
乌鲁木齐河源区12种藓类植物叶尖的微形态结构
2016-03-24 06:32艾菲热阿布都艾尼
西北植物学报 2016年2期
王 虹,艾菲热·阿布都艾尼
(新疆大学 生命科学与技术学院,乌鲁木齐 830046)
乌鲁木齐河源区12种藓类植物叶尖的微形态结构
王虹,艾菲热·阿布都艾尼
(新疆大学 生命科学与技术学院,乌鲁木齐 830046)
摘要:对乌鲁木齐河源区生长的12种藓类植物叶尖的顶端细胞,以及与其相邻的其他叶尖细胞表面的小孔、乳突、角质层纹饰等微结构,进行光学显微镜和扫描电镜观察,为高山冻原区苔藓植物的结构、分类及生理生态学研究提供理论依据。结果表明:大多数藓类植物叶尖的顶端细胞透明,但形态各异,细胞比叶片细胞长,干时细胞壁有的收缩凹陷,有的并不收缩,细胞壁表面有不规则细纹及细小的颗粒;叶尖在干时边缘内卷或拧卷、细胞壁厚、表面有较多的小孔,这些特征有利于水分的吸收及抵御长期寒冷、反射太阳辐射对其伤害而适应环境所形成的生存策略。叶尖的类型对苔藓植物科、属级的分类意义不大,但其微形态如叶尖顶端细胞及其他细胞的形状、数目、细胞壁的凹陷程度以及其上角质层纹饰、乳突、粗疣及小孔等微形态结构特征,在同属的不同种之间存在明显差异,具有一定的分类学价值。
关键词:乌鲁木齐河源区;藓类植物;扫描电镜;叶尖;微形态结构
乌鲁木齐河源区,在有“冰川活化石”之誉的新疆天山一号冰川地区,该地区海拔高,气温低,降水分配不均,年降水量431.2 mm,年平均气温-5.5 ℃,气候寒冷、光照强,生长着耐寒、耐旱、低矮、垫状的高山草甸植物,具典型高山冻原植被带特征[1]。
高山冻原植被——主要是苔藓植物[2-5],其矮小、垫状、密集丛生等特性,具有很强的涵养水源的功能,也成为这一地区的优势类群,对维护高山冻原带的生态平衡起到了不可替代的作用,许多苔藓植物具有很强的耐寒、耐旱等抗逆性状,也使得多种苔藓植物在恶劣的环境中成为先锋植物[6-7]。对新疆天山的苔藓植物的研究工作,有赵建成、买买提明等对新疆东部、博格达地区以及新疆天山的苔藓植物进行区系分类的研究[8-10];张元明等对新疆博格达山地面生苔藓植物及三工河流域的苔藓植物物种多样性及其分布做了详细的调查[11-12];王虹、姜彦成等分别对一号冰川地区4种藓类植物及紫萼藓科的5种植物做了解剖学研究,讨论了它们茎及叶的结构与环境的适应性[13-14];曾健等也对新疆天山一号冰川10种藓类植物的叶尖(毛尖)微形态做了专门的研究,指出其对于苔藓植物属下种间的鉴定具有一定的分类学意义[15]。本实验对长期生长在高寒、干旱等极端环境下的苔藓植物叶尖的微形态结构的研究,以便为以后的学者研究高山冻原区苔藓植物的结构、分类及生理生态学等方面提供一些理论依据。
1材料和方法
本实验的12种藓类植物标本均采自乌鲁木齐河源区的新疆天山一号冰川附近,海拔高度在3 522~3 835 m的岩面、土面或水边(表1),所有的标本均存放于HTC及XJU。
分别将干标本放入盛水的培养皿中,用尖头镊子把植物体基部的泥土进行初步的清理,然后再用毛笔刷洗,最后用Polaron CA7615型超声波清洗仪洗2次,每次30 min。将每种不少于10个植株的标本,在解剖镜下取中部叶片置于载玻片上,再在Olympus显微镜下详细观察至少20个样,并在Nikon光学显微镜下照相。取相同部位的叶片不少于6片,分别经酒精梯度脱水,干燥后每种分背、腹面粘于样品台上,在IB-5离子溅射仪中真空喷镀膜15 min,并用LEO 1430VP型扫描电镜进行观察并拍照。
2观察结果
2.1光学显微镜下叶尖的形态特征
依据胡人亮将藓类植物叶先端形态划分成12种类型[2],本实验将乌鲁木齐河源区的12种藓类植物的叶尖形态特征(表2)分为以下5种类型:(1)先端渐尖,如斜蒴对叶藓、辛氏曲柄藓、东亚泽藓、中华细枝藓、牛角藓原变种、仰叶拟细湿藓、(漆光)镰刀藓(图版Ⅰ,1、2、6、9~12);(2)先端锥形,如葫芦藓、平肋提灯藓(图版Ⅰ,3、4);(3)先端锐,急尖,如全缘匍灯藓(图版Ⅰ,5);(4)先端圆钝,如(钝叶)小鼠尾藓(图版Ⅰ,7);(5)先端具芒,如柔叶(细枝)小鼠尾藓(图版Ⅰ,8)。观察发现,有的(钝叶)小鼠尾藓的叶片也有小芒尖。
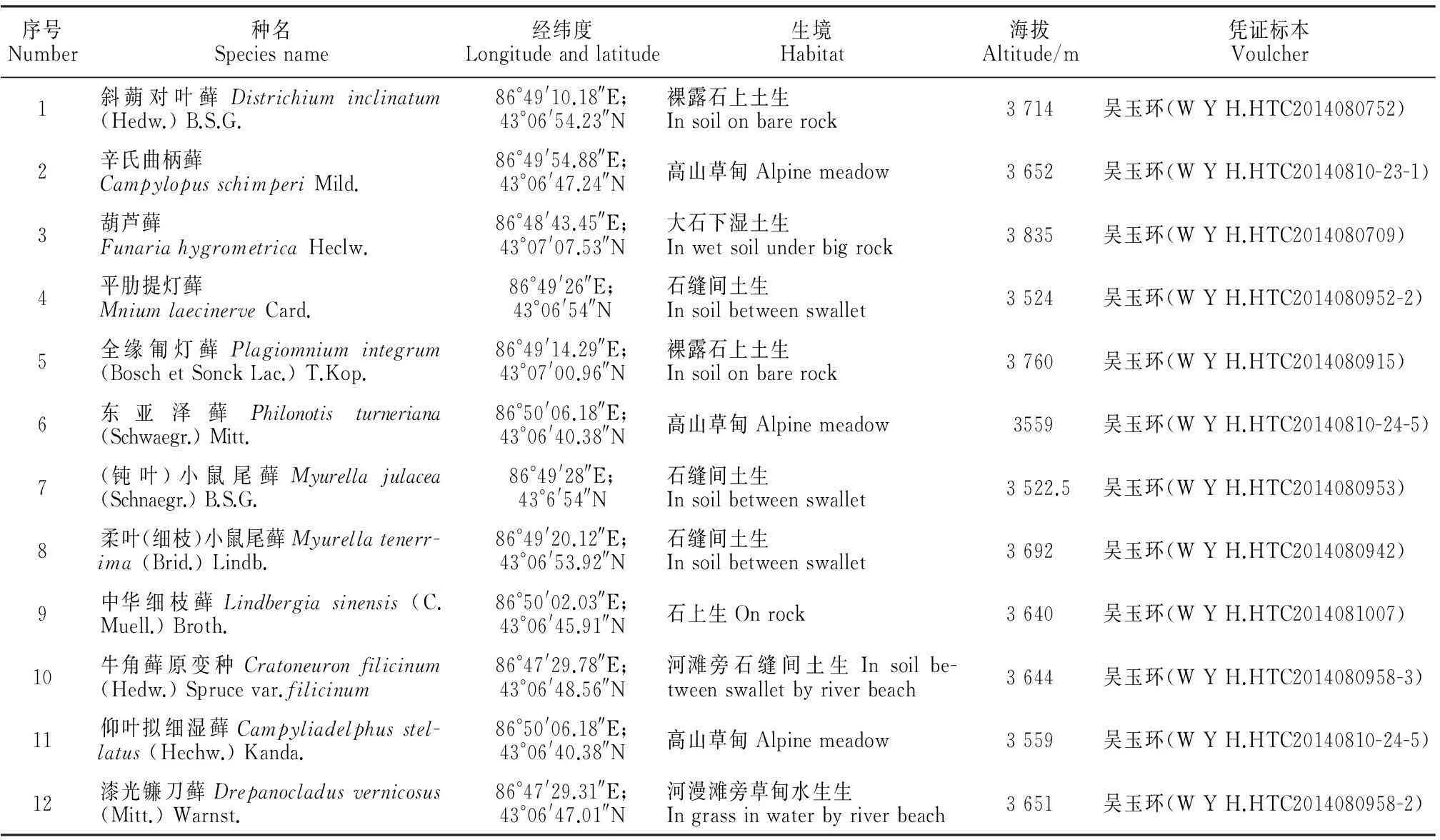
表1 实验材料及来源

2.2扫描电镜下叶尖的顶端细胞形态特征
扫描电镜下叶尖干时:(1)先端圆锥形,具多个小尖头,细胞不收缩。如斜蒴对叶藓(图版 Ⅰ,14、15;图版 Ⅱ,1);(2)先端呈三棱形渐尖,细胞收缩不明显。如辛氏曲柄藓、葫芦藓(图版 Ⅱ,2~7);(3)先端急尖,细胞不收缩。如平肋提灯藓、全缘匍灯藓(图版 Ⅱ,8~12);(4)先端扁平,细胞上下收缩,边缘具的细胞壁在尾部凸起。如东亚泽藓、(漆光)镰刀藓(图版Ⅱ,13、14;图版Ⅲ,11~12);(5)中华细枝藓、牛角藓原变、仰叶拟细湿藓种(图版Ⅲ,4~10);(6)钝圆,如(钝叶)小鼠尾藓(图版Ⅱ,15;图版Ⅲ,1);(7)呈芒尖:如柔叶(细枝)小鼠尾藓(图版Ⅲ,2、3)。
2.3扫描电镜下叶尖顶端细胞表面的角质层纹饰
这12种藓类植物叶尖的顶端细胞表面的角质层纹饰,在组成、形态和分布的疏密程度分为以下7种:(1)具乳突的只有斜蒴对叶藓(图版Ⅰ,14、15);(2)具不规则纹饰(纵纹或细纵纹),如斜蒴对叶藓、全缘匍灯藓、东亚泽藓、中华细枝藓(图版Ⅰ,13~15;图版Ⅱ,11、13;图版Ⅲ,4);(3)具细小颗粒的,如辛氏曲柄藓、葫芦藓、(钝叶)小鼠尾藓、柔叶(细枝)小鼠尾藓、中华细枝藓、牛角藓原变种、 (漆光)镰刀藓(图版Ⅱ,2、5、15;图版Ⅲ,2、4、6、11);(4)具少量纹饰(或粗纹)的,如辛氏曲柄藓、(钝叶)小鼠尾藓(图版Ⅱ,2;图版Ⅲ,15);(5)具密集的纵纹(或细纹)的,如葫芦藓、平肋提灯藓、牛角藓原变种、仰叶拟细湿藓、 (漆光)镰刀藓(图版Ⅱ,5、8;图版Ⅲ,6、8、11);(6)具鳞片状纹饰的,如平肋提灯藓(图版Ⅱ,8);(7)具较多不规则纹饰(或细纵纹)的,如柔叶(细枝)小鼠尾藓(图版Ⅲ,2)。可以看出,有的种其植物叶尖的顶端细胞表面的角质层纹饰同时具有2种。
2.4扫描电镜下叶尖其他细胞的角质层纹饰
与叶尖顶端细胞相邻的其他细胞表面角质层纹饰:(1)具小孔,如斜蒴对叶藓、辛氏曲柄藓(图版Ⅰ,14、15;图版Ⅱ,2~4);(2)具细纵纹(或不规则纹饰),如斜蒴对叶藓、平肋提灯藓、东亚泽藓、(钝叶)小鼠尾藓、柔叶(细枝)小鼠尾藓、中华细枝藓、牛角藓原变种、仰叶拟细湿藓、(漆光)镰刀藓、(匙叶)毛尖藓(图版Ⅰ,14、15;图版Ⅱ,8~10、13~15;图版Ⅲ,1~15);(3)具细小颗粒的,如辛氏曲柄藓、葫芦藓、全缘匍灯藓、东亚泽藓、(钝叶)小鼠尾藓、柔叶(细枝)小鼠尾藓、中华细枝藓、牛角藓原变种、仰叶拟细湿藓、 (漆光)镰刀藓(图版Ⅱ,2~7、11~15;图版Ⅲ,1~12);(4)瘤状,如辛氏曲柄藓(图版Ⅱ,2~4);(5)粗疣,如辛氏曲柄藓(图版Ⅱ,2~4);(6)鳞片状,如辛氏曲柄藓、全缘匍灯藓、牛角藓原变种(图版Ⅱ,2~4、11、12;图版Ⅲ,6、7)。大多数种类与叶尖顶端细胞相邻的其他细胞表面角质层纹饰均为不规则纵纹及颗粒,有的具有3、4种纹饰。
3讨论
3.1叶尖微形态特征与生态的关系
由于苔藓植物体型小,结构简单,以往对于高山地区植物功能生态学研究基本上忽略了苔藓植物的作用[16],实际上苔藓植物在植被类型的演替和水土保持中却起着非常大的作用[6,12],因此现在也越来越引起国内外学者的注意,他们通过对荒漠藓类植物的耐旱机制的研究发现,调整藓类植物叶片在植株上的形态以及叶细胞的超微结构,就能够有效地维持其持水力和光能捕获量的平衡,使苔藓植物从失水中恢复生机,在干旱的环境下存活下来,进而成为生态系统稳定和退化生态系统恢复评价的重要指标之一[17-24],因此,对植物叶尖的微形态特征的研究,是可以与环境因素进行定量分析的。本实验所用的12种藓类植物均密集丛生在海拔较高的寒冷、风多并且空气干燥地区,不仅大多数植物的叶片会强烈卷缩并紧紧裹在茎上,毛尖上部及顶端细胞也呈不同程度的弯曲和拧状,这些方式均可以延缓植物体内水分蒸发速度,降低叶片水分的蒸发量,并抵御长期寒冷、干旱和太阳辐射[7],同时,当水分条件好时,植株可以依靠毛细管传导系统,迅速有效地吸收水分并进行光合作用[23],而苔藓植物的叶尖及顶端细胞也是毛细管系统的一部分,因此,这些藓类植物透明的叶尖及厚壁的顶端细胞,在干时细胞壁有不同程度的凹陷、细胞表面有细小的颗粒、具不规则纵纹等,这些都是苔藓植物为适应长期生长在寒冷、多风、干旱的环境中能反射强光照射以减少水分蒸发,降低强光对叶片所造成的危害[7,25-27],同时还可以在第一时间吸收空气中的水分,传给相邻的其他细胞。叶尖其他的细胞表面比顶端细胞表面粗糙,角质层纹饰密集而多样,这样的结构一方面能抵御恶劣气候环境的伤害,另一方面也有利于吸收水分,使植物迅速恢复有效的生理机能。
3.2叶尖微形态特征对于藓类植物的分类学意义
乌鲁木齐河源区生长的12种藓类植物具有一些相似的结构特征,绝大多数种类的叶先端具叶尖,有的叶尖渐尖,如斜蒴对叶藓、辛氏曲柄藓、东亚泽藓、中华细枝藓、牛角藓原变种、(漆光)镰刀藓;有的叶尖毛状,有的叶尖锥形,如平肋提灯藓和全缘匍灯藓,等等,可以看出,在光镜下这些叶尖先端的特征在分类学上的意义不大,但在扫描电镜下,由于照片的放大倍数远远大于光镜的,因此可以较清楚地看到,不同的藓类植物的叶尖特别是顶端细胞及叶尖的其他细胞表面的多项微形态指标,如在其形态、大小、干时细胞壁的凹陷程度以及表面角质层纹饰的形态、分布等等都有明显的差别,具有一定的分类学价值,特别是在属下种间的区分上[14-17]。依据胡人亮将藓类植物叶先端形态划分成12种类型[3],结合扫描电镜方法,将这12种藓类植物的叶尖形态又分成:(1)渐尖组。A.顶端细胞较短,且叶尖先端具几个小尖头:a.锥形渐尖,如斜蒴对叶藓;b.呈三棱形渐尖,如辛氏曲柄藓;c.扁平状渐尖,如东亚泽藓、(漆光)镰刀藓;B.叶尖先端扁平,只有1个小尖头:如中华细枝藓;C.叶尖先端三棱形,只有1个小尖头:如牛角藓原变种。(2)毛状组。A.叶尖先端三棱形,顶端细胞较长,如葫芦藓;B.叶尖先端针刺状,稍拧,如仰叶拟细湿藓。(3)锥形组。如平肋提灯藓、全缘匍灯藓;(4)圆钝形组,如(钝叶)小鼠尾藓;(5)芒状组,如柔叶(细枝)小鼠尾藓。
3.3叶尖顶端细胞微形态特征对于藓类植物进化趋势的关系
有学者指出导管长的较原始,导管短的较进化[28],而叶尖细胞与导管的功能相近似,我们推测,全缘匐灯藓的的细胞最长,则属于较原始的种类,而柔叶(细枝)小鼠尾藓的叶尖顶端细胞最短,则较属于进化的种类。另外,有研究表明,细胞表面的角质层纹饰是由简单向复杂的方向演化的[29],因此,顶端细胞较光滑的斜蒴对叶藓、辛氏曲柄藓属于较原始的,而(漆光)镰刀藓属于较进化的种类。这两种结果并不一致,说明植物进化是一个漫长而复杂的变化过程,需要多方面综合的分析研究,如与叶尖顶端细胞相邻的其他叶尖细胞表面角质层纹饰,不仅种与种之间不同,而且与同种的叶尖顶端细胞也有所不同,角质层纹饰在叶尖的分布及形态特征也是不同的,而辛氏曲柄藓的角质层纹饰在叶尖顶端细胞表面较光滑,但随着细胞越来越向叶片靠近,出现了小孔、鳞片、小瘤,直到出现大量密集的粗疣,角质层纹饰由简单到复杂又到简单,难以确定它属于进化种类还是原始种类,这可能是它为了更好的吸收水分,反射强光、抵御寒冷等适应环境而形成的生存策略,有待今后进行全面的形态、生理、生态等方面的深入研究。
致谢:实验材料由杭州师范大学吴玉环教授鉴定并提供,特此感谢!
参考文献:
[1]安黎哲,刘艳红,冯国宁,等.乌鲁木齐河源区高寒冰缘植被的生态特征研究[J].西北植物学报,2000,20(1):98-105.
AN L Z,LIU Y H,FENG G N,etal.Studies on ecological properties of altifrigetic subnival vegetation at the source area of Urumqi river[J].ActaBot.Boreal.-Occident.Sinica,2000,20(1):98-105.
[2]胡人亮.苔藓植物学[M].北京:高等教育出版社,1987:1-188.
[3]黎兴江,臧穆,曾淑英.西藏苔藓植物志[M].北京:科学出版社,1985:11-370.
[4]白学良.内蒙古苔藓植物志[M].呼和浩特:内蒙古大学出版社,1997,115-437.
[5]熊源新.贵州苔藓植物志[M].贵阳:贵州科技出版社,2014:334-400.
[6]吴玉环,程国栋,高谦.苔藓植物的生态功能及在植被恢复与重建中的作用[J].中国沙漠,2003,23(3):215-220.
WU Y H,CHENG G D,GAO Q.Bryophyte’s ecology functions and its significances in revegetation[J].JournalofDesertResearch,2003,23(3):215-220.
[7]张元明,曹同,潘伯荣.干旱与半干旱地区苔藓植物生态学研究综述[J].生态学报,2002,22(7):1 129-1 134.
ZHANG Y M,CAO T,PAN B R.A review on the studies of bryophyte ecology in arid and sem iarid areas[J].ActaEcologicaSinica,2002,22(7):1 129-1 134.
[8]赵建成.新疆博格达山苔藓植物的研究[J].新疆大学学报(自然科学版),1993,10(1):73-91.
ZHAO J C.Study on the bryophytes on Mt.Bogda,Xinjiang[J].JournalofXinjiangUniversit(Nat.Sci.Edi.),1993,10(1):73-91.
[9]赵建成.新疆东部天山苔藓植物区系[J].Chenia,1993,(1):99-114.
ZHAO J C.Bryophyte flora of East Mount Tianshan in Xinjiang[J].Chenia,1993,(1):99-114.
[10]买买提明·苏莱曼,赵建成.新疆天山苔醉植物新记录及其地理分布[J].干旱区研究,1996,13(1):32-35.
SULAYMAN M,ZHAO J C.Some new records of bryophytes in Tianshan Mts and their geographical distributions[J].AridZoneResearch,1996,13(1):32-35.
[11]张元明,曹同,潘伯荣.新疆博格达山地面生苔藓植物物种多样性研究[J].应用生态学报,2003,14(6):887-897.
ZHANG Y M,CAO T,PAN B R.Species diversity of floor Bryophyte communities in Bogda Mountains,Xinjiang[J].ChineseJournalofAppliedEcology,2003,14(6):887-897.
[12]张元明,曹同,潘伯荣.新疆三工河流域的苔藓植物及其分布特征[J].干旱区研究,2002,19(1):39-43.
ZHANG Y M,CAO T,PAN B R.Study on bryophytes and their distribution patterns in Sangong River Watershed,Xinjiang[J].AridZoneResearch,2002,19(1):39-43.
[13]王虹,姜彦成,苏俊.一号冰川地区4种藓类植物的解剖学研究[J].植物研究,2008,28(1):25-29.
WANG H,JIANG Y C,SU J.Anatomy studies of four mosses in glacier No.1 of Xinjiang[J].BulletinofBotanicalResearch,2008,28(1):25-29.
[14]姜彦成,王虹,王永波,等.紫萼藓科植物茎及叶的解剖学研究(续)[J].植物研究,2007,27(5):534-537.
JIANG Y C,WANG H,WANG Y B,etal.Anatomical structure of stems and leaves of grimmiaceae (continue)[J].BulletinofBotanicalResearch,2007,27(5):534-537.
[15]曾健,吴玉环,王虹,等.新疆天山一号冰川地区十种藓类植物叶尖的微形态观察[J].植物研究,2015,35(5):672-678.
ZENG J,WU Y H,WANG H,etal.Micromorphological structure of leaf apex of ten species of bryophytes from No.1 glacier of Tianshan Mountains[J].BulletinofBotanicalResearch,2015,35(5):672-678.
[16]柯勒著,吴宁,等译.高山植物功能生态学[M].科学出版社,2009:1-306.
[17]吴玉环,陈佳强,冯虎元,等.耐旱藓类的抗旱生理及其机理研究[J].中国沙漠,2004,24(1):23-28.
WU Y H,CHEN J Q,FEN H Y,etal.Advances of research on desiccation-tolerant moss[J].JounalofDesertResearch,2004,24(1):23-28.
[18]张萍,白学良.耐旱苔藓植物生理学及分子生物学研究进展[J].内蒙古大学学报(自然科学版),2004,35(2):227-233.
ZHANG P,BAI X L The development of the physiology and molecular biology of the desiccation-tolerant mosses[J].ActaScientiarumNaturaliumUniversitatisNeimongol(Nat.Sci.Edi.),2004,35(2):227-233.
[19]张萍,白学良,钟秀丽.苔藓植物耐旱机制研究进展[J].植物学通报,2005,22(1):107-114.
ZHANG P,BAI X L.ZHONG X L.Advances in the desiccation tolerance of mosse[J].ChineseBulletinofBotany,2005,22(1):107-114.
[20]WU N,ZHANG Y M,DOWNING A,etal.Rapid adjustment of leaf angle explains how the desert moss,Syntrichiacaninervis,copes with multiple resource limitations during rehydration[J].FunctionalPlantBiology,2014,41:168-177.
[21]OEEHEL W C,LAWRENCE W T.Physiological Ecology of North American Plant Communities[M]//Chabot B F and M ooney H A.Taiga.New York:Chapm an and Hall,1985:6-94.
[22]YE T,ZHANG Y M.Effects of leaf hair points of a desert moss on water retention and dew formation:implications for desiccation tolerance[J].JournalofPlantResearch,2012,125(3):351-360.
[23]吴玉环,黄国宏,高谦.苔藓植物对环境变化的影响及适应性研究进展[J].应用生态学报,2001,12(6):943-946.
WU Y H,HUANG G H,GAO Q.Research advance in response and adaptation of bryophytes to environmental change[J].ChineseJournalofAppliedEcology,2001,12(6):943-946.
[24]吴玉环,高谦,陈国栋,等.苔藓植物对全球变化的响应及其生物指标意义[J].应用生态学报,2002,13(7):895-900.
WU Y H,GAO Q,CHEN G D,etal.Response of bryophytes to global change and its bioindieatortation[J].ChineseJournalofAppliedEcology,2002,13(7):895-900.
[25]吴鹏程.苔藓植物生物学[M].北京:科学出版社,1998:1-338.
[26]郑云普,赵建成,张丙昌,等.荒漠生物结皮中藻类和苔藓植物研究进展[J].植物学报,2009,44(3):371-378.
ZHENG Y P,ZHAO J C,ZHANG B C,etal.Advances on ecological studies of algae and mosses in biological soil crust[J].ChineseBulletinofBotany,2009,44(3):371-378.
[27]ZANDER R H.Genera of the Pottiaceae:mosses of harsh environments[J].Bull.BuffaloSoc.Nat.Sci.,1993,32:1-378.
[28]詹琪芳,王幼芳,李粉霞,等.两种生境下的8种藓类植物茎结构的比较解剖学研究[J].西北植物学报,2006,26(2):217-225.
ZHAN Q F,WANG Y F,LI F X,etal.Comparison of stem structures of eight mosses in two habitats[J].ActaBot.Boreal.-Occident.Sin.,2006,26(2):217-225.
[29]沙伟,郑云梅.24种国产金发藓科植物叶中肋细胞离析后形态学比较观察[J].武汉植物学研究,2008,26(2):124-128.
SHA W,ZHENG Y M.The observation of morphology on costa of 24 species of polytrichaceae after separating in China[J].JournalofWuhanBotanicalResearch,2008,26(2):124-128.
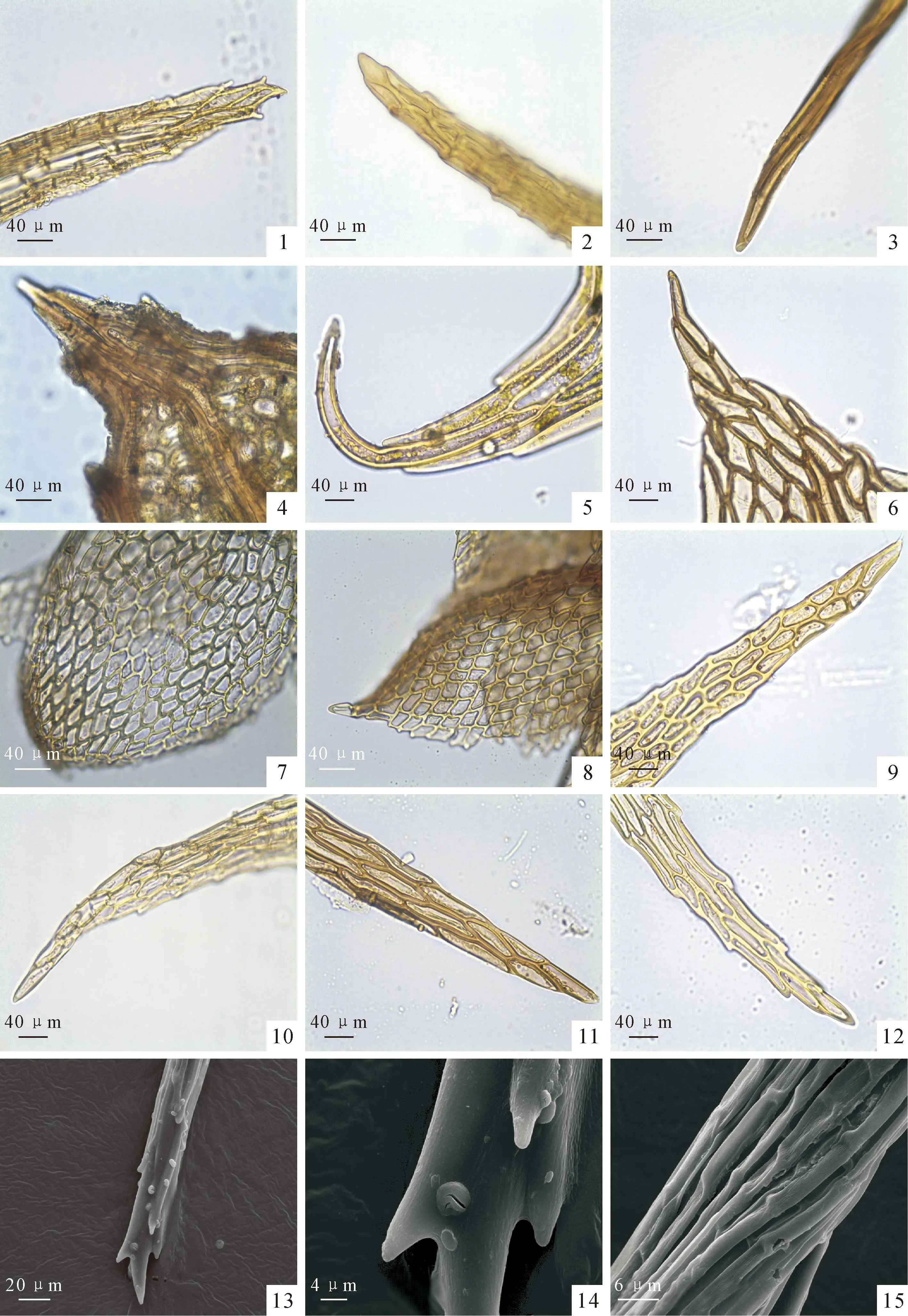
图版 Ⅰ光学显微镜下乌鲁木齐河源区12种藓类植物叶尖形态,×400
1.斜蒴对叶藓;2.辛氏曲柄藓;3.葫芦藓;4.平肋提灯藓;5.全缘匍灯藓;6.东亚泽藓;7.(钝叶)小鼠尾藓;8.柔叶(细枝)小鼠尾藓;9.中华细枝藓;10.(漆光)镰刀藓;11.仰叶拟细湿藓;12.牛角藓原变种;13~15.扫描电镜下斜蒴对叶藓,×8 000,×4 000。
Plate ⅠLight microscope photographs of leaf apex structure of 12 species of bryophytes from source area of Urumqi river,×400
Fig.1.Districhiuminclinatum;Fig.2.Campylopusschimperi;Fig.3.Funariahygrometrica;Fig.4.Mniumlaecinerve;Fig.5.Plagiomniumintegrum;Fig.6.Philonotisturneriana;Fig.7.Myurellajulacea;Fig.8.Myurellatenerrima;Fig.9.Lindbergiasinensis;Fig.10.Drepanocladusvernicosus;Fig.11.Campyliadelphusstellatus;Fig.12.Cratoneuronfilicinum;Fig.13-15.SEM photographsDistrichiuminclinatum,×8 000,×4 000.
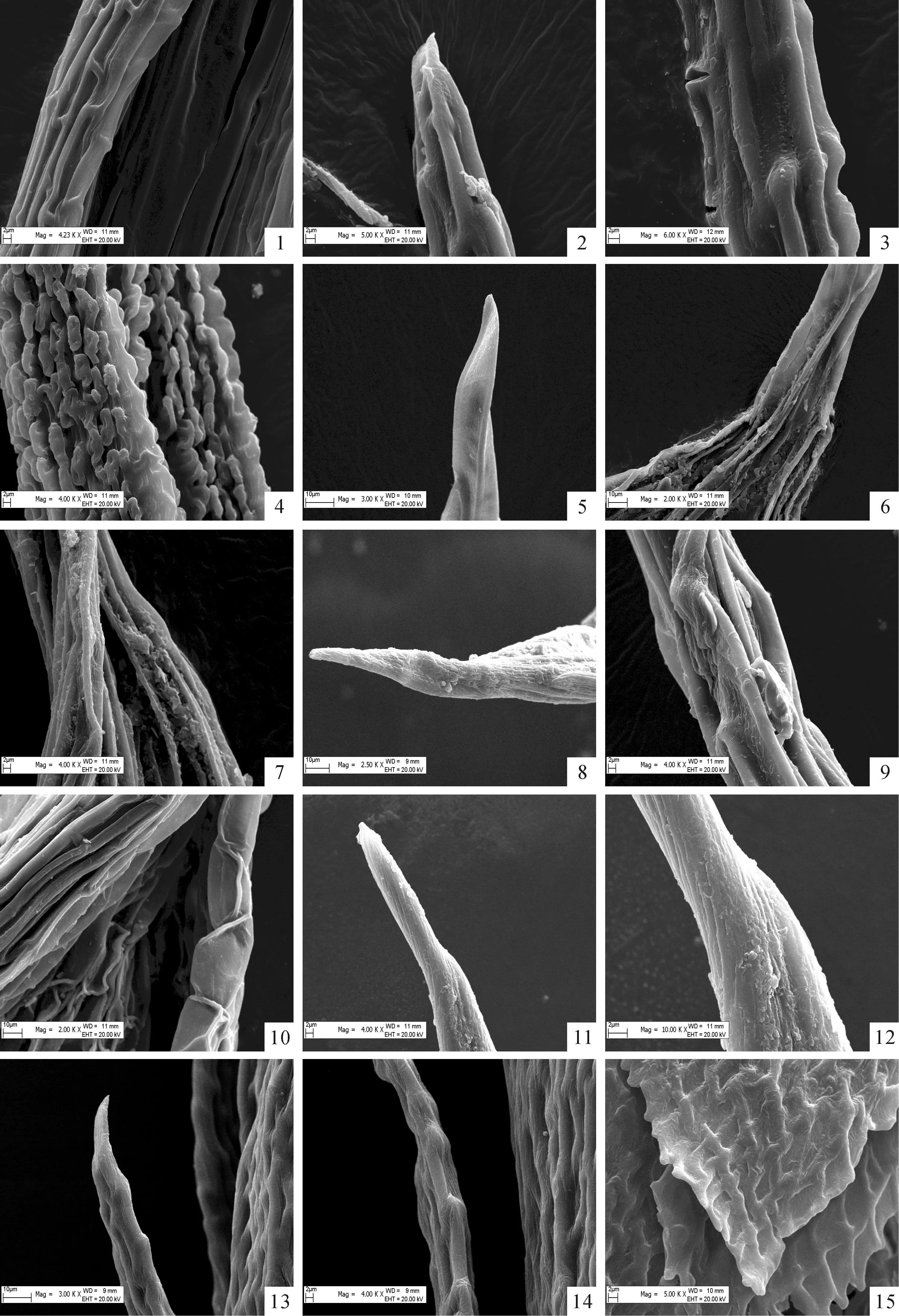
图版 Ⅱ扫描电镜下乌鲁木齐河源区12种藓类植物叶尖形态
1.斜蒴对叶藓,×2 000,×1 500,×3 000;2~4.辛氏曲柄藓,×6 000,×6 000,×4 000;5~7.葫芦藓,×3 000,×4 000,×2 000;8、9.平肋提灯藓,×4 000,×10 000;10~12.全缘匍灯藓,×2 500,×4 000,×2 000;13、14.东亚泽藓,×3 000,×4 000;15.(钝叶)小鼠尾藓,×6 000。
Plate ⅡSEM photographs of leaf apex structure of 12 species of bryophytes from source area of Urumqi river
Fig.1.Districhiuminclinatum,×2 000,×1 500,×3 000;Fig.2-4.Campylopusschimperi,×6 000,×6 000,×4 000;Fig.5-7.Funariahygrometrica,×3 000,×4 000,×2 000;Fig.8,9.Mniumlaecinerve,×4 000,×10 000;Fig.10-12.Plagiomniumintegrum,×2 500,×4 000,×2 000;Fig.13,14.Philonotisturneriana,×3 000,×4 000;Fig.15.Myurellajulacea,×6 000.
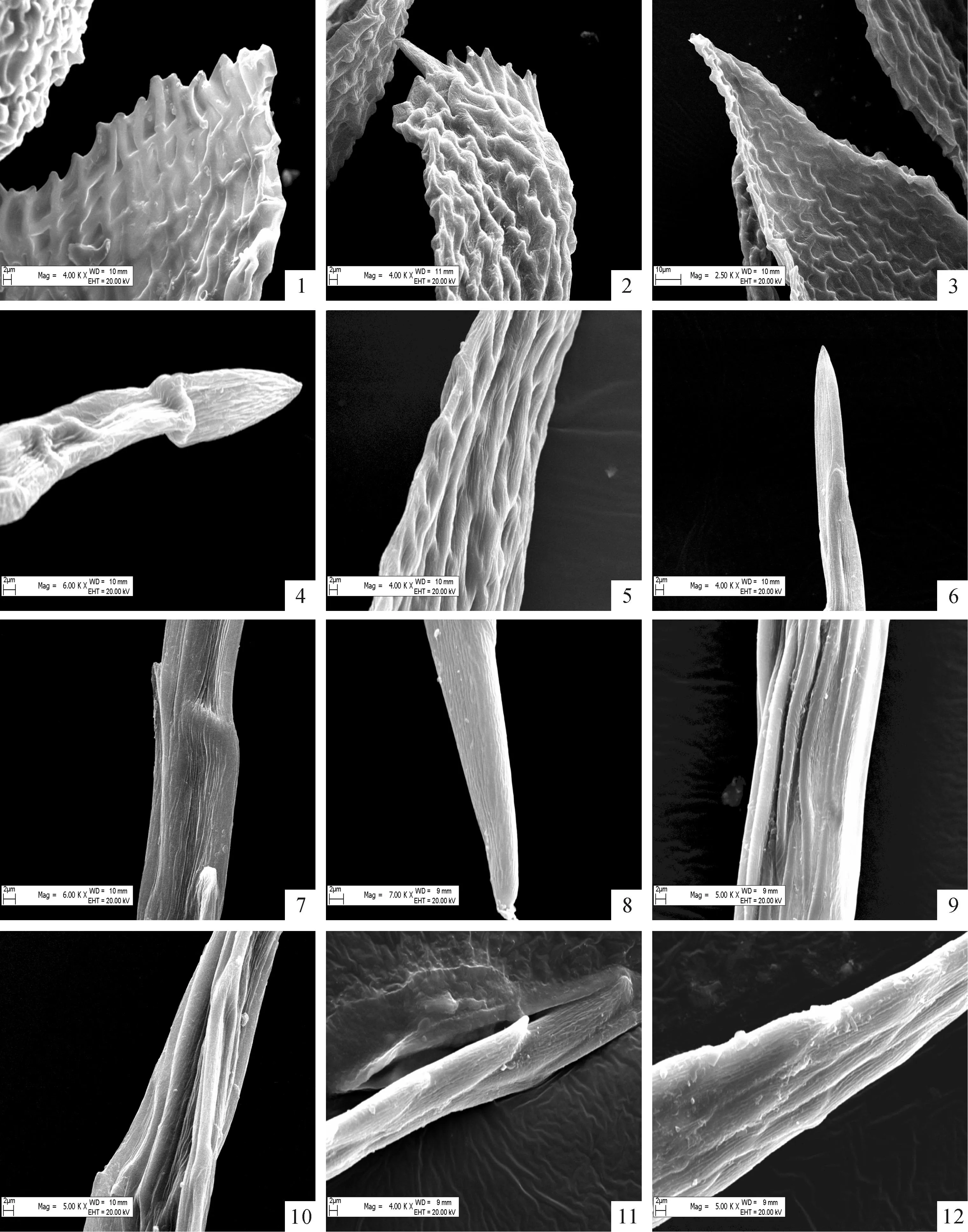
图版 Ⅲ扫描电镜下乌鲁木齐河源区12种藓类植物叶尖形态
1.(钝叶)小鼠尾藓,×8 000;2、3.柔叶(细枝)小鼠尾藓,×3 000,×2 500;4、5.中华细枝藓,×6 000,×4 000;6、7.(漆光)镰刀藓,×4 000,×6 000;8~10.仰叶拟细湿藓,×7 000,×5 000,×5 000;11、12.牛角藓原变种,×4 000;×6 000。
Plate ⅢSEM photographs of leaf apex structure of 12 species of bryophytes from source area of Urumqi river
Fig.1.Myurellajulacea,×8 000;Fig.2,3.Myurellatenerrima,×3 000,×2 500;Fig.4,5.Lindbergiasinensis,×6 000,×4 000;Fig.6,7.Drepanocladusvernicosus,×4 000,×6 000;Fig.8-10.Campyliadelphusstellatus,×7 000,×5 000,×5 000;Fig.11,12.Cratoneuronfilicinum,×4 000,×6 000.
(编辑:潘新社)
Micromorphological Structure of Leaf Apex of Twelve Species of Bryophytes from the Source Area of Urumqi River
WANG Hong,AIFEIRE Abuduaini
(College of Life Science and Technology,Xinjiang University,Urumqi 830046,China)
Abstract:The micromorphological structure of bristle,wart and cutile ornamentation of cells of leaf apex and their neighbor cells of twelve mosses from the source area of Urumqi river were observed by LM and SEM.The results indicated that cells on the top of the leaf apex of most species of the species were transparent,longer and thinner than leaf cell,but with various shape;cell wall was sunken with irregular microgroove and fine particles when it was dry,but not always contracted.Margin of the leaf apex was involuted or twisted,and cell wall was thick with many small pores when it was dry.These characteristics are benefit to absorption of water and adaptive survival strategy for resisting hurt from long period cold and solar radiation.Types of leaf apex have little significance to the classification of family and genus of bryophytes,but the micromorphological structure of cells on the top of the leaf apex and shape,cell number and sunken degree of cell wall and micromorphological structure of wart,mastoid,cutile ornamentation and small pore of other cells are obviously different among different species in same genus,with certain meaning for classification of species under genus level.
Key words:the source area of Urumqi river;bryophytes;SEM;leaf apex;micromorphology
中图分类号:Q246;Q248;Q949.35
文献标志码:A
作者简介:王虹(1962-),女,高级实验师,主要从事植物形态、分类学及细胞发育解剖学的研究。E-mail:wanghong211@163.com
基金项目:国家自然科学基金(41461010,41571049);新疆大学大学生创新训练计划(XJU-SRT-128)
收稿日期:2015-11-23;修改稿收到日期:2016-01-20
文章编号:1000-4025(2016)02-0323-09
doi:10.7606/j.issn.1000-4025.2016.02.0323
猜你喜欢
华声文萃(2022年6期)2022-07-05
上海化工(2021年6期)2021-12-29
航空发动机(2020年3期)2020-07-24
燃气涡轮试验与研究(2019年5期)2019-12-01
中国现代教育装备(2019年21期)2019-11-21
创新作文(小学版)(2019年21期)2019-01-11
智富时代(2018年7期)2018-09-03
智富时代(2018年7期)2018-09-03
中国测试(2017年7期)2017-08-08
岩矿测试(2015年3期)2015-12-21
