
烤烟‘南江3号’大小孢子的发育及雌雄配子体的形成
2016-03-24 06:32高凯悦吉志超廖海民
西北植物学报 2016年2期
关键词:烤烟
高凯悦,吉志超,廖海民
(贵州大学 生命科学学院,贵阳 550025)
烤烟‘南江3号’大小孢子的发育及雌雄配子体的形成
高凯悦,吉志超,廖海民*
(贵州大学 生命科学学院,贵阳 550025)
摘要:以贵州烤烟主栽品种‘南江3号’为试验材料,采用石蜡切片法对其大小孢子发生及雌雄配子体发育进行观察研究,探讨烤烟生殖过程与花蕾形态变化的关系,为田间栽培控制花期以及实现烟草的遗传调控提供理论依据。结果表明:‘南江3号’雄蕊的花药壁发育为基本型,绒毡层为腺质异型;小孢子母细胞减数分裂为同时型,四分体排列方式有四面体型和十字交叉型2种;成熟花粉为二细胞型,花粉具3或4个萌发孔。胚珠倒生,薄珠心;孢原细胞体积较大,直接发育为大孢子母细胞,4个大孢子呈线性排列;胚囊发育为蓼型,成熟胚囊为7细胞8核。烤烟‘南江3号’的雌蕊发育晚于雄蕊,当雄蕊发育为四分体时期时雌蕊才开始发育,且花蕾发育到5.0~6.8 cm时胚囊才逐渐成熟。研究认为,烤烟‘南江3号’的大小孢子及雌雄配子体发育各阶段与花蕾形态有关,通过花蕾的形态可初步判断各个发育阶段。
关键词:烤烟;大孢子发生;小孢子发生;雌雄配子体
烤烟是贵州省重要的经济作物,年种植面积19万hm2以上,是国民经济可持续发展的战略资源。‘南江3号’作为贵州选育出的特有烤烟品种,它适应性强,产量高,易烘烤,抗病害能力强,具有良好的经济价值,现已在贵州烟区大面积种植[1]。目前,‘南江3号’的研究主要集中在生态环境适应性、栽培与烘烤技术、烟碱含量及其他化学成分分析等方面[2-6],对其胚胎学方面的研究还未见报道。研究‘南江3号’的胚胎学发育规律可以防止烤烟过早进入生殖生长,以及避免生殖生长时期花蕾和花数量过多而抑制其营养生长。本研究运用石蜡切片技术对‘南江3号’有性生殖过程中大小孢子的发生和雌雄配子体的发育规律进行系统研究,并探讨了生殖过程与花蕾形态变化的相关性,旨在为制定‘南江3号’有效的田间栽培措施、控制花期以及实现烟草的遗传调控提供理论依据。
1材料和方法
1.1材料
供试材料烤烟‘南江3号’来源于贵州省贵阳市开阳县龙岗烟叶站。
1.2方法
实验采用石蜡切片法[7]。2014年5~9月,取大小不同的花蕾材料,测量花蕾的长度,然后浸入70% FAA中固定,乙醇系列脱水,二甲苯透明,石蜡包埋。切片厚度7~9 μm,番红-固绿染色,加拿大树胶封片。显微镜观察并拍照。
2结果与分析
2.1小孢子体的发生和雄配子体的发育
烤烟‘南江3号’为两性花,每朵花5个雄蕊,每个花药具4个花粉囊。花药横切呈蝴蝶状,初始期花粉囊较小呈半月型,到小孢子母细胞减数分裂后期花粉囊扩大呈圆形。花药壁的发育方式属基本型。小孢子发生和雄配子体发育过程没有观察到异常现象。
雄蕊原基形成花药原始体后,在4个角隅处细胞分裂较快,呈现出四棱的结构形状,其表皮下出现一个或几个细胞核较大、胞质较浓的孢原细胞。这一时期‘南江3号’还未现蕾,顶花花蕾呈深绿色,长0.30~0.50 cm(表1)。孢原细胞经一次平周分裂,形成内外2层细胞,即外层的初生壁细胞以及内侧的初生造孢细胞(图版Ⅰ,1)。初生壁细胞经数次平周和垂周分裂,产生3~5层细胞,连同表皮构成花药壁,从外向内依次为表皮、药室内壁、2~3层中层和绒毡层(图版Ⅰ,3)。
2.1.1花药壁的发育花药壁发育过程中,表皮形成时间最早,刚形成时,细胞排列紧密,行使一般的保护功能。后随着花药长大逐渐扩展成扁长形,散粉时,仅剩下干枯的残迹。药室内壁在造孢细胞时期分化完全,细胞形状规则,排列整齐(图版Ⅰ,1);在减数分裂时期开始径向延长(图版Ⅰ,4~7);单核花粉时期细胞体积达到最大,液泡化。当花药开裂散粉前,该层细胞的切向壁和径向壁出现明显的纤维状加厚带,细胞质与细胞核已消失(图版Ⅰ,12)。
中层在小孢子母细胞减数分裂过程中,细胞内贮存物质减少,细胞变为扁平(图版Ⅰ,4);在二核花粉后期,中层细胞逐渐解体,最终被吸收并消失(图版Ⅰ,10、11)。
绒毡层细胞初为单核、细胞质浓、液泡少而小,随着小孢子母细胞减数分裂,出现二核甚至多核现象(图版Ⅰ,5)。当减数分裂接近完成时,绒毡层细胞出现退化迹象(图版Ⅰ,6)。到小孢子发育后期和出现雄配子阶段,绒毡层细胞仅留残迹或消失。在花粉发育过程中,绒毡层细胞不断分泌各种物质进入花粉囊,提供小孢子发育直至花粉成熟,大部分原位退化,其绒毡层属腺质型绒毡层类型(图版Ⅰ,12)。
2.1.2小孢子的形成初生造孢细胞有丝分裂形成次生造孢细胞,即小孢子母细胞(图版Ⅰ,2)。小孢子母细胞形态和结构与周围细胞明显不同,排列紧密,具有核大、质浓、无明显液泡等特点,质膜外面沉积大量的胼胝质,细胞核较大,染色体为丝状(图版Ⅰ,3)。这一时期,顶花蕾仍为深绿色,蕾长0.50~0.70 cm。第一次减数分裂过程中,中期清晰可见,染色体排列在细胞赤道板两侧。第一次减数分裂后期也较明显,染色体分开,位于细胞的两极(图版Ⅰ,4)。小孢子母细胞第一次减数分裂完成后,细胞质不发生缢裂。第二次减数分裂完成后,形成为共同的胼胝质包围的4个核(图版Ⅰ,5)。接着细胞质发生缢裂,形成四分体(图版Ⅰ,6)。‘南江3号’小孢子的减数分裂为同时型。四分体排列方式为四面体型和十字交叉型2种(图版Ⅰ,7)。开始减数分裂时顶花花蕾深绿色,长0.70~0.95 cm,形成四分体深绿色花蕾,长已达0.95~1.10 cm(表1)。
2.1.3花粉粒的发育从四分体释放出来的小孢子体积较小,含浓厚的原生质,核位于细胞的中央,它们从绒毡层取得营养,细胞体积不断增大,细胞核由中央位置转向细胞一侧,细胞外壁加厚并形成萌发孔,一般的花粉近球形具3个萌发孔,也有一些花粉呈四方形,具有4个萌发孔(图版Ⅰ,8、9)。 此时花蕾已现,顶花浅黄色花冠伸出,花萼变浅绿色,蕾长1.10~1.40 cm(表1)。随后进行一次不对称的有丝分裂,形成2个细胞的花粉粒(图版Ⅰ,10)。花粉粒中大细胞为营养细胞,内含大量淀粉、脂肪等物质;小细胞为生殖细胞,刚形成的生殖细胞呈纺锤形,以后生殖细胞离开花粉壁进入营养细胞的细胞质中(图版Ⅰ,11)。二细胞花粉期顶花花冠浅黄色,花蕾长1.40~1.70 cm,子房长0.35~0.58 cm(表1)。营养细胞以后与花粉管的生成及生长有关,而生殖细胞的作用是产生2个精细胞,直接参与生殖(图版Ⅰ,12)。此时顶花花蕾长大于1.70 cm,花粉粒已成熟,可见花粉管。
2.2大孢子体的发生和雌配子体的发育
2.2.1胚珠的发育与大孢子的发生‘南江3号’子房由2个心皮合生成2室,中轴胎座,倒生胚珠,胚珠多数,胚囊发育为蓼型。大孢子发生和雌配子体发育过程没有观察到异常现象。
胚珠是在胎座上发生的,初出现时是由薄壁组织的细胞组成的珠心。以后在珠心表皮细胞下分化出一个孢原细胞(图版Ⅱ,1),其细胞体积大,细胞质浓厚,细胞核显著。孢原细胞不经分裂直接分化成大孢子母细胞。此时,珠心细胞也随之进行垂周分裂,不断扩大体积包围大孢子母细胞(图版Ⅱ,2)。这一时期,顶花花冠浅黄色,花蕾长1.40~1.70 cm,子房长0.35~0.58 cm(表1)。随后内外两层珠被逐渐伸长包裹珠心,外珠被形成珠孔,胚珠倒生、多数。大孢子母细胞在减数分裂开始时,染色质凝集,细胞扩大,整个细胞被胼胝质包围。经第一次减数分裂形成二分体(图版Ⅱ,3),而后再进一步分裂形成线型排列的四分体(图版Ⅱ,4、5)。这个过程花蕾长1.70~4.00 cm,顶花花冠浅黄色,子房长0.58~0.70 cm(表1)。合点端的大孢子体积增大,继续发育形成功能大孢子,其余的逐渐退化(图版Ⅱ,6)。
2.2.2雌配子体的发育功能大孢子继续发育,沿珠心纵轴方向生长,液泡化且单核悬在中央,形成单核胚囊(图版Ⅱ,7)。单核胚囊期顶花花蕾长4.00~5.00 cm,浅黄色花苞微张,子房长0.70~0.80 cm。当长大到一定程度时,功能大孢子细胞核进行连续的3次有丝分裂,先后形成二核胚囊(图版Ⅱ,8)、四核胚囊(图版Ⅱ,9),最终形成具有8个游离核的胚囊(图版Ⅱ,10)。最初胚囊的两端各有4个核。随后发生细胞化,在珠孔端的4个核有3个组成卵器,包括1个卵细胞和2个助细胞,第4个核成为上极核;合点端的4个核有3个形成反足细胞(图版Ⅱ,11、12)。第4个核作为下极核的功能,通常向上移动至上极核旁,构成含有2个核的中央细胞。这

表1 烤烟‘南江3号’顶花花蕾形态特征及其与雌雄蕊发育之间的关系
2个极核可以融合成为双倍的核,称为次生核。至此,形成7个细胞组成的胚囊。二核与四核胚囊时期顶花花蕾长5.00~6.80 cm,花瓣逐渐呈5裂,裂片粉红色,子房长0.80~0.90 cm。胚囊成熟时花蕾长5.00~6.80 cm,花瓣呈粉红色,子房长0.90~1.10 cm(表1)。
3讨论
目前国内有关茄科植物大、小孢子及雌雄配子发育方面报道主要集中在辣椒、番茄、酸浆以及迪勃纳氏烟草等研究上[8-11]。
‘南江3号’胚胎学主要特征表现为:花药壁的发育为基本型,绒毡层为腺质异型;小孢子母细胞减数分裂为同时型,成熟花粉为二细胞型;四分体排列方式有四面体型和十字交叉型两种;花粉具3或4个萌发孔。胚珠倒生,薄珠心;孢原体积较大,直接发育为大孢子母细胞,大孢子母细胞减数分裂形成的4个大孢子呈线性排列;胚囊发育为蓼型,成熟胚囊为7细胞8核。这与前人对茄科植物胚胎学特征的描述基本吻合,只是在成熟胚囊上与7细胞7核的辣椒不同[12]。同时说明烤烟‘南江3号’的大小孢子及雌雄配子发育非常保守。
‘南江3号’的雌蕊晚于雄蕊的发育,当雄蕊发育为四分体时期时雌蕊才开始发育。花蕾的大小是直观测定辨别雌雄蕊发育进程的形态特征,这为进行有关烟草杂交育种或遗传性状的改良提供理论和实际依据。花蕾发育到5.00~6.80 cm时胚囊才逐渐成熟,这与廖菊够等[12]描述的迪勃纳氏烟草品种有所差别。在实验过程中偶见花粉不育的现象,即萌发花粉管,花药不开裂。这类不育型虽然能形成正常花粉,但由于花药不开裂不能正常散粉,这可能与贵州栽烟季节连绵阴雨的气候环境有关。
参考文献:
[1]李智勇,韩晓红,谭建,等.烤烟新品种南江3号的选育及其特征特性[J].中国烟草科学,2009,30(4):1-5.
LI Z Y,HAN X H,TAN J,etal.Breeding and selecting of a new flue-cured tobacco variety Nanjiang 3 and its characteristics[J].ChineseTobaccoScience,2009,30(4):1-5.
[2]周立彬,周建云,刘国顺,等.烤烟新品种南江3号在贵州烟区的生态适应性研究[J].江西农业学报,2011,23(7):57-62.
ZHOU L B,ZHOU J Y,etal.Study on ecological adaptability of flue-cured tobacco new variety Nanjiang No.3 in Guizhou Province[J].ActaAgriculturaeJiangxi,2011,23(7):57-62.
[3]贺乘龙.烤烟品种南江3号关键栽培技术研究[D].长沙:湖南农业大学,2012:7-13.
[4]艾复清,许齐,刘洋州,等.不同变频风速对南江3号上部烟叶烤后质量的影响[J].贵州农业科学,2012,40(1):75-78.
AI F Q,XU Q,LIU Y Z,etal.Effect of fan speed on quality of flue-cured upper tobacco leaves of Nanjiang 3[J].GuizhouAgriculturalSciences,2012,40(1):75-78.
[5]高维常,瞿永生,袁有波,等.不同烤烟品种烟碱与钾含量变化及分类比较[J].江苏农业科学,2011,39(5):106-108.
GAO W C,QU Y S,YUAN Y B,etal.Nicotine and potassium content changes and classification comparison of different flue-cured tobacco varieties[J].JiangsuAgriculturalSciences,2011,39(5):106-108.
[6]靳涵丞,张洪亮,张军,等.贵州南江3号烟叶全氮含量高光谱估算模型[J].贵州农业科学,2015,43(3):91-95.
JIN H C,ZHANG H L,ZHANG J,etal.Total nitrogen content in leaves of Nanjiang 3 determined by hyperspectral estimation model in Guizhou[J].GuizhouAgriculturalSciences,2015,43(3):91-95.
[7]李祖任,胡楠,杨吉刚,等.繁缕和鹅肠菜的花维管束系统比较解剖学研究及其系统学意义[J].植物科学学报,2013,31(6):525-532.
LI Z R,HU N,etal.Floral vascular comparative anatomy ofStellariamediaandMyosotonaquaticumand its systematic significance[J].PlantScienceJournal,2013,31(6):525-532.
[8]申家恒,李伟,杨虹,等.辣椒大、小孢子发生与雌、雄配子体发育的研究[J].园艺学报,2007,(6):1 443-1 452.
SHEN J H,LI W,YANG H,etal.Studies on the megasporogenesis,microsporogenesis and development of female and male gametophyte ofCapsicumannuumL.[J].ActaHorticulturaeSinica,2007,(6):1 443-1 452.
[9]陈国菊,程玉瑾,吴定华,等.番茄大、小孢子的发生及雌、雄配子体的发育[J].华南农业大学学报,1999,(2):36-40.
CHEN G J,CHENG Y J,WU D H,etal.Megasporogenesis,microsporogenesis and female and male gametophyte development in tomato (Lycopersionesculentum)[J].JournalofSouthChinaAgriculturalUniversity,1999,(2):36-40.
[10]张从顺.酸浆花萼形态解剖学及孢子发生与发育研究[D].郑州:河南大学,2013:9-11.
[11]胡适宜.被子植物生殖生物学[M].北京:高等教育出版社,2005.
[12]廖菊够,康洪梅,代瑾然,等.烟草大小孢子发生和雌雄配子体发育研究[J].西北植物学报,2013,33(1):11-16.
LIAO J G,KANG H M,DAI J R,etal.Megasporogenesis,microsporogenesis and development of female and male gametophyte ofNicotianadebneyi[J].ActaBot.Boreal.-Occident.Sin.,2013,33(1):11-16.
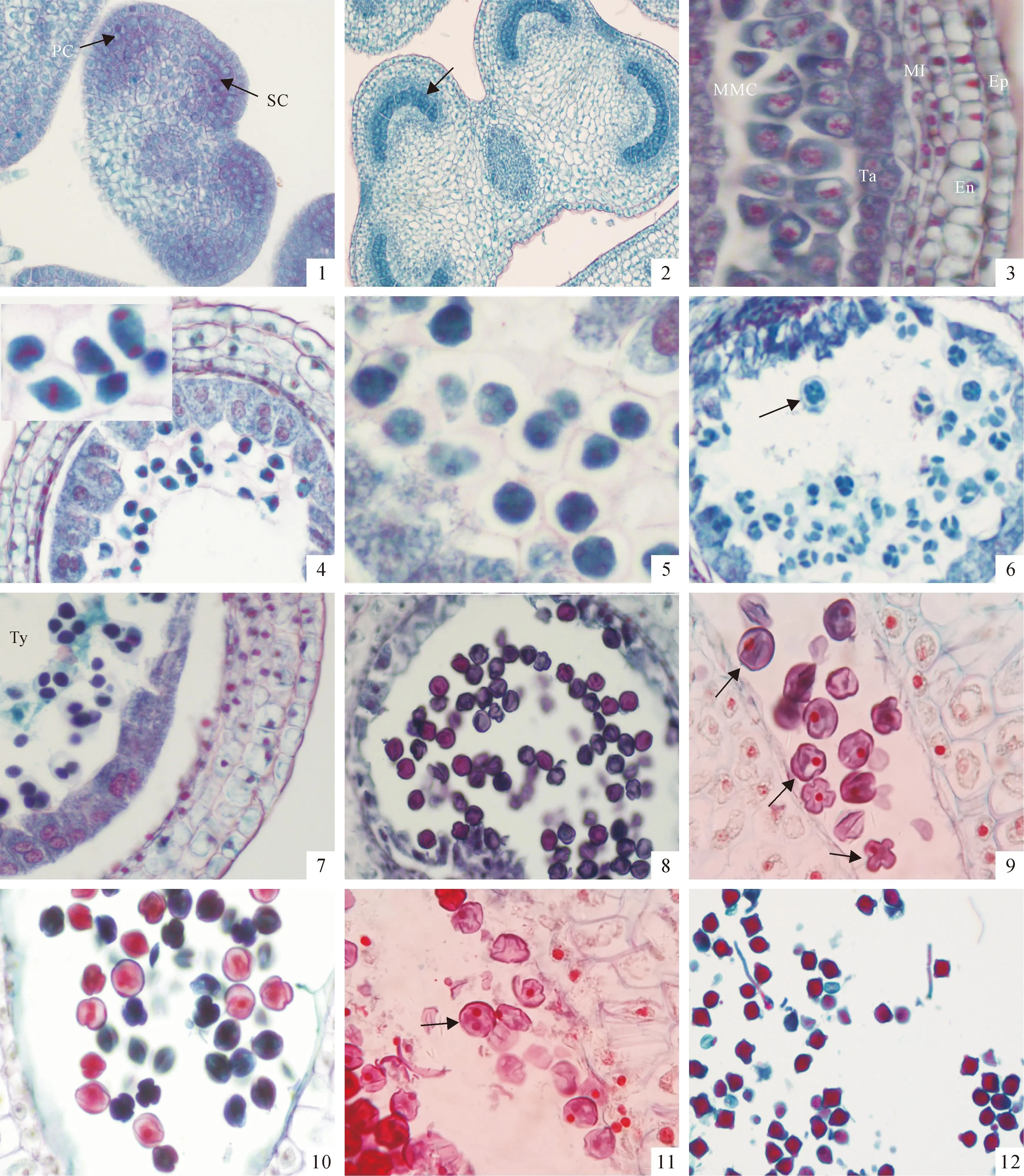
图版 Ⅰ‘南江3号’小孢子发生及雄配子体发育(1、3、5、8~11.×400;2、4、6、12.×200)
PC.周缘细胞;SC.造孢细胞;Ep.表皮;En.药室内壁;MI.中层;Ta.绒毡层;MMC.小孢子母细胞;Ty.四分体
1.孢原细胞平周分裂产生周缘细胞和初生造孢细胞;2.小孢子母细胞;3.小孢子母细胞准备减数分裂,绒毡层细胞质较浓厚;4、5.小孢子母细胞减数分裂时期;6.四分体时期;7.四分体排列方式为四面体型和十字交叉型;8单核花粉粒;9.单核中央期及靠边期;10、11.二细胞花粉;12.萌发花粉管。
Plate ⅠMicrosporogenesis and development of male gametophyte of flue-cured tobacco ‘Nanjiang 3’
PC.Peripheral cell;SC.Sporogenous cell;Ep.Epidermis;En.Endothecium;MI.Middle layer;Ta.Tapetum;MMC.Microspore mother cell;Ty.Tetrad
Fig.1.The archesporiumis divided into a parietal cell and a sporogenous cell,by the periclinal division;Fig.2.The microspore mother cell;Fig.3.The microspore mother cell prepare meiosis with intense tapetal cytoplasm;Fig.4,5.The meiosis period;Fig.6.The tetrad stage;Fig.7.The tetrad arrangement for tetrahedral or decussate tetrad;Fig.8.The monokaryotic microspore;Fig.9.Central and edge stage of the mononuclear pollen;Fig.10,11.The 2-celled pollen stage;Fig.12.The pollen tube elongation.
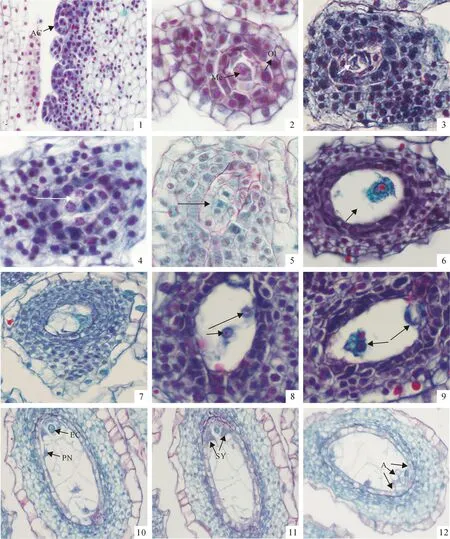
图版 Ⅱ‘南江3号’大孢子发生及雌配子体发育(1.×200;2~12.×400)
AC.孢原细胞;Me.大孢子母细胞;OI.珠被;EC.卵细胞;PN.极核;SY.助细胞;A.反足细胞
1.孢原细胞;2.大孢子母细胞;3.大孢子母细胞第一次有丝分裂,形成二分体;4.线性四分体早期;5.四分体晚期;6.功能大孢子;7.单核胚囊;8.二核胚囊;9.四核胚囊;10.成熟胚囊,示卵细胞、助细胞、极核、反足细胞;11.2个助细胞;12.3个反足细胞。
Plate ⅡMegasporogenesis and development of female gametophyte of flue-cured tobacco ‘Nanjiang 3’
AC.Archesportial cell;Me.Megaspore mother cell;OI.Outer integument;EC.Egg cell;PN.Polar nuclei;SY.Synergid;A.Antipodal cells
Fig.1.The archesportial cell;Fig.2.The megaspore mother cell;Fig.3.The megaspore mother cell forms dyad after first mitosis stage;Fig.4.The early linear tetrad;Fig.5.The late tetrad;Fig.6.The functional megaspore;Fig.7.The mononucleate embryonary sac;Fig.8.The bio- nucleate embryonary sac;Fig.9.The four- nucleate embryonary sac;Fig.10.Mature embryonary sac such as egg cell,synergid cell and polar nuclei and antipodal cell;Fig.11.Two synergid cells;Fig.12.Three antipodal cells.
(编辑:潘新社)
Megasporogenesis,Microsporogenesis and Development of Female and Male Gametophytes of Flue-cured Tobacco ‘Nanjiang 3’
GAO Kaiyue,JI Zhichao,LIAO Haimin*
(Collegs of Life Sciences,Guizhou University,Guiyang 550025,China)
Abstract:Flue-cured tobacco ‘Nanjiang 3’ was a kind of main cultivated varieties in Guizhou Province.Paraffin section method was employed to observe its anatomical structure of microsporogenesis,megasporogenesis and development of female and male gametophytes.The results showed that,the anther wall is defined as basic development type with glandular veriform tapetum.The cytoplasm of microspore mother cell undergoes simultaneous division and produces tetrahedral or decussate tetrad.The mature pollen belongs to 2 cell type and has three or four apertures.The ovary of flue-cured tobacco ‘Nanjiang 3’ is anatropous with thin nucellus.The archesporial cell develops to megaspore mother cell directly and has larger volume.Four megaspore cells form a linear tetrad.The development pattern of embryo sac is polygonum type,finally forms 7-cell or 8-cell nucleate mature embryo sac.Stamen develops earlier than gynoecium and the period of microsporogenesis,megasporogenesis and development of female and male gametophytes can be judged by appearance from the flower bud.
Key words:flue-cured tobacco;megasporogenesis;microsporogenesis;male and female gametophytes
中图分类号:Q944.58
文献标志码:A
作者简介:高凯悦(1991-),女,在读硕士研究生,主要从事结构植物学研究。E-mail:gaokaiyuelove@126.com*通信作者:廖海民,博士,教授,主要从事结构植物学研究。E-mail:lhaimin@163.com
基金项目:贵阳市现代农业与农村科技计划(筑科合同[2013102]4-6号);贵州大学研究生创新基金(2015014)
收稿日期:2015-09-29;修改稿收到日期:2015-12-14
文章编号:1000-4025(2016)02-0303-06
doi:10.7606/j.issn.1000-4025.2016.02.0303
猜你喜欢
湖北农机化(2020年4期)2020-07-24
活力(2019年19期)2020-01-06
中国烟草学报(2017年1期)2017-11-16
中国烟草学报(2017年1期)2017-11-16
作物研究(2014年6期)2014-03-01
作物研究(2014年6期)2014-03-01
中国烟草学报(2012年5期)2012-04-12
中国烟草学报(2012年3期)2012-04-10
中国烟草学报(2012年1期)2012-04-09
中国烟草学报(2012年4期)2012-04-09
