磷0.45%,磷酸二氢钙1.06%,赖氨酸1.50%,植物油0.50%,蛋氨酸0.54%,盐0.20%,蛋氨酸+胱氨酸1.13%,沸石粉1.00%,苏氨酸1.29%,甜菜碱0.10%,色氨酸0.45%,预混料1.00%,精氨酸2.33%。每次投喂后密封保存。
1.1.2 试验用鱼及相关材料
试验用尼罗罗非鱼,规格为(10.8±1.2) g,体质健壮,外观无明显症状,购自广东佛山市三水白金种苗有限公司。光合细菌购自中国水产科学研究院珠江水产研究所药物实验厂,试验开始前7 d开始培养,浓度为1~2 g/m3。生物浮床为由PVC管框架固定的聚乙烯网,其上放空心菜,且长×宽为50 cm×6 cm的生态基垂于聚乙烯网下方。
1.1.3 试验时间及地点
试验于2014年6月5日至8月15日,在中国水产科学研究院(广州)农业部热带亚热带水产资源利用与养殖重点实验室试验基地水泥池内进行,水泥池规格长×宽×高为5.0 m×4.0 m×1.5 m,水深1.2 m。
1.2 试验方法
1.2.1 试验分组
试验分光合细菌组(定义为组1),生物浮床组(定义为组2),罗非鱼、鲫鱼和鳙鱼混养组(定义为组3)和对照组,罗非鱼每组各50尾,混养组的鲫鱼和鳙鱼各10尾,每组3个重复。
1.2.2 管理方法与计算公式
试验用水为经过2级沙滤养殖池塘水,采用3 kW的鼓风机通过微孔曝气盘(气盘直径为1 m)24 h不间断向鱼池充氧,温度为自然水温,pH值为 6.8~7.3,溶解氧量在5.0 mg/L 以上;每天09:00和17:00各投喂1次,投喂量为鱼质量的3%。根据以下公式记录并计算相应的数据:
平均体质量(g)=总体质量/罗非鱼尾数;
增加的均体质量(g)=试验后均体质量-试验前均体质量;
成活率=最后存活尾数/初始养殖尾数×100%;
增重率=(试验后均体质量-试验前均体质量)/初始均体质量×100%。
1.2.3 水质检测
每隔10 d采1次水样,每次取同一水泥池不同的3个重复点,分别检测氨氮、活性磷酸盐、硝酸盐氮、亚硝酸盐氮等4个指标。其中,活性磷酸盐浓度测定采用磷钼蓝分光光度法(HJ 593—2010),氨氮浓度测定采用纳氏试剂分光光度法(HJ 535—2009),NO2-浓度测定采用分光光度法(GB 7493—1987),NO3-浓度测定采用紫外分光光度法(HJ/T 346—2007)。
1.2.4 数据处理
采用Excel和SPSS 17.0进行统计分析,数据以“平均值±标准差(x±s)”表示,在单因子方差分析(ANOVA)基础上采用t检验分析比较,取95%置信度(P<0.05)或99%置信度(P<0.01)。
2 结果与分析
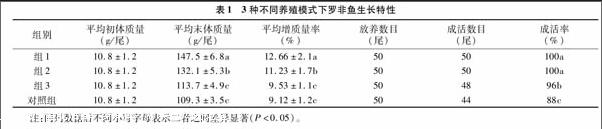
2.1 3种不同养殖模式下罗非鱼生长特性
由表1可知,光合细菌组、生物浮床组、混养组合和对照组的平均增质量率分别为(12.66±2.1)、(11.23±1.7)、(9.53±1.1)、(9.12±1.2) g,光合细菌组平均增长率最大,生物浮床组次之,对照组最低。除混养组合和对照组间的平均增质量率无显著差异外(P>0.05),其余组间均有显著差异(P<0.05)。成活率除光合细菌组和生物浮床组间无显著差异外(P>0.05),其余组间均存在显著差异(P<0.05)。可见,光合细菌和生物浮床对罗非鱼的生长和成活率有显著提高(P<0.05)。
2.2 3种不同养殖模式对精养水体水质的影响
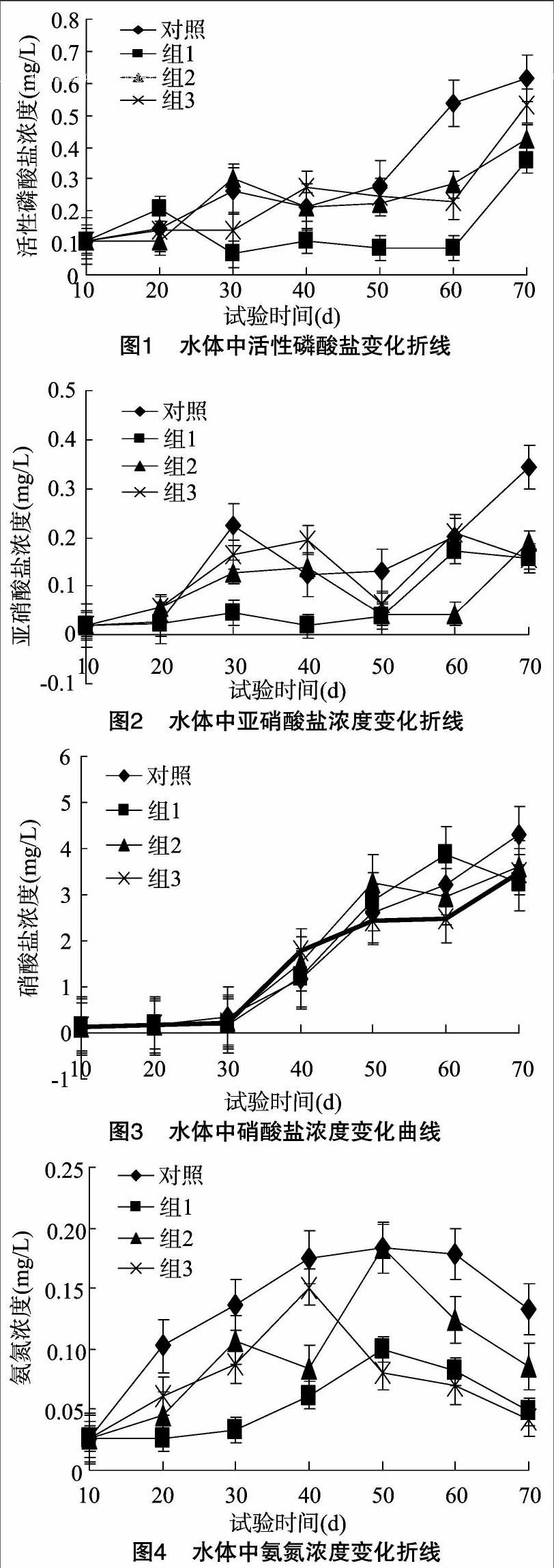
2.2.1 3种不同养殖模式对精养水体中活性磷酸盐浓度的影响
由图1可知,3种养殖模式对试验周期内罗非鱼精养水体中磷酸盐浓度均具有不同程度的降低作用。除前期10 d内的变化无显著差异外(P>0.05),10 d之后光合细菌组的作用尤其显著(P<0.05),其后依次是生物浮床组、混养组和对照组。30 d和40 d的生物浮床组和混养组相比其他组对磷酸盐浓度的降解作用分别呈现最弱现象。
2.2.2 3种不同养殖模式对水体中亚硝酸盐浓度的影响
由图2可知,试验进行20 d周期内,3种养殖模式对水体中的亚硝酸盐浓度的变化无显著影响(P>0.05);随着试验的进行,20~50 d试验周期内,光合细菌组对水体中亚硝酸盐的去除效果最为显著(P<0.05),其次依次是生物浮床、混养模式和对照组。50 d之后,生物浮床组对水体中的亚硝酸盐浓度的去除效果较为显著(P<0.05)。
2.2.3 3种不同养殖模式对水体中硝酸盐的影响
由图3可见,3种不同养殖模式对硝酸盐浓度影响无显著差异(P>0.05)。随着养殖周期的延长,硝酸盐的浓度均呈急剧上升的趋势。50 d之后的养殖周期内,光合细菌组的硝酸盐浓度升高较为显著(P<0.05),混养组的硝酸盐浓度相对较低,生物浮床组与对照组居中。
2.2.4 3种不同养殖模式对水体中氨氮浓度的影响
由图4可知,3组水体中的氨氮含量在养殖周期内均呈现先升高后降低的规律。其中,光合细菌组和生物浮床组水体中的氨氮含量大致在50 d之后呈下降趋势,而混养组水体中的氨氮含量在养殖周期40 d后呈现快速降低,整个养殖周期内,光合细菌组水体中的氨氮含量均显著低于其他组(P<0.05),对照组水体中的氨氮含量始终高于其他组(P<0.05)。
3 讨论
3.1 3种不同养殖模式均可不同程度的提高罗非鱼生长速度和成活率
氨氮与亚硝酸盐浓度的高低是制约水产养殖水质好坏的重要因素[19],而水质又进一步制约着水生生物的生长特性[20]。光合细菌是一种以光为能源的微生物,通过吸收水域中的有机物、二氧化碳、硫化氢等有害物质,能达到改善水质、增强水生生物免疫以促进其生长的作用,其作用已在陆家昌等开展的光合细菌能显著提高凡纳滨对虾生长速度和抗病能力研究[15]中得以证实。生态基作为载体材料可分为有机部分和无机部分,都可以应用于改善养殖水质[21],并能显著促进罗氏沼虾的生长[22]。本试验中,相对于对照组,光合细菌组和下方挂有生态基的生物浮床组对罗非鱼平均增长率最大和成活率具有显著促进作用(P<0.05)(表1),此结果与陆家昌等的研究结果[15,22]基本一致。混养组无显著变化,推测此结果可能与环境容纳量有关,其具体原因有待进一步研究,但在不增加饲料投喂量的情况下可额外获得鱼产量。可见,3种养殖模式均可不同程度的提高罗非鱼的生长与成活率。
3.2 3种不同养殖模式可有效改善罗非鱼精准养殖水体水质
由图1至图4可知,前10 d的养殖周期内,因生物个体小、投饵少及投饵带来的残饵、粪便相对较少,3种养殖模式对水体的磷酸盐、硝酸盐、亚硝酸盐和氨氮的影响无显著差异(P>0.05),此结果与沈锦玉等的研究结果[11]基本一致。Green等研究发现罗非鱼精养池塘对投入物质的氮、磷利用率分别仅为18%~21%和16%~18%[23]。杨世平等研究发现,投饵输入池塘的氮大部分(62%~68%)随着养殖时间的延长、精养池塘中亚础酸盐氮、氨态氮和总磷含量会逐渐增加[24]。
图1试验结果表明,养殖周期30 d时,随着投饵量的增加,光合细菌和混养对水中活性磷酸盐含量的降解作用显著优于其余2组(P<0.05),其原因可能为混养组和光合细菌的数量此时达到生态平衡,混养组的鲫鱼和鳙鱼吸食有机碎屑,减缓了水体中磷酸盐浓度升高的速度,而生物浮床的水生植物吸收相對速率较慢,同时附着在生态基上的生物群落生产繁殖速率低于水体活性磷酸盐的富集速率。图2试验结果表明,20~50 d试验周期内,光合细菌组对水体中亚硝酸盐的去除效果最为显著(P<0.05),其次依次是生物浮床、混养模式和对照组。生物50 d之后,生物浮床组对水体中的亚硝酸盐浓度的去除效果较为显著(P<0.05)。可见,在养殖前期,光合细菌因其稳定的增殖和亚硝酸盐的分解作用优势显著,这与沈锦玉等关于光合细菌水产养殖水质的改良结论[11]基本一致。Azim研究表明,微生物和浮游生物在生物膜上的形成受水质环境因子的影响较大,相对稳定的水体其形成时间约在30 d [25],与本试验50 d后生物浮床对亚硝酸盐去除效果显著的结论相符。
在池塘养殖过程中,氮的消失通常是通过氨的散发和反硝化作用[26],而反硝化作用过程常伴随着氧化的氮化合物扩散[27]。伴随着水体亚硝酸盐的分解转化为硝酸盐和水体内硝酸盐的积累,致使光合细菌组硝酸盐的浓度显著高于其他组(图3)。通常,水体中的氨氮在细菌作用下,转化为亚硝酸盐和硝酸盐,并最终转化为硝酸盐。因此,光合细菌组与生物浮床组对水体氨氮的分解转化效果较为显著(图4),此结论与沈锦玉等关于光合细菌对养殖水质氨氮的影响研究结果[11,15]基本相同。
水泥池或池塘养殖因其封闭性导致池塘水体生态因子之间的相互转化率较流水有显著降低[28],养殖水体维持一定的C/N比可显著提高池塘的稳定性[29],光合细菌、生物浮床和混养养殖模式因转化分解水体的氨氮、亚硝酸盐和硝酸盐等途径不同程度的降低了水体总氮含量,间接提高了C/N比值和物质转化,提高了养殖水体的生态平衡和水体的稳态,进而降低罗非鱼应激发生率和发病率,促进罗非鱼快速生长,可见,光合细菌、生物浮床和混养养殖模式均不失为罗非鱼精准水体健康高效养殖模式。
参考文献:
[1]陈胜军,李来好,杨贤庆,等. 我国罗非鱼产业现状分析及提高罗非鱼出口竞争力的措施[J]. 南方水产,2007,3(1):75-80.
[2]周 玲. 两种罗非精养模式的氮,磷收支研究[D]. 湛江:广东海洋大学,2010:1-9.
[3]李思发. 中国大陆罗非鱼养殖业发展对策[J]. 中国渔业经济研究,1999,7:13-15.
[4]夏德全.中国罗非鱼养殖现状及发展前景[J]. 科学养鱼,2000(5):4-21.
[5]郭鸿鹏,罗良国,任爱胜,等. 我国节水农业发展面临的问题和对策[J]. 中国软科学,2000(7):12-14.
[6]王 武. 精养鱼池耗氧和氧债的研究[J]. 上海水产大学学报,1984,12(2):99-102.
[7]雷衍之. 无锡市河浮口高产鱼塘水质的研究[J]. 水产学报,1983,7(3):185-198.
[8]李如忠. 水质评价理论模式研究进展及趋势分析[J]. 合肥工业大学学报:自然科学版,2005,28(4):369-373.
[9]黄海珠. 池塘水产养殖污染因子成因初探[D]. 南昌:南昌大学,2008.
[10]刘焕亮,黄樟翰. 中国水产养殖学[M]. 北京:科学出版社,2008.
[11]沈锦玉,尹文林,刘 问,等. 光合细菌HZPSB对水产养殖水质的改良和对鱼类促生长作用[J]. 科技通报,2004,20(6):481-484.
[12]张毅敏,高月香,吴小敏,等. 复合立体生物浮床技术对微污染水体氮磷的去除效果[J]. 生态与农村环境学报,2010,26(z1):24-29.
[13]周小平,王建国,薛利红,等. 浮床植物系统对富营养化水体中氮、磷净化特征的初步研究[J]. 应用生态学报,2005,16(11):2199-2203.
[14]全为民,沈新强,严力蛟. 富营养化水体生物净化效应的研究进展[J]. 应用生态学报,2003,14(11):2057-2061.
[15]陆家昌,黄翔鹄,李 活,等. 光合细菌对养殖水质及凡纳滨对虾抗病力的影响[J]. 广东海洋大学学报,2009,29(6):87-91.
[16]王吉桥,李德尚,董双林,等. 鲈-中国对虾-罗非鱼混养的实验研究[J]. 中国水产科学,2000,7(4):37-41.
[17]田相利,王吉桥,李德尚,等. 海水池塘中国对虾与尼罗罗非鱼施肥混养的实验研究[J]. 应用生态学报,1997,8(6):628-632.
[18]王吉桥,李德尚,董双林,等. 中国对虾与海湾扇贝投饵混养的实验研究(英)[J]. 中国水产科学,1999,6(1):97-102.
[19]刘艳红,罗国芝,朱学宝. 海水闭合循环系统生物滤器微生物特性研究[J]. 农业环境科学学报,2004,23(3):540-544.
[20]Pillay T V R. Waste production in aquaculture[M]. Noida:Blackwell Publishing,2004:58-75.
[21]Weerasooriya A C,Yi Y. Effects of AquaMats on growth of Nile tilapia (Oreochromis niloticus) fry in Earthen Ponds at AIT,Thailand[J]. Tropical Agricultural Research,2002,14:72-79.
[22]Haque M R,Islam M A,Rahman M,et al. Effects of C/N ratio and periphyton substrates on pond ecology and production performance in giant freshwater prawn Macrobrachium rosenbergii (de Man,1879) and tilapia Oreochromis niloticus (Linnaeus,1758) polyculture system[J]. Aquaculture Research,2015,46(5):1139-1155.
[23]Green B W,Boyd C E. Chemical budgets for oranically fertilized fish ponds in the drytropics[J]. Journal of the World Aquaculture Society,1995,26(3):284-296.
[24]杨世平,邱德全. 对虾高密度养殖过程中水质的周期变化与分析[J]. 水产科学,2006,25(9):459-462.
[25]Azim M E,Wahab M A. Periphyton-based pond polyculture[M]∥Azim M E,Verdegem M C J,van Dam A A,et al. Periphyton:ecology,exploitation and management. UK:CABI Publishing,2005:207-222.
[26]Schwartz S H. Are there universal aspects in the structure and contents of human values[J]. Journal of Social Issues,1994,50(4):19-45.
[27]吳丽红,李晓惠,杨 芳,等. 1株用于生物强化的高效反硝化菌的筛选鉴定[J]. 江苏农业科学,2014,42(12):371-373.
[28]晏小霞,唐文浩. 养殖水环境生物修复研究进展[J]. 热带农业科学,2004,24(2):69-72.
[29]Avnimelech Y. Carbon/nitrogen ratio as a control element in aquaculture systems[J]. Aquaculture,1999,176(3/4):227-235.
猜你喜欢
环境(2023年5期)2023-06-30
当代水产(2019年1期)2019-05-16
当代水产(2019年3期)2019-05-14
电子制作(2018年14期)2018-08-21
河北渔业(2016年10期)2016-11-22
水利规划与设计(2016年7期)2016-02-28
河北渔业(2015年2期)2015-03-26
河南科技(2014年23期)2014-02-27
