
银杏叶提取物对睡眠剥夺小鼠学习记忆能力的影响及其机制的研究
2016-04-07 20:27唐梦晨陈娇徐文杰李凌
中国当代医药 2016年6期
唐梦晨 陈娇 徐文杰 李凌

[摘要] 目的 研究银杏叶提取物(EGB)对睡眠剥夺小鼠学习记忆能力的影响及其机制。 方法 选取成年健康清洁级小鼠48只,雌雄各半,根据随机数字表法将小鼠随机分为4组(n=12):空白组,对照组,治疗Ⅰ组,治疗Ⅱ组,适应性饲养3天,每组筛选出9只进行造模。应用72 h睡眠剥夺法制作动物模型,用Y迷宫成绩(正确反应次数)评定学习记忆能力,免疫组织化学法反映小鼠海马脑源性神经营养因子(BDNF)的蛋白表达。 结果 Y迷宫测试结果显示,给药后,对照组与治疗Ⅰ组、对照组与治疗Ⅱ组的正确反应次数差异有统计学意义(P<0.05);对照组给药前后的正确反应次数差异有统计学意义(P<0.05),其余3组差异无统计学意义(P>0.05)。与对照组比较,空白组、治疗Ⅰ组、治疗Ⅱ组海马区的BDNF表达增高,其灰度值差异有统计学意义(P<0.05),但空白组、治疗Ⅰ组、治疗Ⅱ组各组间的灰度值差异无统计学意义(P>0.05)。 结论 睡眠剥夺能造成脑损伤、学习记忆能力减退以及BDNF表达减少,EGB可能是通过上调BDNF蛋白及其相关通路蛋白改善睡眠剥小鼠的学习记忆能力。
[关键词] 银杏叶提取物;睡眠剥夺;脑源性神经营养因子;学习记忆
[中图分类号] R332 [文献标识码] A [文章编号] 1674-4721(2016)02(c)-0004-04
[Abstract] Objective To study the influence extract of Ginkgo biloba (EGB) on learning and memory ability in sleep-deprivation mice and its mechanism. Methods Forty-eight healthy and clean adult mice were selected,half male and half female,and they were evenly divided into 4 groups according to the random number table method:blank group,control group,treatment groupⅠ, and treatment group Ⅱ,and they were given adaptive feeding for 3 days,9 mice were selected in each group to make model.72-hour of sleep-deprivation method model was applied.The learning and memory ability was evaluated by achievement of Y maze.The protein expression of brain-derived neurotrophic factor (BDNF) in hippocampal area in mice was reflected by immunohistochemistry. Results Test result of Y maze showed that,after administration,there was a statistical difference of right reaction times between control group and treatment groupⅠ,and between control group and treatment groupⅡ (P<0.05);there was a statistical difference of right reaction times before and after administration in control group (P<0.05),the other three groups had no statistical difference (P>0.05).Compared with control group,BDNF expression of hippocampal area in blank group,treatment group Ⅰ,and treatment group Ⅱ was improved,and there was a statistical difference of gray value (P<0.05),however,there was no statistical difference of gray value among all groups (blank group,treatment group Ⅰ,and treatment group Ⅱ) (P>0.05). Conclusion Sleep-derivation can lead to cerebral damage,decreased learning and memory ability,and reduction of BDNF expression,and EGB may improve learning and memory in sleep-deprivation mice by up-regulating BDNF protein and its related pathway protein.
[Key words] Extract of Ginkgo biloba;Sleep-deprivation;Brain-derived neurotrophic factor;Learning and memory
睡眠剥夺(sleep deprivation,SD)一般指在24 h中的睡眠<4 h,可以引起生理,心理甚至行为的变化[1]。机体处于正常睡眠状态下,脑内某些蛋白增加,对新突触的建立及学习记忆有很大的促进作用[2]。有研究[3]表明,中长期SD大鼠前额叶皮质脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)免疫阳性物质表达显著减少,可造成脑损伤,影响学习记忆能力。研究[4]表明,BDNF直接参与学习的可塑性,是管理和存储记忆的动力学因素,可能是调控学习和记忆的一个重要分子标志物。在成年中枢系统,BDNF的表达受很多因素的调节,同时通过调节突触的可塑性影响学习记忆[5]。大量研究[6]表明,在正常脑中生理剂量的BDNF对学习记忆有促进作用。
银杏叶提取物(extract of Ginkgo biloba,EGB)已被国内外用于改善脑功能不全患者的学习记忆能力, 是一种 “认知增强剂”[7]。 据报道[8],EGB具有提高内源性BDNF表达水平的作用,作用机制有待进一步研究。那么,EGB能否改善SD小鼠的学习记忆能力,目前尚无报道。
本研究拟采用行为学、免疫组织化学法等,一方面研究EGB对SD小鼠学习记忆能力的影响,另一方面研究EGB对SD小鼠海马BDNF的影响,进而揭示其可能的作用机制,为寻找能改善SD对学习记忆能力影响的方法提供依据。
1 材料与方法
1.1 实验动物
选取成年健康清洁级小鼠48只,雌雄各半,体质量为(23±2)g,由徐州医学院动物中心提供。饲养于相对湿度在30%~40%的动物房,光照节律为12L∶12D(8:00~20:00),温度为(25±1)℃。根据随机数字表法将小鼠随机分为4组,每组12只,分别为空白组,对照组,治疗Ⅰ组,治疗Ⅱ组。适应性饲养3 d,期间进行站台训练,每天训练30 min,可自由饮水及进食,避免噪声。每组筛选出9只训练合格的小鼠进行造模。
1.2 迷宫训练
实验时先将小鼠放入迷宫适应3 min,灯亮时该臂有电,小鼠受电击后逃到安全区,此区内无灯光。灯光持续10 s后,随机变化电击区,进行下一次测试。每个实验日连续测试20次,记录正确反应次数,所用参数:电压为50~70 V,延时5 s。各组造模前3天进行每天20次的训练,取第3天成绩作为造模前成绩(正确反应次数);给药后再进行测试。
1.3 动物模型制备
应用改良多平台睡眠剥夺法(modified multiple platform method,MMPM)(72 h睡眠剥夺法)制作动物模型:各组从造模第1天,8:00 开始放入睡眠剥夺箱,连续3 d,至第4天早8:00结束。用大鼠笼[规格:48 cm(长)×40 cm(宽)×34 cm(高)]自制简易睡眠剥夺箱,其底面均匀固定9个上表面积2.5 cm×2.5 cm的木平台。箱内注水,平台高出水面0.5 cm,水温保持在室温24~26℃。箱顶置铁丝网。每天更换箱中的水。
1.4 主要药品与试剂
银杏叶片(每片含总黄酮醇苷19.2 mg,萜类内酯4.8 rag,批号:20070403):贵州信邦制药股份有限公司;BDNF试剂盒:武汉博士德;SABC(兔IgG):武汉博士德;PBS液:北京中杉金桥;DAB显色剂:武汉博士德。
1.5 实验处理
空白组:正常睡眠,等量去离子水灌胃5 d;对照组:睡眠剥夺,等量去离子水胃5 d;治疗Ⅰ组:睡眠剥夺,EGB 9 ml/10 g灌胃5 d;治疗Ⅱ组:睡眠剥夺,EGB 18 ml/10 g灌胃5 d。给药后第6天开始再次分别对4组小鼠进行行为学检测。
1.6 免疫组织化学检测法
造模结束,经行为学检测,麻醉成功后,将小鼠仰卧固定于手术板上,剪开前胸壁肋骨,暴露心脏,小心分离出升主动脉并在其下穿1根1号手术线备用,将灌注针从心尖插入升主动脉,用准备好的手术线将灌注针固定于升主动脉,在左心耳剪1个小口,输液器接于灌注针尾部,止血钳夹闭腹主动脉,小鼠头低脚高位,快速灌注生理盐水直至流出液变清,再以4℃的4%多聚甲醛继续灌注,直至小鼠头颈变硬,快速断头取脑,取海马,连续冠状切片,PBS漂洗3次,3 min/次,去离子水冲洗;以3%过氧化氢溶液孵育20 min,PBS漂洗3次,3 min/次;正常羊血清封闭(3∶200,37℃,15 min);倾去血清,勿洗,滴加一抗(BDNF兔来源)(1∶60,4℃过夜),PBS漂洗3次,3 min/次;滴加二抗(1∶200,37℃),孵育10 min,PBS漂洗3次,3 min/次;滴加AB液(1∶100, 37℃,40 min),PBS漂洗3次,3 min/次;DAB显色(1∶20);最后进行梯度乙醇脱水,二甲苯透明,封片,晾干后进行镜下观察、拍照。蛋白表达相对量以灰度值表示。各组随机选取3只小鼠,每只小鼠随机选取3个部位,每个部位9个视野,应用ImageJ软件进行图像分析。
1.7 统计学处理
采用SPSS 13.0软件分析,Y迷宫成绩、海马内BDNF蛋白含量用均数±标准差(x±s)表示,组间比较采用单因素方差分析(One Way-ANOVA)检验,以P<0.05为差异有统计学意义。
2 结果
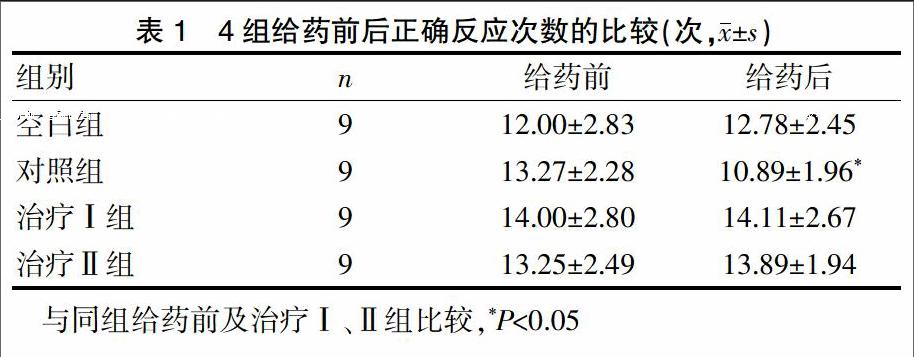
2.1 4组小鼠给药前后正确反应次数的比较
Y迷宫测试结果显示,对照组给药前后的正确反应次数差异有统计学意义(P<0.05),其余3组差异无统计学意义(P>0.05);给药后,对照组与治疗Ⅰ组、对照组与治疗Ⅱ组的正确反应次数差异有统计学意义(P<0.05)(表1)。
2.2 4组小鼠BDNF免疫组织化学染色结果的比较
免疫组织化学染色结果显示,BDNF主要表达于海马细胞的细胞质和细胞膜,呈棕黄色的为阳性。对照组与空白组比较,BDNF蛋白表达明显减少(图1A、图1B)。与对照组比较,空白组、治疗Ⅰ组、治疗Ⅱ组海马区的BDNF表达增高(图1A~图1D),其灰度值差异有统计学意义(P<0.05),但空白组、治疗Ⅰ组、治疗Ⅱ组各组间灰度值差异无统计学意义(P>0.05)(图1A、图1C、图1D)(表2)。
2.3 SD开始前后4组小鼠精神状况与活动能力的比较
SD开始前,4组小鼠饲养条件一致,精神状况与活动能力基本一致,体重持续增长。SD开始后,对照组、治疗Ⅰ组、治疗Ⅱ组精神萎靡、活动减少,且情况持续加重。SD 72 h,36只小鼠均存活。
3 讨论
本实验中,Y迷宫测试结果显示,对照组给药前后的正确反应次数差异有统计学意义,说明SD会造成小鼠学习记忆障碍[9]。给药后,对照组与治疗Ⅰ组、对照组与治疗Ⅱ组的正确反应次数差异有统计学意义,说明EGB对SD小鼠学习记忆有改善作用。
免疫组织化学染色结果显示,EGB可能促进海马BDNF的表达。小鼠海马BDNF的表达增高,对小鼠学习记忆有保护作用;EGB可改善SD小鼠学习记忆障碍,可能是通过上调海马BDNF表达引起的。由此可推测,EGB对改善SD患者的学习记忆障碍有一定作用[10]。目前的研究证实,一定剂量的EGB对认知功能具有改善作用[11-12]。
德国神经生物学家Barde等[13]于1982年首次从猪脑中发现BDNF,目前研究发现,BDNF广泛存在于脑内各个区域,以海马和皮质居多,与疼痛[14]、抑郁[15]、学习记忆[16]等相关,而越来越多的研究[17-19]证实,BDNF在学习记忆中占至关重要的地位,可能与长时程增强(LTP)有关。
本实验中还发现,SD小鼠虽精神萎靡,但攻击性增强,可能处于应激状态。有研究表明[20],长期应激可使下丘脑-垂体-肾上腺轴(HPA轴)高激活,进而导致体内糖皮质激素异常增高,与海马内存在的大量糖皮质激素受体选择性结合,从而导致海马结构异常。
综上所述,EGB可以保护SD小鼠海马区神经细胞,上调海马区BDNF的表达,提高小鼠的学习记忆能力,且EGB安全性好,无明显毒副作用,来源充足,价格低廉,因此,EGB可以作为改善SD学习记忆能力的药物,值得进一步临床研究。不同剂量EGB对改善SD小鼠学习记忆能力的效果也可作相关研究。
[参考文献]
[1] Venancio DP,Andersen LM,Vilamaior PS,et al.Sleep deprivation alters rat ventral prostate morphology 1eading to glandular atrophy:a microscopic study contrasted with the hormonal assays[J].J Biomed Biotechnol,2012,2012:285938.
[2] Maquet P.The role of sleep in learning and memory[J].Science,2001,294(5544):1048-1052.
[3] 冯婷,李峰,宋月晗,等.疏肝补肾法对睡眠剥夺大鼠学习记忆及前额叶皮质BDNF蛋白表达的影响[J].环球中医药,2012,5(4):255-259.
[4] Cunha C,Brambilla R,Thomas KL.A simple role for BDNF in learning and memory?[J].Front Mol Neurosci,2010,3:1.
[5] Musumeci G,Sciarretta C,Rodríguez-Moreno A,et al.TrkB modulates fear learning and amygdalar synaptic plasticity by specific docking sites[J].J Neurosci,2009,29(32):10131-10143.
[6] Zhen YF,Zhang J,Liu XY,et al.Low BDNF is associated with cognitive deficits in patients with type 2 diabetes[J].Psychopharmacology(Berl),2013,227(1):93-100.
[7] Wang Y,Wang L,Wu J,et al.The in vivo synaptic plasticity,mechanism of EGb 761-induced enhancement of spatial learning and memory in aged rats[J].Br J Phamaco1,2006, 148(2):147-153.
[8] 李育臣, 董玉娟, 王建茹,等.银杏叶提取物对脑缺血大鼠脑源性神经营养因子的影响[J].中风与神经疾病,2005,22(1):54-56.
[9] 赵忠新.临床睡眠障碍学[M].上海:第二军医大学出版社,2003.
[10] 袁宝强,耿海峰,李慧,等.银杏叶提取物上调海马脑源性营养因子和神经肽2受体表达、增强突出重建而改善发育期癫痫大鼠认知功能的研究[J].中华临床医师杂志,2012,6(11):2959-2965.
[11] Bachinskaya N,Hoerr R,Ihl R.Alleviating neuropsychiatric symptoms in dementia:the effects of Ginkgo biloba extract EGb 761.Findings from a randomized controlled trial[J].Neuropsychiatr Dis Treat,2011,7:209-215.
[12] Kaschel R.Specific memory effects of Ginkgo biloba extract EGb 761 in middle-aged healthy volunteers[J].Phytomedicine,2011,18(14):1202-1207.
[13] Barde YA,Edger D,Thoenen H.Purification of a new neurotrophic factor from mammalian brain[J].EMBO J,1982,1(5):549-553.
[14] 刘迪,唐倩倩,曹君利.脑源性生长因子参与疼痛-抑郁共病的研究进展[J].中国药理学通报,2015,31(1):26-30.
[15] 乔卉,安书成,徐畅.BDNF与抑郁症的研究现状及进展[J].生理科学进展,2011,42(3):195-200.
[16] Figurov A,Pozzo Miller LD,Olafsson P,et al.Regulation of synaptic responses to high frequency stimulation and LTP by neurotrophins in the hippocampus[J].Nature,1996, 381(6584):706-709.
[17] Barnes P,Thomas KL.Proteolysis of proBDNF is a key regulator in the formation of memory[J].PLoS One,2008,3(9):e3248.
[18] Pang PT,Lu B.Regulation of late-phase LTP and long-time memory in normal and aging hippocampus:role of secreted proteins tPA and BDNF[J].Ageing Res Rev,2004,3(4):407-430.
[19] Pang PT,Teng HK,Zaitsev E,et al.Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plastcity[J].Science,2004,306(5695):487-491.
[20] 徐辉,罗欢欢,朱金富.睡眠剥夺对抑郁样模型大鼠探索行为及海马神经元脑源性神经营养因子表达的影响[J].中华行为医学与脑科学杂志,2011,21(6):481-483.
(收稿日期:2015-09-18 本文编辑:许俊琴)
