
A new microraptorine specimen (Theropoda: Dromaeosauridae) with a brief comment on the evolution of compound bones in theropods
2016-03-29 04:55:41XUXingLIFeng
古脊椎动物学报(中英文) 2016年4期
XU XingLI Feng,2
(1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044 xuxing@ivpp.ac.cn)
(2University of Chinese Academy of SciencesBeijing 100049)
A new microraptorine specimen (Theropoda: Dromaeosauridae) with a brief comment on the evolution of compound bones in theropods
XU Xing1LI Feng1,2
(1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044 xuxing@ivpp.ac.cn)
(2University of Chinese Academy of SciencesBeijing 100049)
Microraptorinae is a recently discovered subgroup of dromaeosaurid theropods, mostly comprising species from the Lower Cretaceous Jehol Group of western Liaoning, China. Here we describe a new microraptorine specimen from the Jiufotang Formation (the upper section of the Jehol Group) of Dapingfang, Chaoyang, Liaoning, which displays interesting morphological features not previously documented within Microraptorinae. Noteworthy are several osteological features, including dental and ischial ones, which are transitional between the condition ofSinornithosaurusand that ofMicroraptor.These features highlight the existence of a spectrum of morphological variation betweenSinornithosaurus, which is more like a typical dromaeosaurid, andMicroraptor, which shares many characteristics with troodontids. However, the taxonomic signi fi cance of these variations has not to be fully assessed yet. A feature deserving special mention is the fusion of the pubes to the ilia in this specimen, which has implications for the evolution of compound bones in theropods+. Our preliminary analysis suggests that many compound bones in birds have been formed by sequential fusion of multiple elements in a peramorphic process.
Lower Cretaceous, Jehol Group, Theropoda, Microraptorinae, variation, compound bones
1 Introduction
Microraptor zhaoianuswas erected in 2000 based on a fully articulated partial skeleton collected from the Jiufotang Formation (the upper section of the Lower Cretaceous Jehol Group) (Swisher et al., 2001) of Langshan, Chaoyang, western Liaoning, China (Xu et al., 2000). The holotype ofM. zhaoianuswas originally suggested to represent an adult individual based on fusion features (such as fusion among the sacral vertebrae, partial fusion of the lastdorsal to the synsacrum, and a complete pubic symphysis) and other features including fine serrations on the tooth crowns, a well-developed accessory crest on the femur, and presence of well-ossi fi ed cortical bone (Xu et al., 2000). Additional evidence for adult status includes the relatively small skull (skull/femur length ratio of 0.85) and sacrum (sacrum/femur length ratio of 0.36, compared to 0.44 inSinornithosaurus; inArchaeopteryx, however, the larger individuals have smaller sacrum/femur length ratio than inMicroraptor). However, the evidence is not conclusive and is contradicted by the absence of fusion of the distal carpals and tarsals either to each other or to the adjoining bones, which suggests a relatively early ontogenetic stage.
In 2003, a second species of the same genus,Microraptor gui, was erected based on two specimens that differed from the holotype ofM. zhaoianusin being considerably larger, and in having a prominent biceps tuberosity on the radius, a much shorter manual digit I, a strongly curved pubis, and a bowed tibia (Xu et al., 2003b). Senter et al (2004) synonymizedM. guiwithM. zhaoianus, based on stratigraphic proximity and a perceived absence of important anatomical differences (Senter et al., 2004). The former reason is not persuasive because closely related taxa can coexist temporally and geographically, though distinct temporal separation can be used as evidence to support taxonomic separation.
Nevertheless, there is reasonable evidence to synonymize the two taxa given that the holotype specimen ofM. zhaoianusis ontogenetically less advance than that ofM. gui(e.g., distal tarsals remain separate from the metatarsals in the former but appear to be fused to them in the latter). Accordingly, the putative interspecific differences listed by Xu et al. (2003b) might be plausibly interpreted as points of ontogenetic variation.
In addition toMicroraptor, fi ve other dromaeosaurid genera have been named based on specimens claimed to have been recovered from the Yixian Formation (the middle section of the Jehol Group; the lower section is called the Dabeigou Formation) of western Liaoning:Sinornithosaurus(Xu et al., 1999),Graciliraptor(Xu and Wang, 2004),Tianyuraptor(Zheng et al., 2010),Chanyuraptor(Han et al., 2014), andZhengyuanlong(Lü and Brusatte, 2015). TheSinornithosaurusfossil was collected by an expedition team from the Institute of Vertebrate Paleontology and Paleoanthropology, Beijing (IVPP); the other fossils were collected by local farmers and their provenance remains unconfirmed. Nevertheless, the fi ve Liaoning dromaeosaurids and the North American genusHesperonychus(Longrich and Currie, 2009) share a unique set of features and have been suggested to form a subclade, the Microraptorinae, within the Dromaeosauridae (Senter, 2007; Longrich and Currie, 2009; Han et al., 2014). However, some studies have placed one or more of these species outside the Microraptorinae (Zheng et al., 2010; Lü and Brusatte, 2015).
In the present paper, we describe a new microraptorine specimen (IVPP V 13476) from the Lower Cretaceous Jiufotang Formation of Dapingfang, Chaoyang, China, which displays interesting morphological characteristics not previously documented within Microraptorinae. We also comment on several features that have implications for both the taxonomy of the group and the evolution of some important avian characters.
2 Description and comparisons
IVPP V 13476 is probably an adult individual (Fig. 1), based on fusion features: all visible neurocentral sutures are closed, the pubes are fused to the ilia, and the distal tarsals are fused to the metatarsals. It is similar in size (Table 1) to the holotype ofMicroraptor gui(Xu et al., 2003a), and its body mass is estimated to be approximately 1.36 kg, based on an empirical equation (Christiansen and Fariña, 2004).
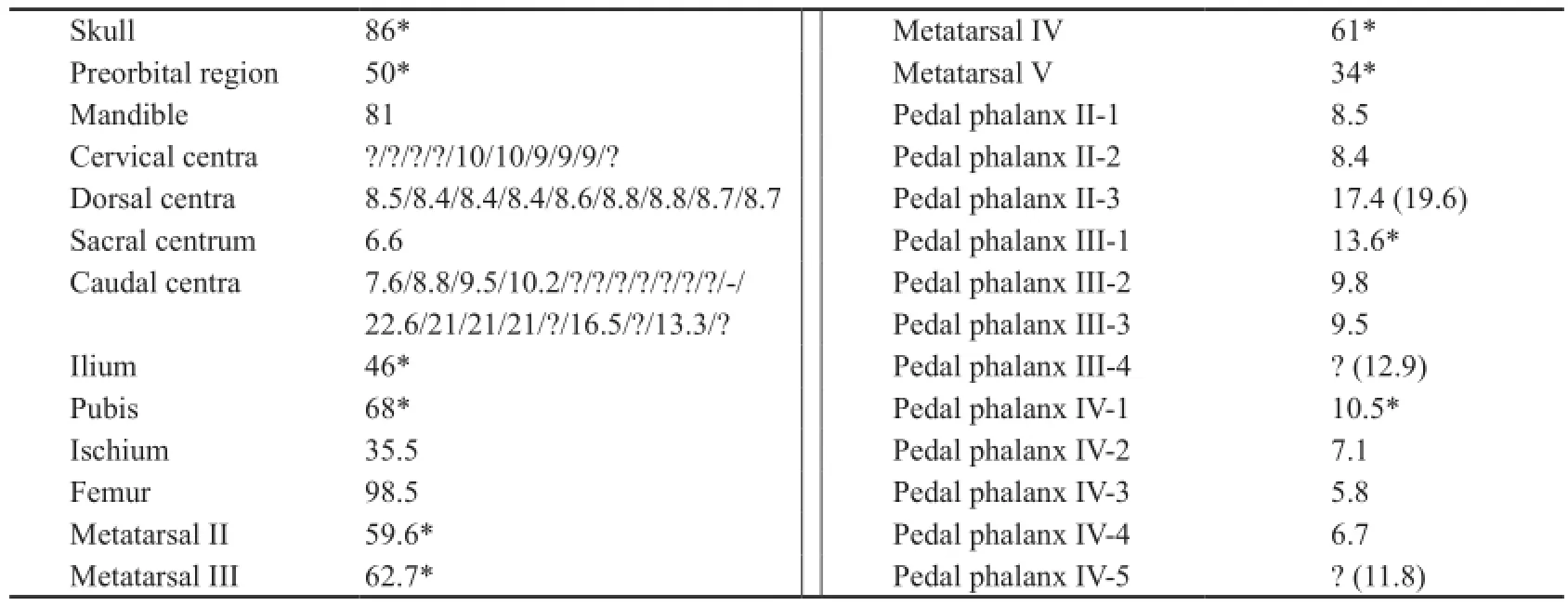
Table 1 Selected measurements of IVPP V 13476 (mm)
Skull and mandible The skull and mandible of IVPP V 13476 are exposed in lateral view (Fig. 2). Most of the cranial elements are preserved in articulation, but some are displaced. For example, the left premaxilla has moved posteriorly and the left maxilla is not only displaced posteriorly but also preserved upside down (Fig. 2C: lm). Although most of the cranial elements are still present, the poor preservation has resulted in a degree of distortion (e.g., the left and right external nares differ in shape: the right one is sub-rectangular and the left one is oval) and dif fi culties in collecting informative morphological data.
The skull of V 13476 measures about 85% of the femoral length and has a proportionally short snout, about 50% of the total skull length. Visible major cranial openings include the external naris, the antorbital fenestra, the orbit, the infratemporal fenestra, and the external mandibular fenestra (Fig. 2C: en, aof, orb, itf, emf). It is noteworthy that the antorbital fenestra is relatively small and the external mandibular fenestra is relatively posteriorly located.
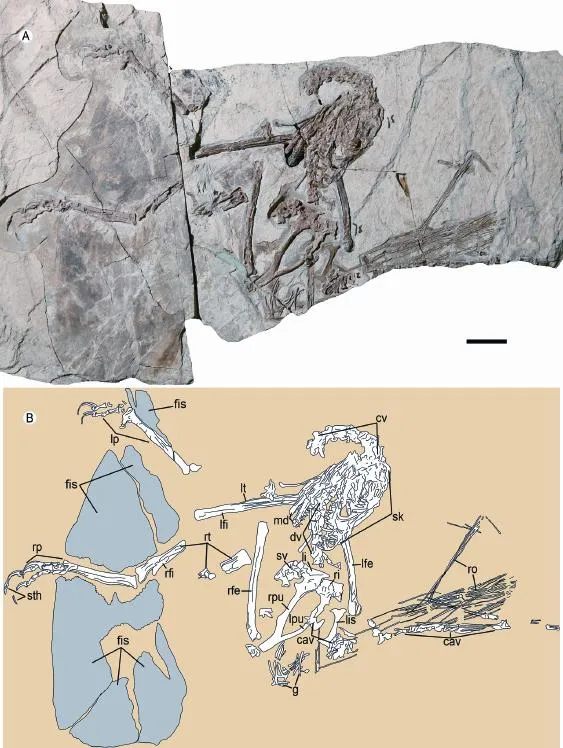
Fig. 1 Photograph (A) and line-drawing (B) of Microraptorine IVPP V 13476 Abbreviations: cav. caudal vertebra 尾椎; ch. chevron 人字骨; cv. cervical vertebra 颈椎; dv. dorsal vertebra背椎; g. gastralia 腹肋; lfe. left femur 左股骨; l fi. left fi bula 左腓骨; li. left ilium 左肠骨; lis. left ischium 左坐骨; lp. left pes 左脚; lpu. left pubis 左耻骨; lt. left tibia 左胫骨; md. mandible 下颌骨; rfe. right femur 右股骨; r fi. right fi bula 右腓骨; ri. right ilium 右肠骨; ro. rod-like extensions 棒状延伸; rp. right pes 右脚; rpu. right pubis 右耻骨; rt. right tibia 右胫骨; sk. skull 颅骨; sv. sacral vertebra 荐椎Scale bar equals 3 cm
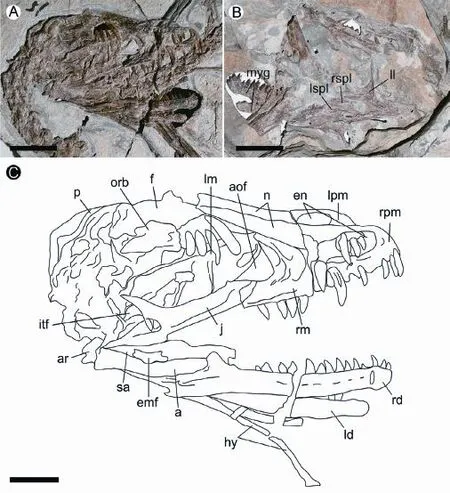
Fig. 2 Skull and mandible in Microraptorine IVPP V 13476 A. in right lateral view; B. in left lateral view; C. line-drawing of the skull Abbreviations: a. angular 隅骨; aof. antorbital fenestra 眶前孔; ar. articular 关节骨; emf. external mandibular fenestra 外下颌骨孔; en. external naris 外鼻孔; f. frontal 额骨; hy. hyoid 舌骨; itf. infratemporal fenestra下颞孔; j. right jugal 颧骨; ld. left dentary 左齿骨; ll. left lacrimal 左泪骨; lm. left maxilla 左上颌骨; lpm. left premaxilla 左前上颌骨; lspl. left splenial 左夹板骨; myg. Meckelian groove 麦氏沟; n. nasal 鼻骨; orb. orbit 眼眶; p. parietal 顶骨; rd. right dentary 右齿骨; rm. right maxilla 右上颌骨; rpm. right premaxilla 右前上颌骨; rspl. right splenial 右夹板骨; sa. surangular 上隅骨. Scale bars equal 2 cm
Both premaxillae are preserved in V 13476 (Fig. 2C: lpm, rpm). Most of the lateral side of the right premaxilla is exposed, as is part of the medial side of the left premaxilla. Due to the poor preservation, the two premaxillae display some significant differences. The right premaxilla has a nearly vertical anterior margin, with a premaxillary angle of about 80°, but the left premaxilla has a posterodorsally directed anterior margin and a premaxillary angle of approximately 70°. The nasal process forms about half of the dorsal border of the external naris and covers the premaxillary process of the nasal for a considerable distance. The prenarial portion of the premaxilla is much higher dorsoventrally than long anteroposteriorly. The narial fossa is well developed both ventrally and anteriorly (Fig. 2A, C). A smaller, deep fossa lies within the anterior portion of the narial fossa (Fig. 2A, C). Anterior to the narial fossa is a distinct, rounded, nearly vertical ridge terminating slightly dorsal to the ventral border of the external naris. Immediately anterior to the ridge is a sharp groove about 3 mm long. The dorsally-faced sub-narial shelf is prominent, and its lateral margin bears a distinct rounded ridge terminating anteriorly at the level of the anterior border of the external naris. Themaxillary process of the premaxilla is short and slender, and appears not to extend to the level of the posterior border of the external naris.
The right nasal is well exposed in V 13476, and accounts for more than 40% the length of the skull roof (Fig. 2C: n). The nasal has the shape of an elongated sub-rectangle, being more than 6 times as long as wide. The nasal is widest near the mid-length, with both ends slightly narrower than the middle portion in dorsal view. The anterior part of the nasal appears dorsoventrally thick and somewhat rugose. The nasal sends anteroventrally a short and slender process, which appears to contact the maxillary process of the premaxilla at the mid-length of the ventral border of the external naris (Fig. 2A, C). The dorsal surface of the nasal is mostly fl at and smooth, although a few small pits are present around the mid-length of the bone. The contact with the maxilla is extensive, occupying nearly whole length of the lateral margin of the nasal. The most unusual feature of the nasal is the apparent presence of a distinct, anteroposteriorly elongated lateral flange occurring near the mid-length of the nasal and overhanging the lateral side of the snout, but this might be a preservational artifact (Fig. 2A). The combined posterior margin of the two nasals appears to be concave in dorsal view.
Both maxillae are preserved in V 13476 (Fig. 2C: lm, rm). Combining information from both sides of the skull suggests a complete maxilla would be triangular, about 40 mm in maximum anteroposterior length, and about 25 mm in the maximum height, with the highest point at the posterior end of the bone. The length of the maxilla, measured along the ventral margin, is more than 50% of the total skull length. Although the anterior ends of both maxillae are poorly preserved, the bone is clearly shallow in pro fi le (Fig. 2A, C). The maxilla houses an antorbital fossa with a sharp ventral rim and an antorbital fenestra that has a rounded anterior border and an anteroposteriorly narrow osseous medial wall as inTsaagan mangas(Norell et al., 2006) andLinheraptor exquisitus(Xu et al., 2015). The dorsally displaced maxillary fenestra lies in a recess, a feature also seen in many other dromaeosaurids includingM. zhaoianus(Xu, 2002) andVelociraptor mongoliensis(Barsbold and Osmólska, 1999). As in other Liaoning dromaeosaurids, short grooves and ridges are present within the anterior portion of the antorbital fossa.
Most of the left lacrimal is exposed (Fig. 2B: ll). The lacrimal is a roughly T-shaped bone, but the descending process forms a sharp angle with the anterior process (about 60°) and an obtuse angle with the posterior process (about 120°). A large, shallow fossa is present in the area where the anterior and posterior processes come together, and the medial surface of the dorsal half of the descending process bears a sharp ridge along the posterior margin. The posterior process is very short, and thin dorsoventrally.
The remaining skull elements preserve little informative morphological data. The tentatively identified right jugal has an unusually deep infraorbital ramus, and its ascending process is oriented posterodorsally (forming an angle of about 40° with the posterior process) (Fig. 2C: j). The frontal is a rather large bone, forming much of the skull roof (Fig. 2C: f). In lateral view the frontal describes a strong curve, at fi rst gently rising dorsally as it extends posteriorly but then evidently arching downwards at the posterior end (Fig. 2A, C). The parietal is proportionally short anteroposteriorly, occupying only a small portion of the skull roof (Fig. 2C: p).
Both dentaries are relatively well preserved (Fig. 2C: ld, rd). As in other dromaeosaurids, the dorsal and ventral margins of the dentary are sub-parallel (concave and convex in lateral view, respectively). The anterior end of the dentary tapers to form a narrow point at the alveolar margin and is also slightly downturned. A row of elongated oval foramina is present on the lateral surface of the dentary below the dorsal margin. Some of the foramina are connected by grooves, forming a single incompletely continuous groove on the lateral surface of the dentary. Ventral to the groove is a second row of foramina, which are less well developed and fewer in number. In medial view, the mandibular symphysial surface is flat, very narrow anteroposteriorly, and anteromedially directed (Fig. 2B). The Meckelian groove is deep, and located close to the ventral margin of the dentary (Fig. 2B: myg).
Both splenials are preserved, but neither is completely exposed (Fig. 2B: lspl, rspl). Although the splenial is largely positioned on the medial side of the mandible, the middle portion of the bone is thickened to wrap around the ventral margin of the dentary and even develops a small lateral lamina. The posterior margin of the splenial bears a small notch, situated close to the dorsal end. The surangular has a convex dorsal margin, and the anterior portion of the angular appears deep (Fig. 2C: a, sa).
Dentition Each premaxilla preserves only the anterior three of the expected four premaxillary teeth (Fig. 3A). The two premaxillae display different stages in the tooth replacement cycle: in the right premaxilla the first tooth is the largest and the second is the smallest, but in the left premaxilla the first tooth is the smallest and the second is much larger. As in many coelurosaurs, all preserved premaxillary teeth lack serrations on both the anterior and posterior carinae. The premaxillary tooth crowns are slightly recurved. They each have a convex lingual surface but a fl at labial surface, making the cross-section D-shaped, and a slight basal constriction.
Nine teeth are preserved on the right maxilla, and at least four additional teeth are missing (Fig. 3B, C). The posteriormost maxillary tooth is located close to the posterior end of the jugal process of the maxilla. The maxillary teeth are similar in general morphology to the premaxillary teeth, but are larger, stouter, and more strongly recurved. All preserved maxillary teeth lack serrations on the anterior carina. However, a degree of variation exists along the maxillary tooth row: the posterior teeth are more robust than the anterior ones and have better developed posterior serrations (posterior serrations are absent in the anteriormost five maxillary teeth, weakly developed in the 6thtooth, and well-developed in the 7ththrough 11thteeth).
Sixteen teeth are preserved on the right dentary, which may represent a complete dentary tooth row (Fig. 3E, F). The dentary teeth are smaller than the maxillary teeth. As with the premaxillary and maxillary teeth, the dentary tooth crowns are somewhat shorter than the corresponding roots (one isolated tooth shows that the root is 1.3 times as long as the crown) (Fig. 3E, F). Variation also exists along the dentary tooth row: towards the posterior end of the row, the teeth become more robust, the lingual surfaces become more convex, and the constrictions at the bases of the crowns become more prominent (Fig. 3E, F). The anteriormost four dentary teeth lack both anterior and posterior serrations, and have weakly convex lingual surfaces which bearweak longitudinal grooves and ridges. The 5thdentary tooth is transitional: serrations are absent on the anterior carina and rudimentary on the posterior carina, being de fi ned only by blood groovesas opposed to notches that separate adjoining serrations along the tooth carina (Fig. 3E). The 6ththrough 14thdentary teeth lack anterior serrations but have well de fi ned posterior serrations. The 15thand 16thdentary teeth have both anterior and posterior serrations, with the latter signi fi cantly larger than the former. Replacement pits are present on the medial side of the tooth roots. In the 7thalveolus, a replacement tooth is clearly visible within the root of the functional tooth.
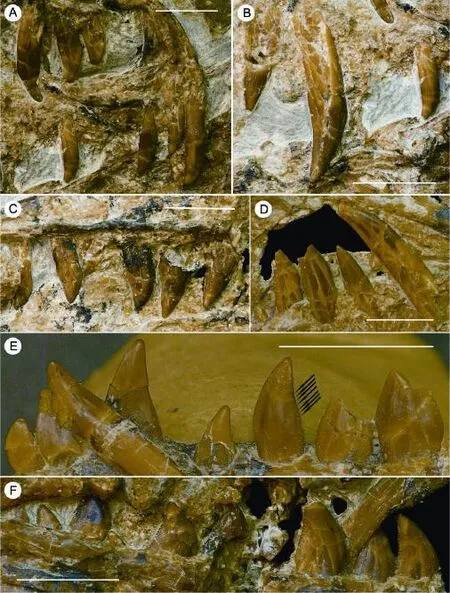
Fig. 3 Close-ups of the dentition in Microraptorine IVPP V 13476 A. Premaxillary teeth in right lateral view (with the 4thtooth missing on both sides) 前上颌齿右侧视(双侧第四前上颌齿缺失); B. Anterior right maxillary teeth (3rd?, 6thand 7th) in labial view 前部右上颌齿(第三?, 第六和第七上颌齿)唇侧视; C. Posterior right maxillary teeth (8thto 13th) in labial view 后部右上颌齿(第八至第十三右上颌齿)唇侧视; D. 6thto 10thleft maxillary teeth in labial view (upside down) 第六至第十左上颌齿唇侧视(上下颠倒); E. 1stto 7thright dentary teeth in lingual view, black arrows indicate the blood grooves on the posterior carina of the 5thdentary tooth 第一至第七右下颌齿舌侧视, 第五下颌齿后齿脊的血槽如黑色箭头所示; F. 8thto 16thright dentary teeth in labial view 第八至第十六右下颌齿唇侧视Scale bars equal 5 mm

Fig. 4 Close-ups of selected parts of postcranial skeleton in Microraptorine IVPP V 13476 A. cervical vertebrae 5thto 7th(anticlockwise) in ventral view 第五至第七颈椎(逆时针)腹侧视; B. posterior dorsal vertebrae in left lateral view 后部背椎左侧视; C. isolated sacral vertebra in posteroventral view 分散荐椎后腹侧视; D. three anterior caudal vertebrae in ventral view 三枚前段尾椎腹侧视; E. articulated posterior caudal vertebrae in left lateral view 相连的后段尾椎左侧视; F. 1stand 2ndchevrons in left lateral view 第一和第二人字骨左侧视; dorsal (G) and ventral (H) views of two isolated middle chevrons两枚分离的中段人字骨背侧视G和腹侧视H; I. unique stiffening rods formed by highly elongated anterior processes of chevrons and equally long prezygapophyses of most caudal vertebrae 由人字骨高度伸长的前突和同样伸长的尾椎前关节突形成的棒状结构; J. fused pubes in right oblique view 愈合耻骨右斜视; K. left ischium in lateral view 左坐骨外侧视; L. proximal end of right femur in posterior view 右股骨近端后侧视; M. right ankle in posterior view 右踝关节后侧视; N. right tarsometatarsus in posterior view 右跗跖骨后侧视; O. right pes in medial view 右脚内侧视. Scale bars equal 1 cm
Axial skeleton The anteriormost nine cervical vertebrae are exposed, and most remain in articulation (Fig. 4). However, little can be said about their morphology due to poorpreservation. The relatively well preserved cervicals 5-7 show that the centra are considerably narrower transversely than the neural arches, and that the ventral surfaces of the centra are probably relatively fl at transversely (Fig. 4A). Moving posteriorly along the series, the centra become wider transversely.
Nine dorsal vertebrae are exposed, most remaining in articulation, and they probably represent the posteriormost nine in the series. As with the cervical vertebrae, little morphological information is available. The dorsal centra are relatively long (ratio of length of centrum to height of posterior end is 1.4 in a posterior dorsal vertebra), and none of them bears a distinct pneumatic foramen on the lateral surface (Fig. 4B).
Only one sacral is visible in IVPP V 13476. The centrum of this only exposed sacral vertebra has a rounded ventral surface (Fig. 4C).
Twenty caudal vertebrae are preserved. As in other Liaoning dromaeosaurids, the caudal vertebrae are significantly elongated, the longest preserved caudal being about 2.5 times as long as the posterior dorsal vertebrae. The anteriormost caudal vertebrae have box-like centra that are much wider transversely than deep dorsoventrally, and each bear a well-defined longitudinal groove along the ventral surface (Fig. 4D). The distal ends of the transverse processes of these vertebrae are strongly expanded anteroposteriorly, as inSinornithosaurus. The centra of the middle caudal vertebrae are also box-like, with fl at ventral surfaces, but are deeper dorsoventrally than wide transversely (Fig. 4E). The posterior caudal centra appear to be much narrower transversely than the corresponding neural arches, as in basal troodontids, and bear longitudinally elongate depressions on their lateral surfaces (Fig. 4E).
The chevrons display a diverse range of morphologies. Two isolated chevrons, probably representing the first and second in the series, are typical of theropod chevrons in being laterally compressed plates whose dorsoventral length greatly exceeds their anteroposterior width (Fig. 4F). The proximal end of each of these chevrons is bridged, forming a closed haemal canal, and an accessory process is present on the posterior margin immediately ventral to the haemal canal. Five isolated chevrons, probably representing chevrons 3-7 are strongly distally expanded in the anteroposterior direction and bear more prominent accessory posterior processes (Fig. 4G). One isolated chevron, which is more posteriorly located in the caudal series, bears two horizontally oriented processes that respectively extend anterior and posterior to the articular facet for the caudals (Fig. 4H). The anterior process is a dorsoventrally thin plate, but the posterior process is less compressed dorsoventrally and is narrower transversely. The remaining chevrons are about 13 in number and have bifurcated anterior and posterior processes, the anterior processes also being highly elongated. These highly elongated anterior processes, together with the equally long prezygapophyses of most of the caudal vertebrae, form the distinctive rod-like structures seen in the tails of most dromaesaurids (Fig. 4I).
Pelvic girdle Both ilia are missing the anterior ramus, and the left ilium is overlappedposteriorly by the right (Fig. 4J). The pubes are almost complete, but are damaged at their distal ends, and the right ischium is absent (Fig. 4K).
The right ilium appears slightly distorted, the acetabular region being de fl ected laterally (Fig. 4C). However, the possibility that this re fl ects the true morphology of the ilium cannot be excluded, because a similar feature is present in a therizinosauroid from the Upper Cretaceous in Nei Mongol (Zhang et al., 2001; Xu et al., 2002). Although part of the preacetabular process is missing, it can be safely inferred that the ilium is significantly shorter than the femur. A proportionally small ilium is also seen in some basal dromaeosaurids, troodontids, and birds (Xu, 2002). There is a concavity on the posterodorsal margin of the postacetabular process, a feature also seen in several other basal paravians includingRahonavis.A similar, but less well-developed, concavity is also present inArchaeopteryx. The most unusual feature is the strong ventral de fl ection of the postacetabular process, the posterior end of which is signi fi cantly ventral to the pubic peduncle of the ilium (Fig. 4C).
The pubes are fused to each other distally to form a conjoined pubic boot. As in other microraptorines, a prominent lateral tubercle is present at about the mid-length of the pubic shaft. The two pubes are completely fused to the ilia, a condition previously unreported in any microraptorine (Fig. 4J).
The ischium is a short plate (ischium/femur length ratio 0.36) with a distally located obturator process, as in other microraptorines (Fig. 4K). However, some variation in ischial morphology occurs among microraptorines. The ischium is relatively slender in lateral view in V 13476 andMicroraptor gui, but is wider inSinornithosaurusandTianyuraptor.The obturator process is relatively small in V 13476 andM. gui(obturator process/ischium length ratio 0.44 and 0.43 in V 13476 andM. gui, respectively), but extremely large inSinornithosaurus(corresponding ratio at least 0.75). In IVPP V 13476 andM. gui, the proximoposterior process and the groove along the anterior margin of the ischial shaft are minimal, but they are better developed inSinornithosaurus.However, IVPP V 13476 is similar toSinornithosaurusin that the obturator process meets the ischial shaft at a sharp angle, while the equivalent angle is considerably larger inM. gui.
Hindlimbs The hindlimbs are generally similar to those of other microraptorines in both general morphology and proportions. The femur is an anteriorly bowed element. One feature that deserves special mention is that the trochanteric crest appears to extend further proximally than the femoral head (Fig. 4L). The tibiotarsus and fi bula are poorly preserved, and little morphological information concerning these bones is obtainable.
The pes displays several interesting features. Individual distal tarsals cannot be identified, as these bones are fused to the metatarsals (Fig. 4M). The proximal articular surface of the tarsometatarsus is fl at and slants distomedially in anterior or posterior view. The proximal end of the unit is expanded transversely, and particularly in the lateraldirection. Although the posterior surface of the metatarsus is partially crushed, it is clear that the proximal part of metatarsal III is extremely thin in posterior view, suggesting a sub-arctometatarsalian condition. The metatarsals display few diagnostic features, though metatarsal V displays a typical microraptorine morphology: it is long, curved, and expanded near the middle of its length (Fig. 4N). InSinornithosaurus, metatarsal V is rod-like along its entire length; in V 13476, the distal half of metatarsal V appears to be strap-like as inM. gui. The second pedal digit is specialized in being highly extensible and equipped with a large raptorial claw (Fig. 4O). However, the proximoventral heel of pedal phalanx II-2 projects only a short distance proximally, unlike inM. guiand other Liaoning dromaeosaurids.
Plumage IVPP V 13476 preserves pennaceous feathers posterior to the tibiotarsus and the tarsometatarsus (Fig. 5A). The feathers form a sharp angle with the tarsometatarsus, and an obtuse angle with the tibiotarsus. The longest feathers attached to the tarsometatarsus measure at least 140 mm, much longer than those attached to the femur. The dense arrangement of the feathers prevents an accurate count, but some morphological details are obtainable. The feathers each have well-developed vanes and a prominent rachis (Fig. 5B-D). One feather located at the distal end of the left tarsometatarsus has a leading vane approximately 2 mm wide and a trailing vane 4 mm wide, measured about one-fourth of the distance from the tip (Fig. 5B). Near the middle of its length, this feather bears 16 barbs in every 5 mm of distance along the rachis. These barbs form an angle of about 30° with the rachis.

Fig. 5 Plumage of Microraptorine IVPP V 13476 Pennaceous feathers attached to tibiotarsus and the tarsometatarsus (A); Close-up of feathers attached to distal left tarsometatarsus (B), distal right tarsometatarsus (C), and right tibiotarsus (D) Scale bar equals 6 cm in A, 1 cm in B to D
3 Discussion
Taxonomic status and morphological variation IVPP V 13476 is referable to the Microraptorinae based on the following features (Xu, 2002; Turner et al., 2012): numerous short ridges and pits in anterior portion of antorbital fossa, most teeth lacking anterior serrations, highly elongated caudal prezygapophyses and chevrons, pubis with lateral tubercle near mid-shaft, ischium short and with distally located obturator process, metatarsus subarctometatarsalian, metatarsal II slender, metatarsal IV robust, and pedal digit II hyperextensible but relatively small.
However, V 13476 is clearly distinguishable from other Liaoning dromaeosaurids, includingMicroraptor, in some dental, ischial, and pedal features. For example, compared toMicroraptor, V 13476 displays a smaller angle between the obturator process and the ischial shaft and a smaller proximoventral heel on pedal phalanx II-2.
The most interesting difference concerns the dental morphology of V 13476, which is transitional between that ofSinornithosaurus,Graciliraptor, andTianyuraptoron the one hand and that ofMicroraptoron the other. The dentition ofSinornithosaurus(Xu and Wu, 2001),Graciliraptor(Xu and Wang, 2004), andTianyuraptor(Zheng et al., 2010) is more similar to that of a typical carnivorous theropod in displaying the following features: tooth count relatively small (approximately 13 dentary teeth inSinornithosaurus); tooth crowns slender, blade-like and posteriorly curved; tooth serrations relatively well developed (middle and posterior teeth with both anterior and posterior serrations inSinornithosaurus); and teeth relatively homodont and evenly distributed.Microraptorhas a number of dental features (many of which are also seen in troodontids) that are absent in many other dromaeosaurids (Xu et al., 2000; Hwang et al., 2002): tooth number relatively large (approximately 19 dentary teeth); tooth crowns stout and basally constricted; tooth serrations relatively poorly developed (all teeth without anterior serrations, and posterior serrations present only in middle and posterior teeth); and teeth relatively heterodont and unevenly distributed (anterior teeth slender and closely packed, and posterior ones stout and sparsely spaced). V 13476 displays an intermediate condition with respect to tooth number (16 dentary teeth), tooth crown shape (relatively stout and weakly curved posteriorly), and the development of serrations (only posteriormost teeth have both anterior and posterior serrations), but is more similar toSinornithosaurus,Graciliraptor, andTianyuraptorin having relatively evenly distributed and homodont teeth. The diversity of dental morphology seen in Jehol dromaeosaurids suggests that some variation in dietary preferences existed within this group.
These differences from previously described taxa suggest that V 13476 might represent a new microraptorine species. However, there is a wide spectrum of morphological variation in Liaoning dromaeosaurids, some of which may reflect ontogeny, sexual dimorphism or individual variation. In the absence of extensive data that would make it possible to properly evaluate these variations, we refrain from naming a new taxon based on this specimen.
Evolution of compound bones in theropods IVPP V 13476 preserves several features previously unreported in other Liaoning dromaeosaurids, including a strongly downturned anterior end of the ilium, bifurcated anterior and posterior processes of the chevrons, and fusion of the ischia to the ilia. The last of these features is especially notable.
Pelvic fusion is unusual among non-avialan theropods, but common among birds. In fact, the skeletons of the latter group are characterized by the presence of many compound bones formed by the ontogenetic fusion of multiple skeletal elements (King and McLelland, 1984; Bellairs and Osmond, 1998). For example, several dorsal vertebrae fuse to form the notarium in many birds, a large number of vertebrae fuse to form the synsacrum, the posterior caudals fuse to form the pygostyle, the three pelvic elements fuse into a single skeletal unit, the distal carpals fuse to the metacarpals to form the carpometacarpus, the proximal tarsals fuse to the tibia to form the tibiotarsus, and the distal tarsals fuse to the metatarsals to form the tarsometatarsus (King and McLelland, 1984; Baumel and Witmer, 1993; Bellairs and Osmond, 1998; Clarke, 2004). This extensive skeletal fusion stiffens and reinforces many parts of the skeleton, better equipping it to withstand the stresses involved in both terrestrial and aerial locomotion (King and McLelland, 1984).
By contrast, fusion of multiple skeletal elements to form compound bones is rare in nonavialan theropods, but examples nevertheless are known in some clades. For example, the notarium is not present in any known non-avialan theropod, but fusion of two posterior dorsals occurs inSinornithosaurus(Xu, 2002). Fusion of the posteriormost caudals to form a pygostyle or pygostyle-like structure appeared independently in the therizinosauroidBeipiaosaurus(Xu et al., 2003a) and several oviraptorosaurians (Barsbold et al., 2000; Persons et al., 2013). A compound tarsometatarsus has evolved independently in some ceratosaurs (Rowe and Gauthier, 1990), several alvarezsauroids (Turner et al., 2009; Xu et al., 2010), the oviraptorosaurianAvimimus(Kurzanov, 1987), and several basal deinonychosaurians (Xu, 2002).
Fusion of the pubis to the ilium inMicroraptorhas important implications for current understanding of the evolution of a compound pelvic bone. Among non-avialan maniraptorans, pelvic fusion has been reported previously only in the dromaeosauridHesperonychus(Longrich and Currie, 2009) and the oviraptorosaurianAvimimus(Kurzanov, 1987), though the ilium, pubis, and ischium are fused in adult individuals of many relatively basal theropods including some coelophysids and ceratosaurs (Rowe and Gauthier, 1990). Interestingly, in bothMicroraptorandHesperonychusthe pubis is fused to the ilium, but the ischium remains separate (the ischium is not preserved in the only known specimen ofHesperonychus, but is inferred to have been separate at least from the ilium based on the smooth articular surface of the ischial peduncle). This suggests that the avialan compound pelvis formed sequentially, with fusion of the ischium to the pubis and ilium representing the last step.
Among basal birds, the three pelvic elements remain separate in the known individuals ofArchaeopteryx(Elzanowski, 2002; Wellnhofer, 2009). In all known individuals including small-sized ones (presumably ontogenetically early ones) of the basal avialansSapeornis,Jeholornis, and confuciusornithids, the pubis appears to be fused to the ilium whereas the ischium remains separate (Chiappe et al., 1999; Zhou and Zhang, 2003a, b). In basal ornithuromorphans, all three pelvic elements seem to be fused together very early in ontogeny (Zhou and Zhang, 2001; 2006) as in crown group birds (King and McLelland, 1984; Bellairs and Osmond, 1998). These data support the interpretation that the compound pelvic bone was built up in tetanuran evolution by initial fusion of the pubis to the ilium followed by subsequent incorporation of the ischium, and that the fusions occur earlier ontogenetically in more crown-ward taxa along the line to modern birds.
This sequential fusion in combination of ontogenetically earlier fusions in more crown-ward theropod taxa seems to apply to the evolution of other compound bones. The compound carpometacarpus may have evolved through a stepwise process in which the distal carpals (speci fi cally, distal carpals 3 and 4) fused initially to each other (Xu et al., 2014) and subsequently fused to the metacarpals. Similarly, the two proximal tarsals may have fused initially to one another, and subsequently to the tibia, to form the tibiotarsus. In both cases, the fusions occur at an earlier ontogenetic stage in more crown-ward theropod taxa. This suggests that the formation of compound bones in birds is a peramorphic process, whereas the avialan cranial con fi guration is largely a product of paedomorphosis (Bhullar et al., 2012). However, much more data will be needed to con fi rm this evolutionary interpretation.
Acknowledgements We thank Corwin SULLIVAN for commenting on the manuscript, DING Xiao-Qing for preparing the specimen, and GAO Wei and ZANG Hai-Long for photographs. This study was supported by the National Natural Science Foundation of China (41120124002) and the 973 program (2012CB821900).
Barsbold R, Osmólska H, 1999. The skull ofVelociraptor(Theropoda) from the Late Cretaceous of Mongolia. Acta Palaeont Pol, 44: 189-219
Barsbold R, Osmólska H, Watabe M et al., 2000. A new oviraptorosaur (Dinosauria, Theropoda) from Mongolia: the fi rst dinosaur with a pygostyle. Acta Palaeont Pol, 45: 97-106
Baumel J J, Witmer L M, 1993. Osteologia. In: Baumel J J, King A S, Breazile J E et al. eds. Handbook of Avian Anatomy: Nomina Anatomica Avium 2nded. Cambridge: Publicartions of the Nuttal Ornithological Club. 23: 45-132
Bellairs R, Osmond M, 1998. The Atlas of Chick Development. San Diego: Academic Press. 323
Bhullar B A S, Marugán-Lobón J S, Racimo F et al., 2012. Birds have paedomorphic dinosaur skulls. Nature, 487: 223-226
Chiappe L M, Ji S A, Ji Q et al., 1999. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the late Mesozoic of Northeastern China. Bull Am Mus Nat Hist, 242: 1-89
Christiansen P, Fariña R A, 2004. Mass prediction in theropod dinosaurs. Hist Biol, 16: 85-92
Clarke J, 2004. Morphology, phylogenetic taxonomy, and systematics ofIchthyornisandApatornis(Avialae: Ornithurae). Bull Am Mus Nat Hist, 286: 1-179
Elzanowski A, 2002. Archaeopterygoidae. In: Chiappe L M, Witmer L M eds. Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. 129-159
Han G, Chiappe L M, Ji S A et al., 2014. A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid fl ight performance. Nat Commun, 5: 4382
Hwang S H, Norell M A, Ji Q et al., 2002. New specimens ofMicroraptor zhaoianus(Theropoda: Dromaeosauridae) from northeastern China. Am Mus Novit, 3381: 1-44
King A S, McLelland J, 1984. Birds: Their Structure and Function. London: Baillière Tindall. 84-109
Kurzanov S M, 1987. Avimimidae and the problem with the origin of birds. Joint Soviet-Mongol Sci Res Geol Exped Trans, 31: 5-92
Longrich N R, Currie P, 2009. A microraptorine (Dinosauria-Dromaeosauridae) from the Late Cretaceous of North America. PNAS, 106: 5002-5007
Lü J, Brusatte S L, 2015. A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution. Sci Rep, 5: 11775
Norell M A, Clark J M, Turner A et al., 2006. A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia). Am Mus Novit, 3545: 1-51
Persons W S, Currie P J, Norell M A, 2013. Oviraptorosaur tail forms and functions. Acta Palaeont Pol, 59: 553-567
Rowe T, Gauthier J, 1990. Ceratosauria. In: Weishampel D B, Dodson P, Osmolska H eds. The Dinosauria. Berkeley: University of California Press. 151-168
Senter P, 2007. A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda). J Syst Palaeont, 5: 1-35
Senter P, Barsbold R, Britt B B et al. 2004. Systematics and evolution of Dromaeosauridae (Dinosauria, theropoda). Bull Gunma Mus Nat Hist, 8: 1-20
Swisher C, Wang X L, Zhou Z H et al., 2001. Further support for a Cretaceous age for the feathered-dinosaur beds of Liaoning, China: new40Ar/39Ar dating of the Yixian and Tuchengzi formations. Chin Sci Bull, 46: 2009-2013
Turner A H, Nesbitt S J, Norell M A, 2009. A large alvarezsaurid from the Late Cretaceous of Mongolia. Am Mus Novit, 3648: 1-14
Turner A H, Makovicky P J, Norell M A, 2012. A review of dromaeosaurid systematics and paravian phylogeny. Bull Am Mus Nat Hist, 371: 1-206
Wellnhofer P, 2009.Archaeopteryx- The Icon of Evolution. München: Verlag Dr. Friedrich Pfeil. 1-208
Xu X, 2002. Deinonychosaurian fossils from the Jehol Group of western Liaoning and the coelurosaurian evolution. Ph.D. Dissertation. Beijing: Graduate School of the Chinese Academy of Sciences
Xu X, Wang X L, 2004. A new dromaeosaur (Dinosauria:Theropoda) from the Early Cretaceous Yixian Formation of western Liaoning. Vert PalAsiat, 42: 111-119
Xu X, Wu X C, 2001. Cranial morphology ofSinornithosaurus milleniiXu et al. 1999 (Dinosauria, Theropoda, Dromaeosauridae) from the Yixian Formation of Liaoning, China. Can J Earth Sci, 38: 1739-1752
Xu X, Wang X L, Wu X C, 1999. A dromaeosaurid dinosaur with a fi lamentous integument from the Yixian Formation of China. Nature, 40: 262-266
Xu X, Zhou Z H, Wang X L, 2000. The smallest known non-avian theropod dinosaur. Nature, 408: 705-708
Xu X, Zhang X H, Sereno P et al., 2002. A new therizinosauroid (Dinosauria, Theropoda) from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol. Vert PalAsiat, 40: 228-240
Xu X, Cheng Y, Wang X et al., 2003a. Pygostyle-like structure from Beipiaosaurus (Theropoda, Therizinosauroidea) from the Lower Cretaceous Yixian Formation of Liaoning, China. Acta Geol Sin, 77: 294-298
Xu X, Zhou Z H, Wang X L et al., 2003b. Four-winged dinosaurs from China. Nature, 421: 335-340
Xu X, Wang D Y, Sullivan C et al., 2010. A basal parvicursorine (Theropoda: Alvarezsauridae) from the Upper Cretaceous of China. Zootaxa, 2413: 1-19
Xu X, Han F L, Zhao Q, 2014. Homologies and homeotic transformation of the theropod ‘semilunate’ carpal. Sci Rep, 4: 6042
Xu X, Pittman M, Sullivan C et al., 2015. The taxonomic status of the Late Cretaceous dromaeosauridLinheraptor exquisitusand its implications for dromaeosaurid systematics. Vert PalAsiat, 53: 29-62
Zhang X H, Xu X, Zhao X J et al., 2001. A long necked therizinosauroid dinosaur from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol, People’s Republic of China. Vert PalAsiat, 39: 282-290
Zheng X T, Xu X, You H L et al., 2010. A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution. Proc R Soc B: Biol Sci, 277: 211-217
Zhou Z H, Zhang F C, 2001. Two new ornithurine birds from the Early Cretaceous of western Liaoning, China. Chin Sci Bull, 46: 1-7
Zhou Z H, Zhang F C, 2003a. Anatomy of the primitive birdSapeornis chaoyangensisfrom the Early Cretaceous of Liaoning, China. Can J Earth Sci, 40: 731-747
Zhou Z H, Zhang F C, 2003b.Jeholorniscompared toArchaeopteryx, with a new understanding of the earliest avian evolution. Naturwissenschaften, 90: 220-225
Zhou Z H, Zhang F C, 2006. Mesozoic birds of China - a synopitic review. Vert PalAsiat, 44: 74-98
小盗龙亚科(兽脚亚目:驰龙科)一新标本及简评复合骨在兽脚亚目中的演化
徐 星1李 峰1,2
(1 中国科学院古脊椎动物与古人类研究所, 中国科学院脊椎动物演化与人类起源重点实验室 北京 100044)
(2 中国科学院大学 北京 100049)
小盗龙亚科是驰龙科兽脚类恐龙中新近发现的一个亚群,多数属种产自中国辽宁西部下白垩统热河群。本文描记一件产自辽宁省朝阳县大平房地区九佛堂组(热河群上段)的小盗龙亚科的新标本,该标本具有一些未曾报道过的有趣形态特征。尤其值得关注的是,一些特征呈现出介于中国鸟龙和小盗龙之间的过渡状态,其中部分来自牙齿和坐骨。这些特征突出显示了在更接近典型驰龙类的中国鸟龙和具有许多伤齿龙科特征的小盗龙之间存在着一系列的形态变异。然而,这些变异特征在分类上的意义尚未得到充分评估。特别值得一提的是该标本中相互愈合的耻骨和肠骨,这一特征有助于阐明复合骨在兽脚类恐龙中的演化模式。初步分析显示许多鸟类的复合骨都是通过在过型形成过程中各组分的顺序愈合形成的。
下白垩统,热河群,兽脚亚目,小盗龙亚科,变异,复合骨
Q915.864
A
1000-3118(2016)04-0269-17
2015-08-28
Xu X, Li F, 2016. A new microraptorine specimen (Theropoda: Dromaeosauridae) with a brief comment on the evolution of compound bones in theropods. Vertebrata PalAsiatica, 54(4): 269-285
国家自然科学基金(批准号:41120124002)和国家重点基础研究发展计划项目(编号:2012CB821900)资助。
猜你喜欢
文萃报·周二版(2022年15期)2022-04-13 15:24:16
金秋(2021年22期)2021-12-02 03:26:33
现代口腔医学杂志(2020年3期)2020-06-17 08:12:20
武夷科学(2019年2期)2019-12-20 08:39:16
中国眼耳鼻喉科杂志(2018年2期)2018-01-16 12:05:27
中国眼耳鼻喉科杂志(2018年2期)2018-01-16 12:05:27
武夷科学(2017年0期)2017-03-10 10:03:31
中国医疗美容(2015年4期)2015-04-27 02:24:05
黑龙江医药科学(2010年4期)2010-04-08 06:58:17
中国美容医学(2004年1期)2004-08-11 00:02:20

- 古脊椎动物学报(中英文)的其它文章
- On the geological age of mammalian fossils from Shanmacheng, Gansu Province
- 陕西洛南龙牙洞小哺乳动物化石新材料
- The first discovery of Urmiatherium (Bovidae, Artiodactyla) from Liushu Formation, Linxia Basin
- A skull of Machairodus horribilis and new evidence for gigantism as a mode of mosaic evolution in machairodonts (Felidae, Carnivora)
- Sciurid remains from the Late Cenozoic fi ssure- fi llings of Fanchang, Anhui, China