
5种地黄花部综合特征与繁育系统的初步研究
2016-03-24 06:32:51程永琴赵建花曾淑云刘占林
西北植物学报 2016年2期
关键词:花粉
程永琴,赵建花,韩 凯,曾淑云,黄 静,刘占林
(西北大学 生命科学学院,西部资源生物与现代生物技术教育部重点实验室,西安 710069)
5种地黄花部综合特征与繁育系统的初步研究
程永琴,赵建花,韩凯,曾淑云,黄静,刘占林*
(西北大学 生命科学学院,西部资源生物与现代生物技术教育部重点实验室,西安 710069)
摘要:生殖隔离是物种分化与延续的基础,地黄属植物是否存在生殖隔离仍不清楚。通过实地观测地黄属5个物种(天目地黄、湖北地黄、裂叶地黄、地黄和茄叶地黄)自然种群的花部形态、开花动态、花蜜体积、花蜜的糖浓度及其日变化、杂交指数、花粉-胚珠比等指标,结合人工授粉套袋,对地黄属植物的花部综合特征和繁育系统进行研究。结果显示:(1)地黄属植物的花为两性花,单花花期5~7 d,种群花期40~60 d;5种地黄在花序类型、花形态、花粉量、花粉-胚珠比等方面都存在差异,其中茄叶地黄的花粉完全败育。(2)5种地黄属植物的繁育系统属于异交,部分自交亲和,需要传粉者,不具有无融合生殖特性,地黄与茄叶地黄存在强烈营养繁殖。(3)5种地黄属植物的花朵基部有蜜腺可以产生糖浓度较高的花蜜,这也支持传粉者的存在,但除地黄确定为蜜蜂传粉外,其它地黄植物在野外并未观测到有效的传粉者,可能与地黄的散生特性、生境相互隔离以及传粉观测期气象条件恶劣有关。研究认为,5种地黄属植物较长的花期能有效弥补传粉者访问频率低的不足;除遗传屏障外,推测地黄属5个物种花展示方式的差异可能吸引不同的传粉者,加之地理、生态因素的作用导致了5种地黄属植物繁育系统的分化。
关键词:开花特性;繁育系统;花粉-胚珠比;杂交指数
地黄属(Rehmannia)是东亚分布物种,隶属玄参科。该属植物现有5种:天目地黄(R.chingii)、裂叶地黄(R.piasezkii)、湖北地黄(R.henryi)、地黄(R.glutinosa)和茄叶地黄(R.solanifolia)[1-2],为多年生草本,具根茎,总状花序,花萼钟状,萼齿5枚,花冠呈管状,分为5枚裂片(个别种内出现6裂片)。除茄叶地黄外其余4种地黄的雄蕊为4枚,茄叶地黄雄蕊多为6枚。种子多数,较小。
地黄属植物除地黄广泛分布于华北外,其它4种分布较为局限:天目地黄分布于浙江、安徽的交界处,裂叶地黄、湖北地黄和茄叶地黄分布于秦巴山区。近年来对地黄属的研究多偏重于地黄的药性、化学成分及品种间遗传基础和系统发育关系等方面[3-6],而对其繁育系统研究较少,仅有怀地黄的花粉形态[7],地黄的繁育系统的研究[8],其它4种的繁育系统研究未见报道。繁育系统的隔离是物种维持与分化的基础,花作为被子植物的繁殖器官,其开放样式和花部设计吸引不同的传粉者[9],再加上地理隔离、生态因素等可能导致不同物种繁育系统的分化。地黄属不同种类是否存在物种特有的花部特征、传粉昆虫以及遗传隔离机制等都是亟待解决的问题。本研究以5种地黄为研究对象,研究地黄属植物自然居群的开花动态、花部形态、杂交指数、花粉-胚珠比、花蜜体积、花蜜的糖浓度,结合授粉和套袋的结果,揭示地黄属植物的花部特征和开花规律,初步确定其繁育系统,判断各种间是否存在遗传隔离,为地黄属植物生殖生物学以及物种形成和分化的深入研究奠定基础。
1材料和方法
1.1研究地自然概况
我们于2013年4~7月选择5种地黄个体较多并且人为干扰较少的自然居群进行观察,观测地分别是:天目地黄——浙江省安吉县(119.55°E,30.53°N),裂叶地黄——湖北省谷城县(111.54°E,32.11°N),湖北地黄——湖北省兴山县(110.63°E,31.36°N),地黄——陕西省兴平市(108.45°E,34.32°N),茄叶地黄——重庆市城口县(108.80°E,31.73°N)。观测后将这5个地点的5种地黄植株移栽于西北大学温室,在相同的环境下培育,于2014年4~7月进行杂交实验的研究。
1.2研究方法
1.2.1开花物候和花部特征的观察在研究地点标记、观测进入生殖生长期植株,每种地黄选择10株,每株标记3朵花。观察并记录其开花进程,以及花被、花丝、花药、子房的颜色、形状并测量花萼长宽,花瓣长宽,花冠直径,花冠管长度,花药和花丝的长宽,子房和花柱的长宽,柱头与花药的空间距离[10-11]。同时我们在花冠的纵轴和横轴测量花冠的张开度,用其比值表示花冠的圆度(比值为1∶1说明花冠张开度是圆形)。
1.2.2花粉败育率的测定参考陈家瑞的孢粉染色法[12]。
1.2.3泌蜜动态测定选择地黄属植物盛花期,测量地黄属植物的花蜜量和花蜜的糖浓度。选取5个植株,取每株植株上刚要开花的2朵花进行标记,然后套袋,对所有标记花的花蜜量进行测量,在8:00~18:00每隔2 h用10 μL的毛细管吸取花蜜1次;如果糖量足够多,用手持糖量计折射仪测定花蜜的糖浓度。
花蜜体积=(毛细管中液柱长/毛细管总长)×毛细管的总体积[13]
1.2.4繁育系统类型的判断(1)单花花粉量及P/O值的测定参考Dafni等[14]的方法,选取20朵始花期、未开裂的花测每朵花的花粉数。将每朵花的子房直接在载玻片上压开,统计其胚珠数。
P/O值=单花花粉数/单花胚珠数
(2)去雄-套袋-授粉实验检测繁育系统类型 参考Dafni等[14]和Bowers[15]的描述方法,在实验地选取位置相对一致的花,在花蕾期挂牌,分别进行套袋、去雄及人工授粉实验,具体的处理参照刘林德等的方法[16]。
(3)人工杂交实验杂交授粉实验于2015年4月1日~5月30日进行,选取种植于温室的5种地黄进行正反交,每个品种选3~4株生长健康的植株作为母本。由于5种地黄的始花期时间不同,但是他们的花期有重叠,在花期重叠的时间内,将其它4种地黄的花粉用毛笔和镊子涂抹到另外一种地黄的柱头上进行人工授粉,授粉后套袋,待种子成熟后记录结实情况,记录种子萌发率。
(4)OCI指数的估算按照Dafni的标准[17]进行花序直径、花朵大小及开花行为的测量及繁育系统的评判。
1.2.5访花昆虫的调查在花期,随机标记5株已开放的地黄属植株,每株标记4个花,从日出到日落(约7:30~20:00)进行连续观察记录访花昆虫的种类并照相,捕捉访花昆虫,毒瓶杀死阴干,用于鉴定。
1.2.6数据分析方法用Statistica 6.0处理数据,图表在Origin 9.0下完成。
2结果与分析
2.15种地黄的开花物候期和花部特征
野外观察发现5种地黄的开花时间不同,天目地黄于3月底开花,湖北地黄和地黄的开花时间在4月上旬,茄叶地黄于4月中旬开花,裂叶地黄的开花时间比其它4种地黄晚,于5月份开花。5种地黄的单花花期为5~7 d,野外观察5种地黄属植物的花期显示,茄叶地黄花期最短,为40 d左右,地黄花期为50 d左右,天目地黄、湖北地黄、裂叶地黄的花期都在60 d左右。
地黄属植物花均为两性的管状花,花朵两侧对称,花瓣上唇2枚下唇3枚。花蕾多在中午前后开放,大多数地黄的花在开花当天就产生花蜜,花开放时,柱头明显高于花药的位置,花柱在花期内伸长不明显,而花丝在开花当天或第2天开始伸长,直到花药开裂花粉散出。地黄属的花中除茄叶地黄为6个花丝外,其余地黄均为4个花丝,在这4个花丝中一般2枚较长的花丝先伸长且花药开裂散粉,其余2枚滞后1~2 d。花柱在花朵内部直立生长,花丝弯曲生长,花药紧贴花管内部,花凋谢时花瓣和雄蕊一起脱落。5种地黄花朵的功能形态特征如表1所示,裂叶地黄的植株明显高于其它4种地黄。地黄属植物花朵开放时初始花蕾颜色为淡黄或者淡粉,等花朵开放,5种地黄的花瓣呈现不同的颜色,天目地黄的花瓣为紫红色,裂叶地黄呈粉红色,湖北地黄为白色,地黄呈淡黄色,茄叶地黄呈淡紫色。通过我们观测发现虽然5种地黄的花均为管状,但其花朵基部张开角度(花朵基部的张开角度是以花中花柱为中心轴,花冠底部与花柱所呈现的角度大小)却不同,湖北地黄花朵基部的张开角度最大,且其花冠的圆度最接近1∶1。通过观察花朵内部结构发现5种地黄中只有裂叶地黄的花药位置比柱头高,这样的结构方便花粉自然落到柱头上;5种地黄的柱头形状也有所不同,地黄和茄叶地黄的柱头呈圆形,其它3种地黄的柱头呈两片状。用孢粉染色法观察5种地黄的花粉败育,发现茄叶地黄的花粉败育率为100%,裂叶地黄的花粉败育率为76%,地黄的花粉败育率为53%,天目地黄的花粉败育率为39%,湖北地黄的花粉败育率为38%。

表1 5种地黄花朵的功能形态特征
注:同行不同小写字母表示差异显著(P<0.05)。
Note:Different normal letters in the same row mean significant difference(P<0.05).
2.2花蜜量的测定
地黄属植物的花在开放过程中分泌有花蜜,其蜜腺位于花冠筒和花柱基部,由于茄叶地黄观测点环境恶劣,花朵受风雨打击较大,花朵基部多存有雨水,因此没有测其花蜜量。对4种地黄的花蜜量和糖浓度测定结果显示,泌蜜量(图1,A)和糖浓度(图1,B)变化趋势相似,从8:00时起逐渐升高,12:00时达到最高值,随后逐渐下降。从图1中我们可以看出湖北地黄和地黄的花蜜量最多,但是从测定的糖浓度看,地黄的花蜜糖浓度却最低。
2.3繁育系统观测结果
2.3.1单花花粉量及花粉-胚珠比不同植物的花粉-胚珠比(P/O值)可以反映植物的不同繁育系统类型,根据Cruden P/O值和繁育系统的对应关系[18],如表2所示,天目地黄的P/O值为220,湖北地黄的P/O值为252,介于31.9~396.0之间,繁育系统属于兼性自交类型,裂叶地黄P/O值为456,地黄的P/O值为400,介于244.7~2 588.0之间,繁育系统属于兼性异交。由于茄叶地黄花粉完全败育,故没有测其P/O值。
2.3.2套袋-去雄-授粉的实验结果(1)繁育系统检测结果用不同的授粉处理进一步确定地黄属植物的繁育系统类型。表3授粉结实数据表明,去雄套袋处理后,均未见结实,说明地黄属植物不存在无融合生殖;天目地黄、湖北地黄和地黄的自花授粉不结实,而裂叶地黄自花授粉可以结实。自然条件下的传粉和人工异株异花授粉的结实率较高,套纸袋与套网袋后结实率较低,表明套袋隔离访花者后,传粉成功率降低。
(2)人工杂交实验结果5种地黄杂交实验结果如表4所示,茄叶地黄杂交均不结实,在其它杂交组合中除天目地黄与地黄的正交、地黄与裂叶地黄的正交无结实外其余组合均有结实。茄叶地黄花粉败育故无法授粉给其它种地黄,然而我们将其它种地黄的花粉授予茄叶地黄时发现不结实。为了进一步验证结实情况,我们种植杂交结实的种子,统计其出苗率,结果显示:天目地黄×湖北地黄的正交和反交种子出苗率为100%;天目地黄×裂叶地黄的正交产生的种子出苗率为100%,反交产生的种子出苗率仅为15%;湖北地黄×裂叶地黄的正交产生的种子出苗率为100%,反交产生的种子出苗率为30%。
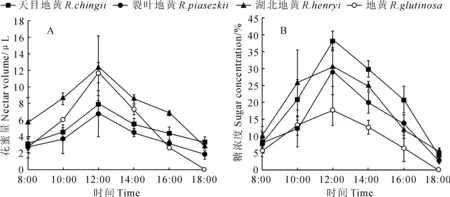
图1 地黄属植物的花蜜量(A)和糖浓度(B)

测量项目Itemofobservation天目地黄R.chingii裂叶地黄R.piasezkii湖北地黄R.henryi地黄R.glutinosa茄叶地黄R.solanifolia每花花粉数量Pollengrainnumberperflower88000±2000b489600±13948a307640±25733a160700±14849b-每花胚珠数目Ovulenumberperflower400±50b1074±63a1293±399a402±45b-花粉胚珠比P/O220±30b456±34a252±72b400±9a-繁育系统类型TypeofbreedingsystemFAFXFAFX-
注:FA.兼性自交;FX.兼性异交;同行不同小写字母表示差异显著(P<0.05);- 表示该操作不能进行。
Note:FA.Facultative autogamy;FX.Facultative xenogamy;Different normal letters in the same row mean significant difference(P<0.05);- means this test cannot be carried out.

表3 5种地黄去雄、套袋及人工授粉实验的结果
注:0表示不座果;+表示座果;-表示该操作不能进行。
Note:0 means no fruit setting;+ means setting some fruits;- means this test cannot be carried out.
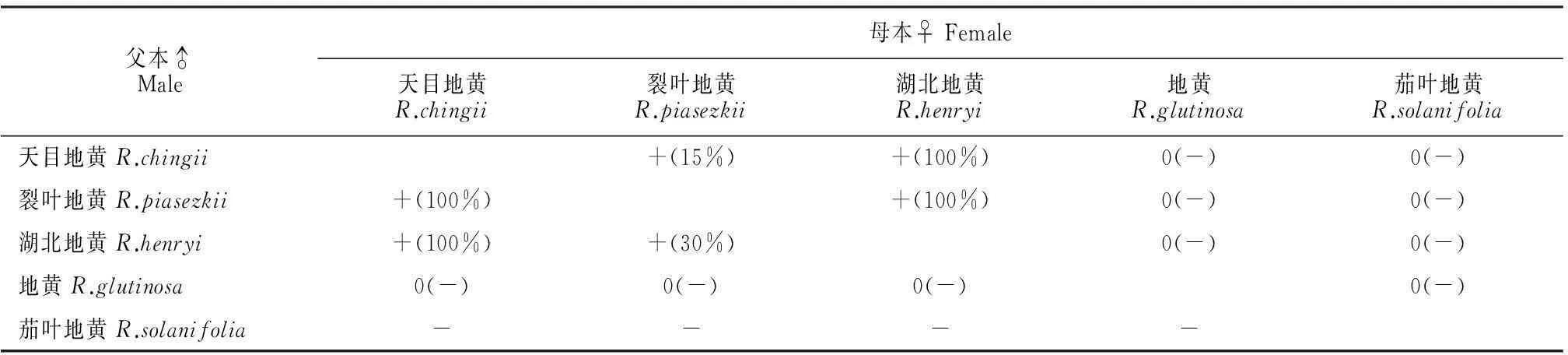
表4 地黄属植物种间杂交结实与出苗率统计
注:0表示不座果;+表示座果;-表示该操作不能进行,括号内数据表示出苗率。
Note:0 means no fruit setting;+ means setting some fruits;- means this test cannot be carried out.The data in the bracket represent seedling rates.

表5 5种地黄杂交指数观测结果
2.3.3OCI指数的估算按照Dafni方法[17]的测量结果如表5,地黄和茄叶地黄的花朵直径虽然较其它3种地黄小,但是直径均大于6 mm,所以均记为3;5种地黄均为雄蕊先熟,所以记为1;而柱头和花药之间也均存在空间分隔,记为1。OCI的结果显示5种地黄的繁育系统类型均为异交,部分自交亲和,需要传粉者。
2.4传粉昆虫
对5种地黄的传粉情况观察表明,访花昆虫极少,在天目地黄花朵内观察到蓟马,在湖北地黄中发现有寄生的蜗牛和蚊蝇类昆虫,在裂叶地黄中观察到蝴蝶停留在花朵上,这些昆虫是否是有效传粉昆虫,有待于进一步确定。在地黄中发现蜂类吸取花蜜。这些昆虫的访花时间都较短,访花时间集中在9:00~16:00,16:00过后访花昆虫活动减少。
3结论与讨论
3.1地黄属的开花特性
本研究野外观测到5种地黄的始花期有些差异,这可能与分布区的气候条件和物种间差异有关,但当观察移栽于同一地点的5种地黄,发现这5种地黄的始花期依然不同,说明地黄属不同物种对光和环境的适应与调节机制不同,这些差异使得移栽于同一地点的不同物种的开花时间不同,开花时间上的不同以及地理隔离,阻止了不同物种之间的基因交流(花粉流),维持了物种的特异性和稳定性。
3.2地黄的繁育系统
据Danfi的标准[17]所得出的杂交指数表明,地黄属植物的繁育系统为异交,部分自交亲和,需要传粉者。按照Cruden的花粉胚珠比(P/O)的标准[18]划分,天目地黄和湖北地黄的繁育系统属于兼性自交类型,裂叶地黄和地黄的繁育系统类型属于兼性异交类型。套袋实验结果显示,地黄属植物不存在无融合生殖,只有裂叶地黄自花授粉可以结实,自然条件下的传粉和人工异株异花授粉的结实率相对较高。由此推测,该属物种可能属异花授粉。
天目地黄和湖北地黄是姐妹类群[4],它们的花部特征和繁育系统很相似,植株高度、柱头形状、花粉败育率、P/O值等都没有显著差异。两个物种最明显的区别在于花朵颜色,天目地黄的花呈玫红色,湖北地黄的花呈白色。野外观察发现,湖北地黄和天目地黄的花药均低于柱头,如果不借助风媒和虫媒传粉,花粉很难到达柱头,这种柱头和花药之间的隔离有利于异花授粉,而且雌雄蕊异熟也是它们不能自花授粉的一个重要原因。因此推测天目地黄和湖北地黄的繁育系统类型为异交,需要传粉者。
裂叶地黄的始花期较晚,花朵大,花期长,花朵颜色鲜艳,在裂叶地黄中发现其花丝长度高于花柱或者与花柱等高,这使得花粉很容易落在柱头上,套袋-去雄-授粉的实验结果也表明裂叶地黄自花授粉后会结实,这说明裂叶地黄自交亲和,而异交的结实率更高。在气候等条件不利时,裂叶地黄可以自交产生种子以繁殖后代。这种多样的繁殖方式对裂叶地黄的繁殖和物种的延续都更有保障。
地黄和茄叶地黄的花较其它几种地黄花朵小,颜色不鲜艳。根据花部特征和对繁育系统的验证,地黄属于少部分自交亲和,一般为异交,需要传粉者。在野外我们也观察到了蜜蜂对地黄花的访问,这与张猛猛[8]的研究结果一致。茄叶地黄的花期最短,且花粉完全败育,我们根据花部特征以及茄叶地黄的“成堆式”分布,推测茄叶地黄是用块根进行营养繁殖。地黄也存在营养繁殖现象[19]。
地黄属植物的花粉数均显著多于胚珠数,但是其花粉败育率也高,以及雌雄蕊异熟,使得自交结实成功的概率并不大。植物自交产生的后代一般对环境的抵抗力差,但是在缺乏异交的条件以及环境的压力下,逐渐形成了异交为主、自交为辅的方式,自交亲和降低了生殖失败的风险,保障了后代的延续和物种的存活。
3.3传粉者的推测
对于自交不亲和的植物而言,传粉者对于植物繁殖的重要性不言而喻。花朵通过鲜艳的颜色、便于传粉者停留的花部结构以及提供花蜜花粉等报酬吸引不同的传粉昆虫,增加传粉机会进而提高传粉成功的概率,为植物后代的繁衍和物种的延续奠定基础。5种地黄的花颜色多样,不同颜色的花以及不同的花香会吸引不同类型的传粉昆虫[20]。花特征的演变与传粉者之间的变化有很大关系,尤其是花管长度的变化[21-22],地黄属植物花冠管的基部会产生糖浓度很高的花蜜,传粉者或者采蜜的昆虫想要食得花蜜必须有和管状花相适应的长舌结构等,然而野外实验并未观察到多种类的昆虫访问,这可能和观测期当地长期阴雨的天气相关,也可能是授粉昆虫没有与花部特征相适应的结构,吸吮不到管状花的花蜜,久而久之采蜜和授粉的昆虫转为其他更容易获得传粉报酬的植物传粉。在野外观察发现地黄属的5个物种并没有重叠分布,各自占据不同的生态位,再结合花部特征分析,推测地黄属的植物可能吸引的是不同的传粉者。
3.4地黄繁殖隔离的因素分析
繁殖隔离是物种形成和分化的基础,对于异域分布物种地理隔离减弱或者阻止了基因流,逐渐形成生殖隔离[23]。而同域分布的植物多以物候隔离作为生殖隔离的主要机制[24-26]。5种地黄中,地黄和茄叶地黄是四倍体物种,其它地黄属于二倍体[4],倍性差异导致的生殖屏障是繁殖隔离的关键因素。湖北地黄和裂叶地黄分布于秦巴山区,属于同域分布物种,但二者开花时间分别为4月上旬和5月份,花期为60 d,虽然花期有重叠,但没有相同的传粉昆虫在他们之间充当基因交流的桥梁,使得它们之间的基因交流不能正常进行,而花色不同导致的传粉者差异可能加剧了物种合子前隔离。天目地黄分布于华东地区,与湖北地黄的华中分布不同,地理隔离是基因交流主要障碍。人工杂交的实验结果,也验证了这一点:二倍体的天目地黄、湖北地黄和裂叶地黄之间并没有遗传上的生殖屏障,只是地理和生态隔离使得它们之间不能进行基因交流。对于同是四倍体的地黄和茄叶地黄而言,茄叶地黄分布区较地黄偏南,而且海拔更高(1 200 m vs 400 m),地理和生态隔离可能同时起到重要作用,而地黄和茄叶地黄的杂交并不结实,这说明四倍体的地黄和茄叶地黄可能在遗传机制上有所差别导致生殖隔离。综合上述分析,我们认为多种因素导致的生殖隔离是自然条件下地黄属物种稳定与维系的关键。需要指出的是,地黄属植物均为异交为主的繁育系统,地理、生态或者遗传因素究竟对种间生殖隔离的影响程度如何,仍需要进一步研究。
参考文献:
[1]李晓东,昝艳燕,罗曼曼,等.地黄属的分类学修订[J].植物科学学报,2011,29(4):423-431.
LI X D,ZAN Y Y,LUO M M,etal.Taxonomic revision of the genusRehmannia[J].PlantScienceJournal,2011,29(4):423-431.
[2]李宏庆,刘国丽.地黄属的一个新异名[J].广西植物,2012,32(1):23-26.
LI H Q,LIU G L.A new synonym ofRehmannia(Scrophulariaceae s.l.)[J].Guihaia,2012,32(1):23-26.
[3]肖玲,赵先贵,常思明.地黄块根的发育解剖学研究[J].西北植物学报,1996,16(5):44-47.
XIAO L,ZHAO X G,CHANG S M.Developmental anatomy of the root tuber ofRehmanniaglutinosa[J].ActaBot.Boreal.-Occident.Sin.,1996,16(5):44-47.
[4]ALBACH D C,LI H Q,ZHAO N.Molecular systematics and phytochemistry ofRehmannia(Scrophulariaceae)[J].BiochemicalSystematics&Ecology,2007,35(5):293-300.
[5]闫坤,赵楠,李宏庆.地黄属种间亲缘关系研究[J].西北植物学报,2007,27(6):1 112-1 120.
YAN K,ZHAO N,LI H Q.Systematic relationships amongRehmannia(Scrophulariaceae) species[J].ActaBot.Boreal.-Occident.Sin.,2007,27(6):1 112-1 120.
[6]SUN P,GUO Y H,QI J J,etal.Isolation and expression analysis of tuberous root development related genes inRehmanniaglutinosa[J].MolecularBiologyReports,2010,37(2):1 069-1 079.
[7]李建军,王莹,朱命炜,等.怀地黄不同主栽品种花粉形态比较[J].河南农业大学学报,2012,46(2):127-130.
LI J J,WANG Y,ZHU M W,etal.Observation and comparison of morphological characteristics of different cultivated varieties ofRehmanniaglutinosa[J].JournalofHenanAgriculturalUniversity,2012,46(2):127-130.
[8]张猛猛,王奎玲,刘庆华,等.地黄开花生物学特性研究[C]//中国观赏园艺研究进展.山东青岛,2014:258-262.
[9]BARRETT S C,HARDER L D.Ecology and evolution of plant mating[J].TrendsinEcology&Evolution,1996,11(2):73-79.
[10]OLESEN J M.Floral morphology and pollen flow in the heterostylous speciesPulmonariaobscuraDumort (Boraginaceae)[J].NewPhytologist,1979,82(3):757-767.
[11]黄双全,郭友好.传粉生物学研究进展[J].科学通报,2000,45(3):225-237.
HUANG S Q,GUO Y H.New advances in pollination biology[J].ChineseScienceBulletin,2000,45(3):225-237.
[12]陈家瑞.植物孢粉染色技术综述及其应用[J].植物学集刊,1991,5:259-276.
CHEN J R.Technique of staining spore and pollen and its application for botanical and agronomic studies[J].BotanicalResearch,1991,(5):259-276.
[13]CRUDEN R W,HERMANN S M.Studying Nectar?Some Observations on the Art[M]//BENTLEY B L,ELIAS T.The Biology of Nectaries.New York:Columbia University Press,1983:223-241.
[14]DAFNI A,KEVAN P G,HUSBAND B C.Practical Pollination Biology[M].Canada:Enviroquest Ltd,2005:39-64.
[15]BOWERS K AW.The pollination ecology ofSolanumrostratum(Solanaceae)[J].AmericanJournalofBotany,1975,62(6):633-638.
[16]刘林德,祝宁,申家恒,等.刺五加、短梗五加的开花动态及繁育系统的比较研究[J].生态学报,2002,22(7):1 041-1 048.
LIU L D,ZHU N,SHEN J H,etal.Comparative studies on floral dynamics and breeding system betweenEleutherococcussenticosusandE.sessiliflorus[J].ActaEcologicaSinica,2002,22(7):1 041-1 048.
[17]DAFNI A.Pollination Ecology:a Practical Approach[M].Oxford:Oxford University Press,1992:1-57.
[18]CRUDEN R W.Pollen-ovule rations:a conservative indicator of breeding systems in flowering plants[J].Evolution,1977,31(1):32-46.
[19]温学森,杨世林,魏建和,等.地黄栽培历史及其品种考证[J].中草药,2002,33(10):946-949.
WEN X S,YANG S L,WEI J H,etal.Textual research on planting history ofRehmanniaglutinosaand its cultivated varieties[J].ChineseTraditionalandHerbalDrugs,2002,33(10):946-949.
[20]PEAKALL R,WHITEHEAD M R.Floral odour chemistry defines species boundaries and underpins strong reproductive isolation in sexually deceptive orchids[J].AnnalsofBotany,2014,113(2):341-355.
[21]NEWMAN E,MANNING J C,ANDERSON B.Matching floral and pollinator traits through guild convergence and pollinator ecotype formation[J].AnnalsofBotany,2014,113(2):373-384.
[22]NIET T V,PIRIE M D,SHUTTLEWORTH A,etal.Do pollinator distributions underlie the evolution of pollination ecotypes in the Cape shrubEricaplukenetii?[J].AnnalsofBotany,2014,113(2):301-315.
[23]SMADJA C M,BUTLIN R K.A framework for comparing processes of speciation in the presence of gene flow[J].MolecularEcology,2011,20(24):5 123-5 140.
[24]HUSBAND B C,SABARA H A.Reproductive isolation between autotetraploids and their diploid progenitors in fireweed,Chamerionangustifolium(Onagraceae)[J].NewPhytologist,2004,161(3):703-713.
[25]HUSBAND B C,SCHEMSKE D W.Ecological mechanisms of reproductive isolation between diploid and tetraploidChamerionangustifolium[J].JournalofEcology,2000,88(4):689-701.
[26]LAMONT B B,HE T,ENRIGHT N J,etal.Anthropogenic disturbance promotes hybridization betweenBanksiaspecies by altering their biology[J].JournalofEvolutionaryBiology,2003,16(4):551-557.
(编辑:潘新社)
Preliminary Study on Floral Syndroma and Breeding System for Five Species ofRehmannia(Scrophulariaceae)
CHENG Yongqin,ZHAO Jianhua,HAN Kai,ZENG Shuyun,HUANG Jing,LIU Zhanlin*
(Key Laboratory of Resource Biology and Biotechnology in Western China,Ministry of Education,College of Life Sciences,Northwest University,Xi’an 710069,China)
Abstract:Reproduction isolation is the basis of species preservation and divergence,but little is known about the breeding system of Rehmannia.We studied the floral syndroma and reproductive characteristics of the five Rehmannia species (R.chingii,R.piasezkii,R.henryi,R.glutinosa,R.solanifolia) through investigating the following indices in field:flowering dynamics of natural populations,floral morphologies of flower organs,nectar volume and sugar concentration,out-crossing index,pollen-ovule ratio,pollination and bagging experiment.Our results indicate that:(1)Rehmannia species flowers are hermaphroditic.The flowering duration of a single florescence was 5-7 days,and the whole population flowering duration was 40-60 days.There are distinct difference among the five Rehmannia species in inflorescence types,floral characteristics,flower color,pollen amounts and P/O values.We found that R.solanifolia were pollen abortion.(2)The evidence of the out-crossing index,pollen-ovule ratio and hybridization experiment supported that the breeding system of Rehmannia was belonged to cross pollination system,partially self-compatible,with no apomictic ability,and pollinators required.We also found there was vegetative reproduction in Rehmannia,especially in R.glutinosa and R.solanifolia.(3)There are nectaries on the base of Rehmannia corolla and they can produce nectar with higher concentration of sugar,and this also supports the existence of pollinators.However,we did not observe any effective pollinators in Rehmannia species except in R.glutinosa with bees,which might be due to the scattered individual,isolated habitats and the bad weather conditions during pollination observation.Longer flowering duration could compensate for the insufficiency of pollinator visits.According to our results,we suppose that the different flowers display of five Rehmannia species possibly attracting distinct pollinators,together with genetic,geographic and ecological isolation,results in the differentiation of the breeding system in Rehmannia species.
Key words:flowering characteristics;breeding system;pollen-ovule ratio;outcrossing index
中图分类号:Q944.4
文献标志码:A
作者简介:程永琴(1990-),女,在读硕士研究生,主要从事植物繁殖生态学研究。E-mail:courage4132231@163.com*通信作者:刘占林,教授,主要从事进化生物学研究。E-mail:liuzl@nwu.edu.cn
基金项目:国家自然科学基金项目(31370353);陕西省自然科学基础研究计划(2015JM3106)
收稿日期:2015-10-30;修改稿收到日期:2015-12-14
文章编号:1000-4025(2016)02-0404-07
doi:10.7606/j.issn.1000-4025.2016.02.0404
猜你喜欢
奥秘(创新大赛)(2023年5期)2023-09-02 03:15:18
儿童故事画报·自然探秘(2023年4期)2023-03-15 16:58:57
哈哈画报(2022年8期)2022-11-23 06:21:20
好孩子画报(2022年4期)2022-06-08 23:10:48
中国生殖健康(2019年3期)2019-02-01 06:12:22
中国蜂业(2018年6期)2018-01-21 15:44:41
中国果业信息(2018年8期)2018-01-17 23:49:08
儿童故事画报·发现号趣味百科(2017年4期)2017-06-30 08:05:59
文理导航·科普童话(2016年6期)2016-09-12 23:24:08
江西理工大学学报(2015年3期)2015-12-22 05:26:25
