
黄柳与小红柳导管分子形态特征及其生态适应性比较研究
2016-03-24 06:29刘冠志刘果厚李红颖刘利红
西北植物学报 2016年2期
关键词:生境
刘冠志,刘果厚*,兰 庆,李红颖,曹 瑞,王 健,刘利红,2
(1 内蒙古农业大学 生态环境学院,呼和浩特 010019;2 正蓝旗草原工作站,内蒙古锡林郭勒盟 027200)
黄柳与小红柳导管分子形态特征及其生态适应性比较研究
刘冠志1,刘果厚1*,兰庆1,李红颖1,曹瑞1,王健1,刘利红1,2
(1 内蒙古农业大学 生态环境学院,呼和浩特 010019;2 正蓝旗草原工作站,内蒙古锡林郭勒盟 027200)
摘要:黄柳和小红柳为浑善达克沙地2种重要的水土保持灌木,黄柳适宜在流动或半流动沙丘上生长,而小红柳适宜在丘间低地生长,二者既可混生,也可形成纯林,存在规律性分布。该研究采用组织离析法和显微照相技术,对黄柳和小红柳的根、茎部导管分子形态特征进行比较研究,探讨导管分子形态特征与其生态适应性的关系,为黄柳和小红柳次生木质部解剖学研究提供资料。结果显示:(1)黄柳和小红柳导管分子均以网纹和两端具尾、一端具尾类型为主,具单穿孔板、互列纹孔。(2)黄柳与小红柳的根部导管分子长度、直径、端壁斜度差异显著,且黄柳较小红柳的直径大、端壁斜度小,为进化特征,而长度较长,为原始特征;但茎部导管分子长度、直径、端壁斜度无显著差异。(3)黄柳与小红柳的导管分子的形态特征与生境间存在一定的相关性,黄柳根部导管具有高输水性而较小红柳适应干旱环境。该研究为黄柳和小红柳造林提供理论指导,并为植物生态适应性的相关研究提供参考。
关键词:黄柳;小红柳;导管分子;生境;生态适应性
黄柳(SalixgordejeviiY.L.Chang et Skv.)与小红柳[S.microstachyavar.bordensis(Nakai) C.F.Fang]为杨柳科(Salicaceae)柳属(Salix)旱中生灌木,高1~2 m,广泛分布于浑善达克沙地,是良好的水土保持植物[1]。然而,经过多年的研究与调查发现,黄柳与小红柳在沙地上的分布随水分、土壤和垂直高度的变化具有一定的规律性。在水肥条件较好的丘间低地,小红柳的分布占绝对优势,黄柳仅有少量生长;在沙丘低处相对高度1~3 m间或丘间低地相对高度在1 m以上处,两者的混生比例相近;在沙丘顶部,相对高度在20 m以上,土壤含水量相对较少,黄柳占绝对优势,小红柳几乎全部消失[2]。在沙丘上垂直分布越高,生长的黄柳越单纯,丛株状态基本消失,而以单株密集呈小片状分布为主;垂直分布越低,纯林面积越小,以丛状混交为主。黄柳和小红柳规律性的分布是其对生态环境适应与改造的反应[3]。二者在造林推广上也有着较大的差异,黄柳较小红柳得到了更广泛的推广[4-5]。由此推测,黄柳较小红柳具有更强的耐干旱、耐沙埋的特性,更适于固定流动或半流动沙地。
植物形态结构与生理功能及生态环境之间相互联系。对于植物的水分生理而言,导管分子无疑是最主要的结构基础。导管分子的形成与进化一直是人们特别关注的问题,其结构特征在研究植物分类以及探索植物细胞与环境间的关系等方面具有非常重要的意义。目前,关于黄柳已进行了诸多方面的研究,主要集中在形态特征[6]、耐热性和耐旱性[7]、光合作用[8]、气孔导度和水分状况[9-10]、种群数量[11]、群落演替[3]、更新复壮[12-13]、根系分布和冠层结构[14]、叶片与根系解剖[15-17]、遗传多样性[18]、造林及再生沙障[2,19]等方面的研究。对小红柳的研究相对较少,主要进行了小红柳的更新复壮[2]、再生沙障[20]、化学成分及体外抗氧化活性[21]等方面的研究。而关于黄柳与小红柳次生木质部导管分子形态特征的研究尚未见报道。本实验通过组织离析方法和显微照相技术,对黄柳与小红柳的根、茎导管分子形态特征进行了比较研究,意在探究二者导管分子形态形态特征与其生态适应性的关系,为黄柳和小红柳次生木质部解剖学研究填补空白,并为导管分子与环境间关系的研究提供参考。
1材料和方法
1.1研究地区自然概况
研究区位于内蒙古锡林郭勒盟正蓝旗桑根达来镇人工造林区,地理位置42°41′~42°43′N,115°57′~115°59′E。该区域属于温带半干旱大陆性季风气候。根据内蒙古正蓝旗气象局提供的气象资料,该地区年平均温度1.5 ℃,极端最低温度-40 ℃,极端最高温度35 ℃,平均日照数为3 037 h,≥10 ℃年积温为2 000 ℃;年均降雨量367.1 mm,多集中在7~9月;年均蒸发量1 925.5 mm;无霜期110 d;土壤类型为栗钙土,其上分布有风沙土。植被类型为乔木、灌木和草本,其中乔木有浑善达克沙地榆(Ulmuspumilavar.sabulosaJ.H.Guo Y.S.Li et J.H.Li),灌木有黄柳、小红柳、羊柴(HedysarumlaeveMaxim.)等;草本植物主要有沙芦草(AgropyronmongolicumKeng)、羊草[Leymuschinensis(Trin.) Tzvel.]、大针茅(StipagrandisP.Smirn.)和星毛委陵菜(PotentillaacaulisL.)等。
1.2材料
实验材料取自3年生的黄柳和小红柳人工林,采样时间为2014年5月,随机选取生长状况良好,冠幅和高度相近的3年生灌丛3个,在直径约1 cm的枝条上截取茎段,并挖取直径0.5 cm以上的根,截取根段,保存带回实验室。
1.3实验方法
把茎段和根段次生木质部切成火柴棒粗细的小段,长约10 mm,煮沸几次至全部下沉为止,取出材料,放置于装有离散液(10%铬酸∶10%硝酸=1∶1)[22]的瓶中,置于36 ℃恒温箱中离散3~7 d(因材料不同,离散时间不同,至材料易被捣散为止)。倒去离散液,清水冲洗,放于70%乙醇长期保存备用。用镊子夹少许材料放于培养皿中,再用玻璃棒将材料轻轻捣散,加1滴1%番红,染色5 min,吸去染液,加少许水将材料均匀冲散,用胶头滴管吸取少许材料放于载玻片,中性树胶封片。
采用Motic BA210光学显微镜观察导管分子的形态,并拍照,利用Motic Images Plus 2.0 ML软件进行导管长度、直径、端壁斜度指标的测量。每个重复分别选取50个导管进行各指标的测量。
1.4数据分析
数据采用Excel 2010、SPSS Statistics 19.0软件进行分析处理。
2结果与分析
2.1导管分子类型
从细胞壁木质化的加厚方式来看,黄柳与小红柳根、茎次生木质部导管分子的类型均以网纹导管(图版Ⅰ,1、3、5、7)为主,具有少量的孔纹导管(图版Ⅰ,2、4、6、8);导管端壁上的穿孔板均为单穿孔板类型(图版Ⅰ,2、3、6、7),纹孔排列方式为互列式(图版Ⅰ,1、3、5、7)。
从具尾情况来看,黄柳与小红柳的导管分子为两端具尾(图版Ⅰ,9、12、15、18),一端具尾(图版Ⅰ,10、13、16、19)和两端无尾(图版Ⅰ,11、14、17、20)3种类型。黄柳与小红柳次生木质部的导管分子以两端具尾和一端具尾为主,两端无尾较少(图1)。其中黄柳茎部两端具尾和一端具尾的导管比例显著大于根部,而两端无尾显著小于根部(P<0.05);小红柳茎部和根部两端具尾比例无显著性差异(P>0.05),而茎部一端具尾比例显著大于根部,两端无尾显著小于根部(P<0.05)。从二者根部导管分子比较来看,黄柳两端具尾的比例显著小于小红柳,两端无尾的比例显著大于小红柳(P<0.05),一端具尾的比例与小红柳无显著性差异(P>0.05);从二者茎部导管分子比较来看,二者两端具尾、一端具尾和两端无尾的比例均无显著性差异(P>0.05)。
2.2导管分子长度
从表1可以看出,黄柳根部与茎部导管分子长度具有显著性差异(P<0.05),前者较后者大124.77 μm;小红柳根部与茎部导管分子无显著性差异(P>0.05)。黄柳与小红柳根部导管分子长度具有显著性差异(P<0.05),分别为379.44和312.41 μm,前者较后者大67.03 μm;二者茎部导管分子长度无显著差异(P>0.05),分别为254.67和236.85 μm。根据国际木材解剖学家协会理事会的标准[23]对导管分子长度的进行等级划分,黄柳根导管分子为中级水平,茎导管分子与小红柳根、茎导管分子均为短级水平。
从黄柳与小红柳根、茎导管分子长度范围分布情况可以看出(图2),黄柳根部导管分子长度主要在300~450 μm,占长度范围的66.66%;小红柳根部导管分子长度主要在250~400 μm,占62.75%;黄柳茎部导管分子长度主要为150~350 μm,占96.96%,小红柳茎部导管分子长度主要在150~300 μm,占80.01%。

不同小写字母表示同一指标不同处理间在0.05水平存在显著差异
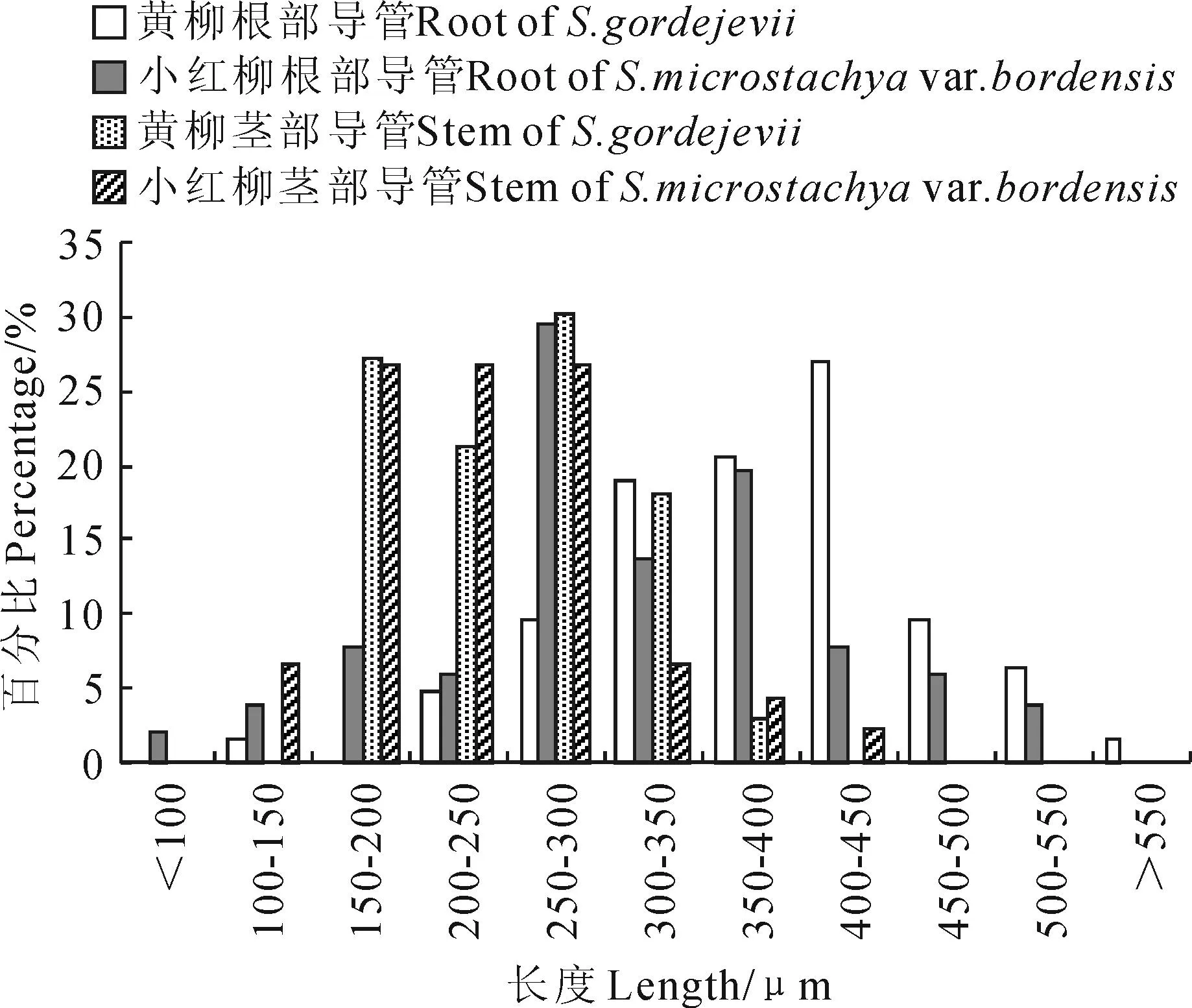
图2 黄柳与小红柳根、茎部导管分子长度范围
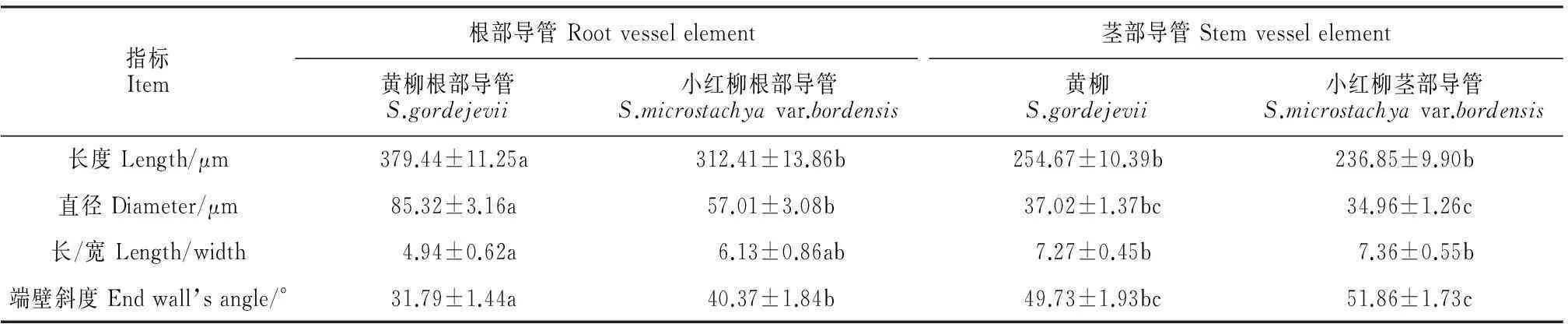
指标Item根部导管Rootvesselelement黄柳根部导管S.gordejevii小红柳根部导管S.microstachyavar.bordensis茎部导管Stemvesselelement黄柳S.gordejevii小红柳茎部导管S.microstachyavar.bordensis长度Length/μm379.44±11.25a312.41±13.86b254.67±10.39b236.85±9.90b直径Diameter/μm85.32±3.16a57.01±3.08b37.02±1.37bc34.96±1.26c长/宽Length/width4.94±0.62a6.13±0.86ab7.27±0.45b7.36±0.55b端壁斜度Endwall’sangle/°31.79±1.44a40.37±1.84b49.73±1.93bc51.86±1.73c
注:同行中不同字母表示差异显著(P<0.05)。
Note:Different letters indicate significant difference in the same row(P<0.05).
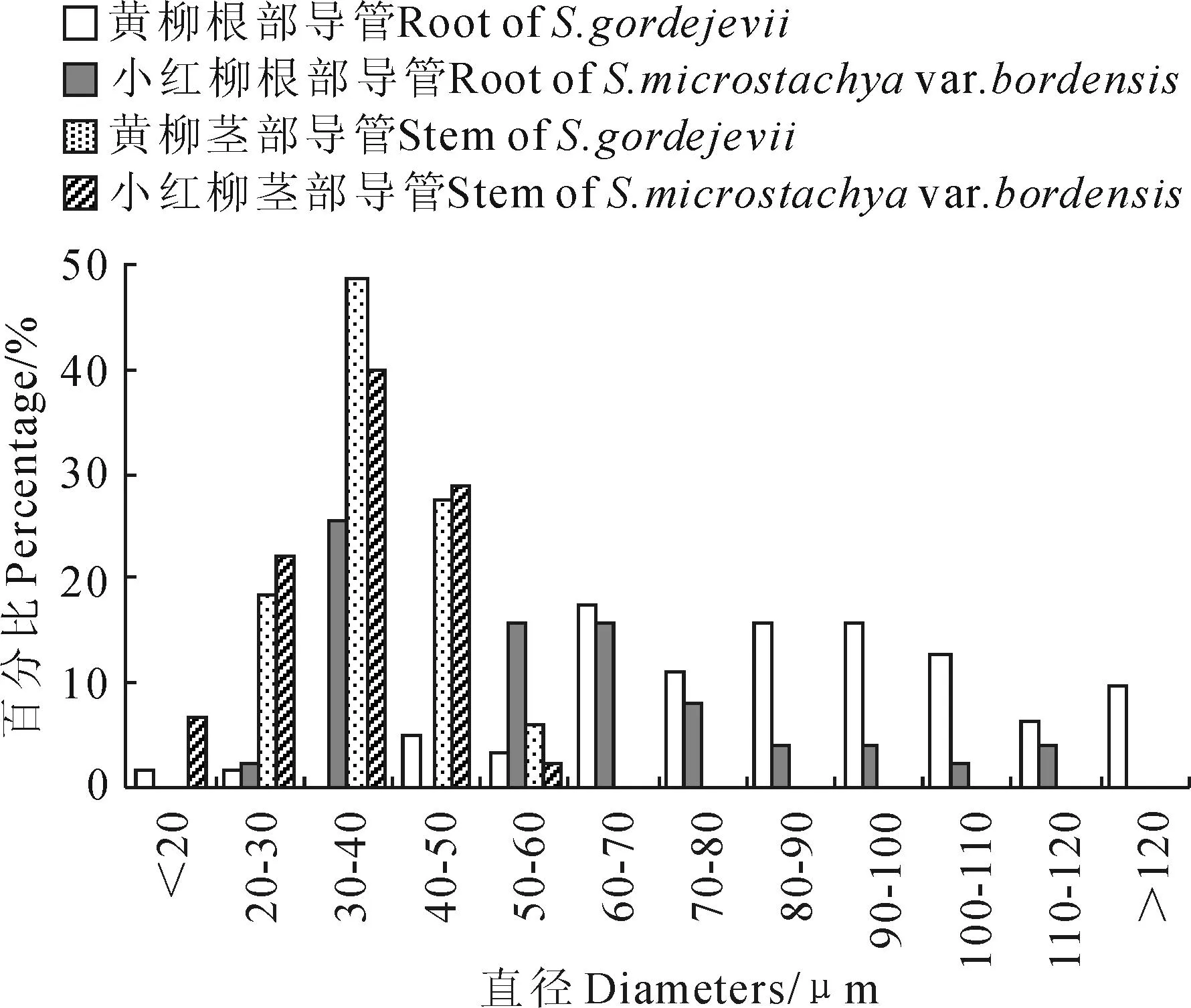
图3 黄柳与小红柳根、茎部导管分子直径范围
2.3导管分子直径
从表1可以看出,二者根部与茎部导管分子直径具有显著性差异(P<0.05),且其根部导管较茎部分别大48.30和22.05 μm。黄柳与小红柳根导管分子直径具有显著差异(P<0.05),分别为85.32和57.01 μm,前者较后者大28.31 μm;二者茎导管分子直径无显著差异(P>0.05),分别为37.02和34.96 μm。根据国际木材解剖学家协会理事会的标准[23]对导管直径进行等级划分,黄柳与小红柳根、茎导管分子直径均为小级水平。
从黄柳与小红柳根、茎导管分子直径范围分布情况可看出(图3),黄柳根部导管分子直径主要分布在60~110 μm,占73.01%。小红柳根部导管分子直径主要在30~70 μm,占76.48%;二者茎部导管分子直径主要分布在20~50 μm,分别占93.93%和91.11%。
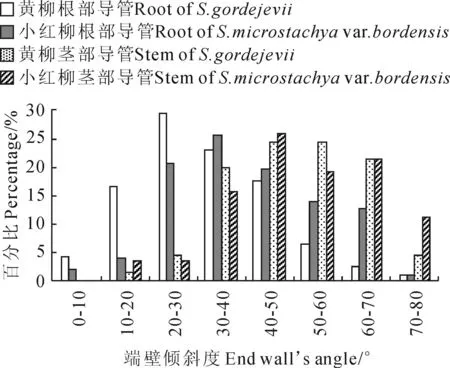
图4 黄柳与小红柳根、茎部导管分子端臂倾斜度范围
2.4导管分子长/宽
黄柳根部与茎部导管的长/宽具有显著差异性(P>0.05),小红柳根部与茎部导管无显著差异性(P>0.05)。黄柳与小红柳根导管分子长/宽无显著差异(P>0.05),分别为4.94和6.13(表1);二者茎导管分子长/宽无显著差异(P>0.05),分别为7.27和7.36。
2.5导管分子端壁斜度
黄柳与小红柳的导管分子端壁斜度比较分析(表1),二者根部与茎部导管分子端壁斜度具有显著性差异(P<0.05),且其根部导管较茎部分别小17.94°和11.49°。二者根部导管分子端壁斜度具有显著差异(P<0.05),分别为31.79°和40.37°,小红柳较黄柳的大8.58°;二者茎部导管分子的端壁斜度无显著差异(P>0.05),分别为49.73°和51.86°。
对黄柳与小红柳根、茎导管分子端壁斜度分布范围进行分析(图4),黄柳根部导管分子端壁斜度主要分布在10°~50°,占86.52%,小红柳根部导管分子端壁斜度主要分布在20°~70°,占92.17%;黄柳茎部导管分子端壁斜度主要分布在30°~70°,占89.39%,小红柳根茎导管分子端壁斜度主要分布在30°~80°,占93.26%。
3讨论
3.1植物导管分子特征与演化
导管分子的类型的进化顺序为:环纹导管分子→螺纹导管分子→梯纹导管分子→网纹导管分子→孔纹导管分子;纹孔排列方式的演化趋势为:对列→互列;穿孔板的类型中梯状穿孔板是最原始的类型,而且横隔越少,进化程度越大,单穿孔板是最进化的类型;导管分子无尾较有尾更为进化[24]。黄柳与小红柳均以网纹导管为主,具有少量孔纹导管;二者纹孔排列方式为互列式;从导管具尾情况来看,二者均以两端具尾和一端具尾导管类型为主,两端无尾导管类型较少。二者根部较茎部两端无尾率大,说明二者根部导管较茎部进化。分析得出黄柳导管无尾率较小红柳的大。因此,黄柳导管分子在类型上较小红柳略为进化。
从演化趋势上来看,短导管分子较长导管分子进化;导管分子直径越大,进化程度越高,长/宽越小,进化程度越高[24]。黄柳根部导管长度较茎部原始,但直径较为进化,长/宽进化程度较一致;小红柳根部与茎部导管长度、长/宽进化程度较为一致,而直径较为进化。虽然黄柳和小红柳的茎部导管分子的长度、直径无显著差异(P>0.05),但根部导管分子的长度、直径具有显著性差异(P<0.05),黄柳的导管分子长度较小红柳长67.03 μm,而直径较小红柳大28.31 μm。所以,从导管分子长度的演化水平来看,小红柳较为进化,黄柳较为原始;从导管分子的直径演化水平来看,黄柳较为进化,小红柳较为原始。因此,也可以看出,导管分子的某些特征在进化中存在异速性[25]。从长/宽来看,二者并无显著性差异(P>0.05)。
从演化趋势上来看,导管分子端壁斜度越大越原始[24]。黄柳和小红柳根部导管较茎部导管端壁斜度进化;二者的茎部导管分子端壁斜度无显著差异(P>0.05),而根部导管分子端壁斜度有显著性差异(P<0.05),前者根部导管分子端壁斜度较后者小8.58°。所以,从导管分子端壁斜度演化水平上来看,黄柳较为进化,小红柳较为原始。
综合以上分析,黄柳根部导管分子无论从类型还是特征方面均较茎部进化,而小红柳根部仅部分特征较茎部表现出一定的进化趋势。二者比较来看,黄柳根部导管分子明显较小红柳进化,而二者茎部进化程度相似。
3.2植物导管分子特征与生境的关系
从物理学角度分析,导管分子长度、直径和端壁斜度的大小决定了水分在植物体内运输阻力的大小和运输效率的高低。导管分子长度越长(单位长度内横壁越少),直径越大,端壁斜度越小(端壁面积较小),则输水阻力越小,输水效率越高,反之,则输水阻力越大,输水效率越小[26-27]。黄柳和小红柳根部导管分子相对于茎部的长度长、直径大、端壁斜度小,因此说明二者存在共性,即根部导管分子发挥着较强的输水性,而茎部导管分子发挥着较强的水分运输安全性。黄柳根部导管分子的长度相对于小红柳长,直径相对较大,端壁斜度相对较小,说明二者也存在不同,即黄柳的根部导管特征更利于水分运输。因此,从水分运输状况来看,黄柳较小红柳具有更强的耐干旱、耐沙埋的特性,更适应在干旱环境生长,而小红柳适应在水分条件相对较好的环境生长。因此,形成了二者在浑善达克沙地上的规律性分布。然而,李国秀等[26]认为适应干旱环境的物种导管长度较长、直径较小。分析认为长度较长、直径较小的导管虽能增加水分的内聚力,从而保证水分运输的安全性和便于存贮性,但不利于水分的高效运输,而长度较长、直径较大的导管能够为其提供保障。说明导管分子进化过程与植物自身的生物学特性有关,其进化异速性特征间可能存在功能上的平衡。
以上结论与黄柳和小红柳在浑善达克沙地规律性分布的特点相一致,表明了导管分子的形态特征与生境间具有一定的相关性,与Carlquist[28]提出的木质部进化的生态学途径相一致。同时依据植物与环境协同进化的理论,说明干旱环境加速了导管分子的进化[29]。本研究结论也为近年来黄柳能够在防风固沙及大面积造林中被广泛应用的实践提供理论支持。
参考文献:
[1]马毓泉.内蒙古植物志(第2卷)[M].呼和浩特:内蒙古人民出版社,1990:69-73.
[2]李佃勇.营造小黄柳林和更新小红柳林是治理浑善达克沙地的有效途径[J].中国沙漠,1983,3(3):40-44.
LI T Y.An effective approach of afforestingSalixgordejeviiand rejuvenatingSalixmicrostachyavar.bordensisto Hunshandake sandland[J].JournalofDesertResearch,1983,3(3):40-44.
[3]林品一,陈秀英.浑善达克沙地小黄柳的生长状况及与小红柳的演替规律[J].中南林业调查规划,1996,15(2):22-25,28.
LIN Y P,CHEN X Y.The growth status ofSalixgordejeviiand the evolutionary rules ofSalixmicrostachyavar.bordensisin Hunshandake Sandland[J].CentralSouthForestInventoryandPlanning,1996,15(2):22-25,28.
[4]王晟,李君.黄柳生物再生沙障在巴盟的推广应用[J].内蒙古林业,2003,(12):28.
WANG SH,LI J.The popularization and application ofSalixgordejeviiliving sand barrier in Bayenaoer league[J].JournalofInnerMongoliaForestry,2003,(12):28.
[5]潘湘海.黄柳扦插造林试验研究[J].河北林业科技,1999,(2):4-5.
PAN X H.The afforestation ofSalixgordejeviicutting experimental study[J].JournalofHebeiForestryScienceandTechnology,1999,(2):4-5.
[6]乔本梅.黄柳[J].中国水土保持,1995,(12):25.
QIAO B M.Salixgordejevii[J].SoilandWaterConservationSocietyinChina,1995,(12):25.
[7]杨甲定,赵哈林,张铜会.黄柳与垂柳的耐热性和耐旱性比较研究[J].植物生态学报,2005,29(1):42-47.
YANG J D,ZHAO H L,ZHANG T H.A Comparative study on heat and drought tolerance betweenSalixgordejeviiandSalixbabylonica[J].ActaPhytoecologicalSinica,2005,29(1):42-47.
[8]高玉葆,任安芝.科尔沁沙地黄柳再生枝与现存枝形态和光合特征的比较[J].生态学报,2002,22(10):1 758-1 764.
GAO Y B,REN A ZH.A comparative study on photosynthesis and morphological characteristics ofSalixgordejeviibetween regenerated shoots and standing shoots[J].ActaEcologicalSinica,2002,22(10):1 758-1 764.
[9]周海燕,赵爱芬.科尔沁沙地黄柳气孔导度和水分状况随季节和地形的变化特征[J].植物学通报,2000,17(6):543-547.
ZHOU H Y,ZHAO A F.Seasonal and topographical changes in stomatal conductance and water state ofSalixgordejeviiin Horqin sandy land[J].ChineseBulletinofBotany,2000,17(6):543-547.
[10]崔秀萍,刘果厚.黄柳叶片对水分亏缺的生理响应[J].干旱区资源与环境2009,23(3):181-187.
CUI X P,LIU G H.Physiological response ofSalixgordejeviileaves under the conditions of water deficiency[J].JournalofAridLandResourcesandEnvironment,2009,23(3):181-187.
[11]梁宇,高玉葆,任安芝.不同沙地生境下黄柳种群若干数量特征的比较研究[J].生态学报,2000,20(1):80-87.
LIANG Y,GAO B Y,REN A ZH.Quantitative characteristics ofSalixgordejeviipopulation in different sandy land habitats[J].ActaEcologicalSinica,2000,20(1):80-87.
[12]崔秀萍,刘果厚.浑善达克沙地黄柳更新特点与规律研究[J].中国沙漠,2012,32(1):60-64.
CUI X P,LIU G H.Characteristics and rules of regeneration ofSalixgordejeviiin Hunshandake sandland[J].JournalofDesertResearch,2012,32(1):60-64.
[13]胡小龙,薛博,袁立敏,等.科尔沁沙地人工黄柳林平茬复壮技术研究[J].干旱区资源与环境,2012,26(5):135-139.
HU X L,XUE B,YUAN L M,etal.Techniques of stumping and rejuvenating ofSalixgordejeviiplantation in Kerqin sandy land[J].JournalofAridLandResourcesandEnvironment,2012,26(5):135-139.
[14]任安芝,高玉葆,王金龙.不同沙地生境下黄柳的根系分布和冠层结构特征[J].生态学报,2001,21(3):399-404.
REN A ZH,GAO Y B,WANG J L.Root distribution and canopy structure ofSalixgordejeviiin different sandy land habitats[J].ActaEcologicalSinica,2001,21(3):399-404.
[15]崔秀萍,刘果厚,张瑞麟.浑善达克沙地不同生境下黄柳叶片解剖结构的比较[J].生态学报,2006,26(6):1 842-1 847.
CUI X P,LIU G H,ZHANG R L.Comparison of leaf anatomical structure betweenSalixgordejeviigrowing under contrasting habitats of Otingdag sandland andSalixmicrotachyavar.bordensisgrowing on the lowlands of dunes[J].ActaEcologicalSinica,2006,26(6):1 842-1 847.
[16]SU H,LI Y G,LAN ZH J,etal.Leaf-level plasticity ofSalixgordejeviiin fixed dunes compared with lowlands in Hunshandake Sandland,North China[J].JournalofPlantResearch,2009,122:611-622.
[17]刘冠志,刘果厚,贺晓,等.黄柳不同级序根的解剖结构及其细根的研究[J].西北植物学报,2014,34(5):932-937.
LIU G Z,LIU G H,HE X,etal.Anatomical characteristics of roots with different orders and fine root ofSalixgoridevii[J].ActaBot.Boreal.-Occident.Sin.,2015,43(5):123-129.
[18]董建芳.内蒙古黄柳遗传多样性的RAPD分析[D].呼和浩特:内蒙古农业大学,2008.
[19]刘冠志,刘果厚,刘哲荣,等.浑善达克沙地黄柳再生沙障插穗指标[J].东北林业大学学报,2015,43(5):123-129.
LIU G ZH,LIU G H,LIU ZH R,etal.Cuttings individual indicators ofSalixgordejeviiliving sand barrier in Hunshandake sandy land[J].JournalofNortheastForestryUniversity,2015,43(5):123-129.
[20]张瑞麟,刘果厚,崔秀萍.浑善达克沙地黄柳活沙障防风固沙效益的研究[J].中国沙漠,2006,26(5):717-721.
ZHANG R L,LIU G H,CUI X P.Effects of livingSalixgordejeviibarrier on wind-breaking and sand-fixation in Hunshandake sandland[J].JournalofDesertResearch,2006,26(5):717-721.
[21]王婷,赵永亮,赵晓宁,等.小红柳化学成分及体外抗氧化活性研究[J].中药材,2014,37(12):2 222-2 225.
WANG T,ZHAO Y L,ZHAO X N,etal.Constituents and antioxidant activityinvitroofSalixmicrostachyavar.bordensis[J].JournalofChineseMedicinalMaterials,2014,37(12):2 222-2 225.
[22]王英英,刘宁.植物生物学实验[M].北京:高等教育出版社,2005:143.
[23]成俊卿.木材学[M].北京:中国林业出版社,1985:102-104.
[24]李红芳,田先华,任毅.维管植物导管及其穿孔板的研究进展[J].西北植物学报,2005,25(2):419-4 24.
LI H F,TIAN X H,REN Y.Research progress in vessel and perforation plate of vascular plants and someconsiderations for future research[J].ActaBot.Boreal.-Occident.Sin.,2005,25(2):419-424.
[25]陈树思.洋蒲桃次生木质部中导管分子的解剖学[J].植物学通报,2006,23(6):677-683.
CHEN SH S.Anatomy of vessel elements in the secondary xylem ofSyzygiumsamarangense[J].ChineseBulletinofBotany,2006,23(6):677-683.
[26]李国秀,郑宝江.10种茶藨子属植物导管分子形态特征及其生态适应性比较研究[J].植物研究,2014,34(1):25-31.
LI G X,ZHENG B J.Comparative study on morphological characteristics and ecological adaptability of vessel elements of tenRibesL.varieties[J].BulletinofBotanicalResearch,2014,34(1):25-31.
[27]谷利伟.黑龙江绣线菊属15种植物导管分子形态结构研究[J].电子显微学报,2015,34(1):74-75.
GU L W.Morphology structure of vessel element ofSpiraeain Heilongjiang Province[J].JournalofChineseElectronMicroscopySociety,2015,34(1):74-75.
[28]CARLQUIST S.Wood anatomy of coriariaecae:phylogenetic and ecological implications[J].Syst.Bot.,1985,10(2):174-183.

[29]CARLQUIST S,SCHNEIDER E L.Origin and natures of vessels in monocotyledons.5.Araceae subfamily Colocasioideae[J].BotanicalJournaloftheLinneanSociety,1998,128:71-86.图版 Ⅰ黄柳与小红柳根、茎部导管分子形态特征
黄柳:1.根网纹导管、互列纹孔;2.根孔纹导管、单穿孔板;3.茎网纹导管、单穿孔板、互列纹孔;4.茎孔纹导管;9.根两端具尾导管;10.根一端具尾导管;11.根两端无尾导管;12.茎两端具尾导管;13.茎一端具尾导管;14.茎两端无尾导管;小红柳:5.根网纹导管、互列纹孔;6.根孔纹导管、单穿孔板;7.茎网纹导管、单穿孔板、互列纹孔;8.茎孔纹导管;15.根两端具尾导管分子;16.根一端具尾导管;17.根两端无尾导管;18.茎两端具尾导管;19.茎一端具尾导管;20.茎两端无尾导管。
Plate ⅠThe characteristics of root and stem vessel elements ofS.gordejeviiandS.microstachyavar.bordensis
S.gordejevii:Fig.1.Root reticulate vessel element with alternate pitting;Fig.2.Root pitted vessel element with simple perforation plate;Fig.3.Stem reticulate vessel element with simple perforation plate and alternate pitting;Fig.4.Stem pitted vessel element;Fig.9.Both end walls with one tail of root vessel element;Fig.10.A tail in one wall of root vessel element;Fig.11.No tails in both walls of root vessel element;Fig.12.Both end walls with one tail of stem vessel element;Fig.13.A tail in one wall of stem vessel element;Fig.14.No tails in both walls of stem vessel element;S.microstachyavar.bordensis:Fig.5.Root reticulate vessel element of alternate pitting;Fig.6.Root pitted vessel element with simple perforation plate;Fig.7.Stem reticulate vessel element with simple perforation plate and alternate pitting;Fig.8.Stem pitted vessel element;Fig.15.Both end walls with one tail of root vessel element;Fig.16.A tail in one wall of root vessel element;Fig.17.No tails in both walls of root vessel element;Fig.18.Both end walls with one tail of stem vessel element;Fig.19.A tail in one wall of stem vessel element;Fig.20.No tails in both walls of stem vessel element.
(编辑:潘新社)
Comparative Study on Morphological Characteristics and Ecological Adaptability of Vessel Elements ofSalixgordejeviiandS.microstachyavar.bordensis
LIU Guanzhi1,LIU Guohou1*,LAN Qing1,LI Hongying1,CAO Rui1,WANG Jian1,LIU Lihong1,2
(1 College of the Ecological Environment,Inner Mongolia Agricultural University,Hohhot 010019,China;2 The Prairie Workstation in Zhenglanqi,Xilin Gol League,Inner Mongolia 027200,China)
Abstract:Salix gordejevii and S.microstachya var.bordensis are important shrubs for soil and water conservation in Hunshandake sandland.The former is suitable for growing on flow and semi-flow sand dunes,and the latter does on dune slack.They can grow mixed,and also can form pure forest.So that they have the feature of regular distribution.Through the method of tissues segregation and micrographic technique,this paper comparatively studied on the morphological characteristics and ecological adaptability of vessel elements of S.gordejevii and S.microstachya var.bordensis.The results showed that:(1)the two species were given priority to reticulate vessels with both end walls with one tail and a tail in one wall.Both ends of their vessel elements with alternate pitting had simple perforation plates.(2)There were significant differences between the length,diameter and end wall slope angle of xylem vessels in the root of S.gordejevii and S.microstachya var.bordensis(P<0.05).The diameter of root vessel elements of the former was wider and the end wall slope angle of those was smaller than the latter’s,which showed the evolutionary features.However,the length of root vessel elements of the former was longer which showed the primitive feature.There were no significant differences between the length,diameter and end wall slope angle of xylem vessels in the stem of S.gordejevii and S.microstachya var.bordensis(P>0.05).(3)There appeared some correlation between the characteristics of vessel elements and habitats of the two species.The arid environment was more suitable for S.gordejevii to grow due to the higher water transmitting ability than S.microstachya var.bordensis.This study provided theoretical support for afforestation with S.gordejevii and S.microstachya var.bordensis,and provided reference for the correlational research of plant ecological adaptability.
Key words:Salix gordejevii;Salix microstachya var.bordensis;vessel element;ecological adaptability
中图分类号:Q944.5
文献标志码:A
作者简介:刘冠志(1990-),男,在读博士研究生,主要从事植物多样性保护与利用研究。E-mail:liuguanzhi1989@163.com*通信作者:刘果厚,教授,博士生导师,主要从事植物多样性保护和荒漠化植被恢复与重建研究。E-mail:guohouliu@imau.edu.cn
基金项目:内蒙古自治区科技创新引导奖励资金(20121607);内蒙古自治区研究生创新项目(S20131012907)
收稿日期:2015-11-13;修改稿收到日期:2016-01-04
文章编号:1000-4025(2016)02-0316-07
doi:10.7606/j.issn.1000-4025.2016.02.0316
猜你喜欢
——基于荟萃分析和遥感大数据分析
生态学报(2022年12期)2022-08-03
生物学杂志(2022年1期)2022-02-21
中国农村水利水电(2021年11期)2021-11-29
食品与发酵工业(2021年17期)2021-09-17
风景园林(2021年2期)2021-04-24
落叶果树(2021年6期)2021-02-12
福建质量管理(2020年11期)2020-06-18
现代园艺(2018年1期)2018-03-15
华东师范大学学报(自然科学版)(2017年1期)2017-02-27
体育科技(2016年2期)2016-02-28
