
猪流行性腹泻病毒(XJ - DB2)株的分离鉴定
2016-03-31 07:43刘小兰陈兰玲杜久斌新疆天康畜牧生物技术股份有限公司新疆乌鲁木齐830011
中国兽医杂志 2016年1期
刘小兰,贺 笋,陈兰玲,杜久斌(新疆天康畜牧生物技术股份有限公司,新疆乌鲁木齐830011)
猪流行性腹泻病毒(XJ - DB2)株的分离鉴定
刘小兰,贺笋,陈兰玲,杜久斌
(新疆天康畜牧生物技术股份有限公司,新疆乌鲁木齐830011)
摘要:从新疆佃坝地区某猪场疑似流行性腹泻病毒(PEDV)感染的仔猪中采集肛拭子样品,将其接种到Vero细胞中,连续传代培养,并逐日观察细胞病变(CPE),分离得到一株PEDV病毒,命名为新疆佃坝2株(XJ-DB2)。将分离的XJ-DB2株进行ORF3基因序列分析比对和遗传进化分析,结果表明,XJ-DB2株为PEDV强毒株。其ORF3基因和我国分离的强毒株CHS株、CHGS株,韩国分离的强毒株处在同一个分支上,而目前广泛使用的疫苗毒株CV777、韩国分离的弱毒株、我国分离的弱毒株CHJS07则处在相对独立的分支上。XJ-DB2与CHS基因的核苷酸同源性最高为99.0%,与韩国分离的强毒株S-DR13同源性为98.8%,而与疫苗株CV777核苷酸同源性为97.3%。
关键词:猪流行性腹泻病毒;分离鉴定;ORF3基因;序列分析
自1971年首次在英国的猪群中发生猪流行性腹泻(Porcine epidemic diarrhea,PED)以来,直至1978年,英国和比利时才证实引起PED的病原为冠状病毒[1]。PED是由流行性腹泻病毒(PEDV)引起的以腹泻、呕吐、脱水和对哺乳仔猪高致死率为主要特征的一种高度接触性肠道传染病。在出生仔猪1~2周龄时,死亡率高达90%[2]。由于PED与猪传染性胃肠炎(TGE)等其他猪的消化道传染病近似,单靠临床症状和组织病理学检查来作出鉴别诊断是比较困难的,所以,PED的诊断要借助于血清学和病毒学的检验。但是,由于PEDV未能适应在传代细胞培养物中增殖,直到1988年,Hoffmann等首次在培养基中含胰酶的Vero细胞上成功繁殖了PEDV,并认为胰酶对PEDV纤突糖蛋白切割作用增强了病毒对Vero细胞的感染力,此方法在后续的PEDV分离、细胞培养和疫苗的研究中被广泛应用[3]。
PEDV为不分节段的线性单股正链RNA,其中ORF3位于sM蛋白和S蛋白之间,编码功能未知的多肽性片段,其编码分子量为25.3 ku[4-6]。Park等(2007)对PEDV强、弱毒的ORF3序列进行了分析,发现致弱的PEDV在ORF3有17个氨基酸的缺失,为强、弱毒鉴别诊断方法的建立提供了依据,推测这是和病原性密切相关的重要位点(Seong-JunPark,2007)[7]。已有的研究显示,ORF3与病毒毒力有关(Duarte M,et al,1994)[8]。
本研究针对新疆某猪场发生的流行性腹泻,采集腹泻仔猪肛拭子,采用特殊的病毒分离方法,分离到1株猪流行性腹泻病毒,通过分子生物学方法进行了鉴定和对ORF3基因进行了序列分析,充分评估该毒株的变异情况,从而为疫情防控提供了试验依据。
1 材料
1.1病料疑似流行性腹泻病毒(PEDV)感染的仔猪肛拭子。
1.2细胞非洲绿猴传代肾细胞(Vero)由本实验室保存。
1.3细胞培养基及血清细胞培养基MEM和胎牛血清,均购自Hyclone公司。
1.4质粒与菌种DH5α感受态细胞由本实验室制备、保存;pMD19-T载体,购自宝生物工程(大连)有限公司。
1.5主要试剂核酸提取试剂盒,购自宝生物工程(大连)有限公司;胶回收试剂盒,购自天根生物科技(北京)有限公司。
1.6引物参照GenBank中登录的PEDV毒株的ORF3基因序列,设计引物,由上海生工生物工程技术服务有限公司合成。
2 方法
2.1PEDV的分离及在Vero细胞中传代培养
取适量处理后的腹泻病料接种铺满单层的Vero细胞,感作过夜,次日,弃去毒液,用PBS冲洗3次,添加5 mL无血清MEM培养基(添加适量胰酶),置37℃,5%CO2的培养箱中静置培养,每日观察细胞状态,72 h收获病毒,反复冻融3次后置-80℃以下保存,备用。按以上相同的方法将病毒在Vero细胞上连续传代。
2.2引物的设计与合成参照GenBank中登录的PEDV毒株的ORF3基因序列,设计引物,引物由上海生工生物工程技术服务有限公司合成,片段跨幅为整个ORF3基因,长度大小约为833 bp,该引物的序列为上游引物F:5′-TCCTAGACTTCAAC CTTACG-3′;下游引物R:5′-GGTGACAAGTGAAG CACAGA-3′。
2.3ORF3开放阅读框的克隆与序列测定用所设计引物进行PCR扩增,扩增产物电泳,经胶回收试剂盒切胶回收,纯化后与T载体连接,转化感受态细胞DH5α,用PCR方法鉴定重组质粒,选择阳性重组质粒送测序公司测序。
2.4PEDV ORF3序列比对将测序回复的序列经DNAStar软件拼接后与我国分离的CHS株,CHGS株,韩国分离的S-DR13株,经典CV777弱毒株,韩国弱毒株Attenuated DR13株(A-DR13),CHJS07弱毒株进行遗传进化分析绘制其遗传进化关系树。
3 结果
3.1病毒的分离与传代XJ-DB2株经Vero细胞传至第6代,无典型细胞病变,传至7代时出现轻微细胞病变,至第8代,细胞变圆,空泡化,形成合胞体,进而细胞脱落死亡,出现典型的细胞病变,如图1所示。随着代次增加,细胞病变趋于稳定,病变时间基本维持不变。本毒株在Vero细胞传代过程中,接毒量占总体积的10%左右,病变时间基本维持在48 h,病毒的TCID50可达106.0/mL。

图1 正常细胞对照及病变细胞A:未接种PEDV病毒的Vero细胞;B:接种PEDV 48 h后病变的Vero细胞
3.2PEDV目的片段的RT-PCR鉴定用本实验室自己设计并合成的特异性引物进行RT-PCR扩增,结果如图2所示,扩增片段大小约833 bp。
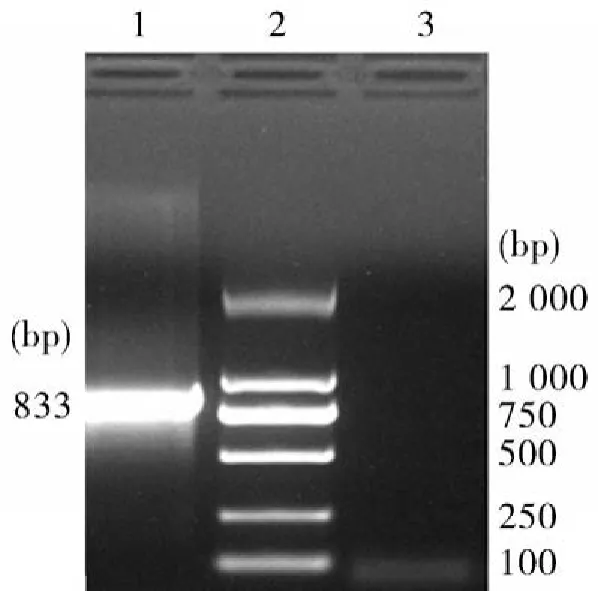
图2 目的片段的RT- PCR扩增注:1:目的片段;2:DNA Marker 2 000;3:PCR阴性对照
3.3ORF3遗传进化分析将本试验的分离株DB2与我国分离的CHS株,CHGS株,韩国分离的S-DR13株,经典CV777弱毒株,韩国致弱的A-DR13株,CHJS07弱毒株等的ORF3全基因序列应用DNAStar绘制遗传进化树(Clustal W Method)。从PEDV ORF3基因遗传进化可以看出,PEDV在遗传进化上呈2个明确的分支:强毒株和弱毒株(图3),强毒株又可分为2个亚群。从进化树上可以看出本次分离的强毒株与我国分离的CHS株和韩国分离的强毒株S-DR13处在同一分支上。XJ-DB2与CHS基因核苷酸同源性最高为99.0%,其次与韩国分离的强毒株S-DR13同源性为98.8%,而与疫苗株CV777核苷酸同源性为97.3%。

图3 XJ - DB2株ORF3基因遗传进化树
4 讨论
病毒分离的主要目的是通过对病毒的检测和纯化,获得纯化的病毒粒子,并将其进行传代培养。自20世纪70年代初发现PEDV以来,曾用各种方法试图把病毒适应于细胞培养,但是都没有成功,成为病毒学上的一道难题。直到1988年瑞典Hoffman等首次在Vero传代细胞的培养液中添加胰酶,病毒培养获得成功。本研究参考国内外文献,将胰酶浓度控制在3~8 μg/mL,在Vero细胞中连续传代而获得XJ-DB2毒株。虽然Vero细胞是目前用于分离培养PEDV的最佳细胞系,但是Vero细胞不同来源差异、毒株间的差异及培养条件均是影响成功分离PEDV的重要因素。
ORF3是PEDV基因组中惟一的附属基因,PEDV野毒株在ORF3基因上存在差异[9]。因此,开展以ORF3基因为基础的PEDV序列分析具有重要意义。本研究中分离得到的PEDV新疆株ORF3基因序列分析结果显示,其ORF3基因没有碱基缺失或插入,大小为675 bp,编码224个氨基酸蛋白。该毒株与我国分离的强毒株CHS株,CHGS株,韩国分离的强毒株S-DR13株,亲缘关系较近,与CHS基因的核苷酸同源性最高为99.0%,与韩国分离的强毒株S-DR13同源性为98.8%,而与疫苗株CV777核苷酸同源性为97.3%。表明新疆近年来流行病毒株基因发生变异。
在过去的30年间,流行性腹泻病毒在亚洲猪场中的感染非常严重,2010年以来,我国暴发流行了仔猪腹泻,在我国PEDV感染给养猪业造成了巨大的经济损失。目前,流行性腹泻病毒季节性变得不明显,很多地方出现常年发病的情况,所以,现在很多猪场已开始不分季节,全年常态化免疫母猪。而PEDV变异株在细胞上的成功培养对其研究具有重要意义,为研究PEDV生化特性、疫苗制备及其相关分子生物学的研究奠定了基础。
参考文献:
[1]王宗仪.国外兽医学:畜禽传染病[M].北京:科学出版社,1989:4-6.
[2] Pensaert M B,Debouck P.A new coronavirus-like particle associated with diarrhea in swine[J].Arch Virol,1978,58(3):243-247.
[3] Hoffmann M,Wyler R . Propagation of the virus of Procine Epidemic Diarrhea in cell culture[J] . J Clin Microbiol,1988,11:2235-2239.
[4]张强敏,郭福生,尹燕博,等.猪流行性腹泻病毒分子生物学特征[J].中国病毒学,2002,17(4):381-384.
[5]吕茂杰,陈建飞,时红艳,等.猪流行性腹泻病毒核衣壳蛋白与感染细胞核磷蛋白的共定位分析[J].微生物学报,2011,51(5):643-647.
[6] Park S,Moon H J,Lou Y,et al . Cloning and further sequence analysis of the ORF3 gene of wild-and attenuated-type Porcine epidemic diarrhea viruses[J].Virus Genes,2008,36(1):95-104.
[7] Seong J,Hyoung J,Jeong S,et al.Sequenee analysis of the Partials Pike glyeo Protein gene of Poreine epidemic diarrhea viruses isolated in Korea[J].Virus Genes,2007,35:321-332.
[8] Duarte M,Laude H.Sequenee of the Spike Protein of the Porcine Epidemie DiarrhoeaVirus[J].J GenVirol,1994,75:1195-1200.
[9] Haijema B J,Volders H,Rottier P J.Live attenuated coronavims vaccines through the directed deletion of group- specific genes provide protection against feline infection peritonitis[J] .J Gen Virol,2004,78(8):3863-3871.
Isolation and identification of Porcine Epidemic Diarrhea Virus strain XJ - DB2 in Xinjiang Province
LIU Xiao-lan,HE Sun,CHEN Lan-ling,DU Jiu-bin
(Xinjiang tencon Livestock Biotechnology Co.,Ltd,Urumqi 830011,China)
Abstract:Porcine epidemic diarrhea virus(PEDV)XJ-DB2 strain was isolated from piglets suffering from a severe diarrhea in a pig farm of Xinjiang Province Dianba Area using Vero cell based on continuous passage culture and daily observation of cytopathic effect(CPE).The PEDV ORF3 gene was used for genetic and phylogenetic analysis.The results showed that the isolated XJDB2 was a virulent strain.Based on the genetic evolutionary,the PEDV strains which isolated in China(virulent CHS and CHGS)and South Korea(S-DR13)were in the same phylogenetic relationship,whereas the candidate vaccine strains CV777 attenuated DR13 and CHJS were in the other phylogenetic lineage.Compared with the ORF3 sequences published previously in GenBank by DNA Star software,the ORF3 sequence of XJ-DB2 strain had the highest nucleotide homology at 99.0%with CHS strain,and 98.8%with S-DR13.It also had 97.3%homology with the Vaccine strain CV777.
Key words:Porcine epidemic diarrhea virus;Isolation and identification;ORF3 gene;Genetic analysis
作者简介:刘小兰(1981-),女,助理兽医师,本科,从事兽用疫苗研发工作,E-mail:122830140@qq.com
收稿日期:2014-09-19
中图分类号:S852.65+1
文献标志码:A
文章编号:0529- 6005(2016)01- 0017- 03
猜你喜欢
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2016年11期)2017-03-21
湖北农业科学(2016年23期)2017-03-17
江苏农业科学(2017年1期)2017-02-27
湖北畜牧兽医(2016年9期)2017-02-13
山东农业科学(2016年11期)2016-12-17
江苏农业科学(2016年4期)2016-06-14
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年8期)2015-09-10
湖北农业科学(2014年11期)2014-09-10
