
君子兰自交不亲和性及自交不亲和机理
2017-03-21 00:00高巍马艳丽尹立辉
江苏农业科学 2016年11期
关键词:君子兰
高巍+马艳丽+尹立辉

摘要:以大花君子兰为材料,研究君子兰的自交不亲和性及作用机理。采用套袋自交法测定君子兰自交亲和指数;采用荧光显微镜观察法研究自交不亲和机理。结果表明:君子兰同花自交亲和指数0.29,同株自交亲和指数 3.43,种内自交亲和指数10.15。由此总结出君子兰同花自交和同株自交具有自交不亲和性,胼胝质主要出现在花粉管中,属配子体不亲和性。
关键词:君子兰;亲和指数;胼胝质;自交不亲和性
中图分类号: S334.2+3;S682.1+30.3 文献标志码: A
文章编号:1002-1302(2016)11-0207-02
自交不亲和性(SI)指某一植物的雌雄两性机能正常,但不能进行自花受精或同一品系内异株花粉受精的现象[1-2]。自交不亲和性在高等植物中普遍存在[3],是一种有利于异花授粉的生殖隔离[4]。
君子兰(Clivia miniata)为石蒜科(Amaryllidaceae)君子兰属(Clivia Lindl.)多年生草本植物。君子兰全株可入药,并且具有较高的观赏价值[5]。研究君子兰自交不亲和性对其生殖生物学研究和杂种优势利用意义重大。
1 材料与方法
1.1 材料
供试材料为大花君子兰胜利杂交品种,栽培于吉林农业大学药用植物苗圃温室中,正常栽培管理。田间试验在吉林农业大学温室中进行,室内试验在吉林农业大学中药材学院育种实验室进行。
1.2 方法
1.2.1 亲和指数测定 选取9株健壮君子兰植株,分为3组。从含蕾期后期开始,用硫酸纸袋将君子兰花朵单独套袋。待花朵开放时,于09:00—10:00轻轻打开硫酸纸袋,对其进行授粉,授粉2 d后去掉硫酸纸袋,观察坐果情况。A组为同花自交授粉、B组为同株异花授粉、C组为异株授粉,试验重复3次。相关公式:
坐果率=坐果花朵数/授粉花朵数×100%;
亲和指数=结籽数/自交花数。
1.2.2 自交不亲和机制研究 于每天10:00取样,分别取不同自交组合授粉后4 h的君子兰雌蕊、花柱连同子房固定于乙醇(FAA)固定溶液福尔马林-乙醇中。每个处理取3朵花,重复3次。
观察前将柱头从固定液中取出,依次通过70%、50%、10%乙醇梯度复水,然后置于1 mol/L氢氧化钠(NaOH)溶液中,于60 ℃水浴软化90 min,蒸馏水冲洗3次,再用 0.1 mol/L 磷酸钾(K3PO4)配制的0.1%苯胺蓝溶液染色 24 h,用100%甘油压片,在OlympusIX71倒置型荧光显微镜下观察柱头上的花粉萌发及花粉管生长情况,并拍照[6-7]。
1.3 数据处理及分析
研究数据均为试验样品的平均值,应用DPS软件对参数的平均值进行方差分析,以评价本试验中处理的差异显著性。
2 結果与分析
2.1 君子兰自交亲和指数
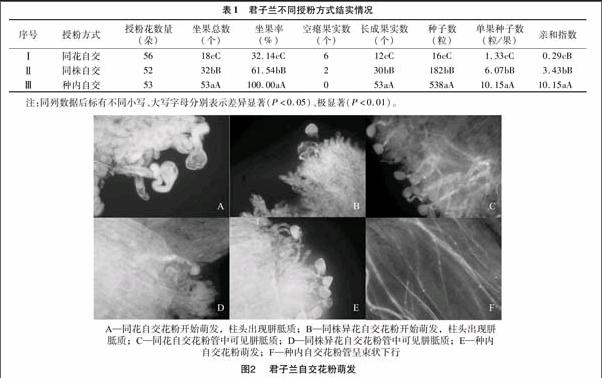
植物在种子繁殖时果实虽然能够膨大,可以坐果,但不一定生成可育种子(图1)。君子兰授粉花数量和结种数见表1,可见同花自交授粉坐果18个,包括空瘪果实6个。君子兰空瘪果颜色同样变为红色,内部可见败育胚珠(图1-D)。同花自交授粉法长成果实12个,共结种子16粒,每果结种子1~2粒,亲和指数为0.29。同株异花授粉亲和指数有3.43,授粉花数52朵,坐果32个,空瘪果实2个,结种数182粒,单果结种子6粒左右。同品种(系)授粉结实率100%,单果包含种子10~11粒,亲和指数达到10.15(表1)。王晓佳等采用亲和指数判定菘蓝自交不亲和性,以0<自交亲和指数<0.5作为强自交不亲和判定依据[8]。本研究中同株同花自交、同株异花授粉亲和指数显著低于异株授粉,可见君子兰同花授粉和同株授粉具有自交不亲和性。
2.2 君子兰自交不亲和性机制
用荧光显微技术观察柱头荧光现象已成为细胞生物学的一个非常重要而有效的研究手段。萌发的花粉管进入柱头后从花柱穿过,最终到达胚囊释放精细胞,精、卵细胞相互识别并且融合,完成受精过程[9]。自交不亲和表现为这一过程中的某个环节出现抑制障碍,受精过程不能顺利完成。
君子兰花粉黏住柱头,花粉开始萌发,柱头蛋白质与花粉蛋白质相互识别,2 h后花粉萌发[10]。同株同花授粉4 h(图2-A)花粉管开始萌发,柱头上出现胼胝质,导致部分花粉管在柱头上盘绕,不能进入柱头中;同株异花授粉4h柱头上出现胼胝质(图2-B),花粉管中出现不均匀胼胝质(图2-C、图2-D),使花粉管向花丝底部生长受阻;异株授粉4 h花粉在柱头顺利萌发,花粉管呈束状在花柱道内下行(图2-E、图2-F)。
王冲等研究君子兰中间杂交时可见胼胝质出现在柱头乳突细胞和花柱通道[11]。李守丽等研究百合远缘杂交时,在柱头乳突细胞和花柱通道发现胼胝质,可见胼胝质是阻碍植物受精的屏障[12-13]。已有研究表明,配子体不亲和,花粉管在花柱中被抑制[14-16],本研究中君子兰花粉管被抑制的部位在雌蕊的花柱,胼胝质大量出现在花柱中,柱头上也有少数可见。
3 结论
君子兰同花自交授粉、同株自交授粉过程中,胼胝质大量出现在花粉管中,花粉管在花柱中生长中途停顿;同时也可见少量胼胝质出现在柱头表面,导致花粉管缠绕和不能正常发育的现象。君子兰自花不亲和现象发生在同花自交和同株自交上,属配子型自交不亲和。已有研究表明,S等位基因控制植物自交不亲和性,关于君子兰自交不亲和性的基因研究有待进一步开展。
参考文献:
[1]East E M.The distribution of self-sterility in flowering plants[J]. Proc Am Phil Soc,1940,82:449-518.
[2]Lewis D.Sexual incompatibility in plants[M]. London,UK:Edward Amold Limited,1979.
[3]Brewbaker J L.Biology of the angiosperm pollen grain[J]. Indian J Genet Plant Breed,1959,19:121-133.
[4]Boves D C,Nasrallah J B. Physical linkage of the SLG and SRK genes at the sel-incompatibility locus of Brassica oteracea[J]. Mol Gen Genet,1993,236(2/3):267-269.
[5]姜澤慧. 中国君子兰[M]. 北京:中国林业出版社,2003:128-149.
[6]Burke J J. Moisture sensitivity of cotton pollen[J]. Agronomy Journal,2002(94):883-886.
[7]Hirasuka S,Zhang S L,Nakagawa E,et al.Selective inhibition of the growth of incompatible pollen tubes by S-protein in the Japanese pear[J]. Sexual Plant Reproduction,2001,13(4):209-215.
[8]王晓佳,朱利泉. 甘蓝自交不亲和性的测定方法[J]. 农业生物技术学报,1998,6(2):195-198.
[9]胡适宜. 被子植物生殖生物学[M]. 北京:高等教育出版社,2005.
[10]高 巍,任跃英,杨义波,等. 君子兰花粉萌发条件研究及生活力测定[J]. 安徽农业科学,2011,39(36):22269-22270.
[11]王 冲,雷家军,姜 闯,等. 君子兰种间杂交亲和性[J]. 中国农业科学,2011,44(18):3822-3829.
[12]李守丽,石 雷,张金政. 大百合与百合属间授粉后花粉管生长发育的观察[J]. 园艺学报,2006,33(6):1259-1262.
[13]王文和,王树栋,赵祥云,等. 百合远缘杂交互粉萌发及花粉管生长过程观察[J]. 西北植物学报,2007,27(9):1790-1794.
[14]高新起,王秀玲. 植物的自交不亲和性[J]. 曲阜师范大学学报,1999,25(2):84-86.
[15]Heslop-Harrison J,Knox R B,Heslop-Harrison Y. Pollen-wall proteins:exine-held fractions associated with the incompatibility responses in cruciferae[J]. Theoretical and Applied Genetics,1974,43(3):133-137.
[16]Edlund A F,Swanson R,Preuss D. Pollen and stigma structure and function:the role of diversity in pollenation[J]. Plan Cell,2004,16:84-97.
猜你喜欢
作文成功之路·小学版(2021年2期)2022-01-01
作文成功之路·教育教学研究(2021年2期)2021-04-07
杂文选刊(2014年6期)2014-06-06
杂文选刊(2014年6期)2014-06-06
