
基质栽培番茄临界氮浓度和氮营养指数研究
2016-03-23 02:57杜娅丹曹红霞谷晓博
节水灌溉 2016年9期
杜娅丹,曹红霞,谷晓博,王 萍
(西北农林科技大学旱区农业水土工程教育部重点实验室,陕西 杨凌 712100)
氮素是作物吸收的主要养分之一,缺氮会限制作物的生长,氮素过多会造成环境污染、氮肥利用效率低等问题,氮肥的供应与作物需求比例失调是农业生产中亟待解决的关键问题[1]。明确番茄不同生育阶段的氮需求特征是合理施肥的基础。临界氮浓度被定义为在一定生长时期内获得最大生物量时的最小氮浓度值[2],确定作物的临界氮浓度值是作物氮营养诊断的基本方法之一。近年来,国内外学者已针对小麦[3,4]、玉米[5]、籼稻[6]、番茄[7]、高粱[8]、冬油菜[9]等建立了不同作物的临界氮浓度稀释曲线模型,并利用模型对作物进行氮营养诊断。基于临界氮浓度,氮营养指数(N nutrition index,NNI)被定义为实际氮浓度与临界氮浓度的比值,因其对作物氮营养水平变化敏感,可以作为评价氮营养状况的一个可靠指标[10]。HU Da-Wei等[11]构建了马铃薯氮营养指数与氮肥利用效率、产量、光能利用效率及叶片参数间的关系,结果表明氮营养指数与相对产量、叶面积指数、叶片含氮量呈线性正相关关系,与光能利用效率呈二次曲线关系。Christos A.Dordas[12]研究了亚麻籽氮营养指数和氮肥利用效率的关系和利用氮营养指数估测产量的可行性。梁效贵等[13]探讨了氮营养指数评价玉米氮营养状况的可行性,并得到氮营养指数与相对氮累积量、相对地上部生物量和相对产量均具显著相关性。Ziadi等[5]也将氮营养指数作为玉米氮亏缺诊断的评价指标。而在不同浓度硝铵比营养液基质栽培番茄条件下,构建不同氮源番茄的临界氮浓度、氮营养指数和氮亏缺模型并诊断番茄氮营养状况,以及探究NNI和植株氮累积量、地上部生物量和产量间关系的研究还未见报道。
试验以“天硕308”番茄品种为供试材料,采用桶栽基质栽培技术,研究不同浓度硝铵比下番茄地上部生物量、植株含氮量的动态变化,建立基质栽培番茄不同浓度硝铵比的临界氮浓度稀释模型、氮营养指数模型和氮亏缺模型,并探讨氮营养指数和相对地上部生物量、相对氮累积量和相对产量间的关系,以及利用氮营养指数估测基质栽培番茄植株氮素盈亏水平的可行性,以期为基质栽培番茄氮素的合理施用提供理论依据和技术支持。
1 材料与方法
1.1 试验设计
试验于2014年8-12月在西北农林科技大学旱区农业水土工程教育部重点实验室日光温室内进行。供试番茄品种为“天硕308”。试验用桶底部直径25 cm,顶部30 cm,高30 cm,桶底部打孔,下垫有托盘,试验期间托盘内未观察到液体渗出,每桶装珍珠岩0.75 kg。于2014年8月18日选取长势一致,无病虫害的健壮植株定植于桶内,每桶1株。番茄全生育期可划分为苗期(8月18日至9月3日)、开花坐果期(9月4日至9月19日)、一穗果膨大期(9月20日至10月4日)、二穗果膨大期(10月5日至11月12日)、收获期(11月13日至12月2日),本试验中番茄品种为无限生长型,生长过程中留3穗果打顶。
营养液采用1/2Hoagland营养液,略作修改,即在营养液中加入不同形态的氮源(NH+4-N和NO-3-N),铵态氮由NH4Cl提供,硝态氮由NaNO3提供。采用硝铵比和施氮水平2因素随机试验方案,设置3个硝铵配比:100∶0,75∶25,50∶50,分别标记为N100,N75+A25和N50+A50;以Hoagland标准配方的氮浓度为一个剂量(记为1s),每个硝铵比组合分别设置3个氮浓度水平为(1/4)s,(1/2)s和(3/4)s,共9个处理,即N100(1/4)s、N100(1/2)s、N100(3/4)s、N75+A25(1/4)s、N75+A25(1/2)s、N75+A25(3/4)s、N50+A50(1/4)s、N50+A50(1/2)s和N50+A50(3/4)s,每个处理重复15次。营养液中大量元素的含量分别是P 0.5 mmol/L、K 3 mmol/L、Ca 2 mmol/L、Mg 1 mmol/L、S 1 mmol/L,微量元素的含量分别是 B 26.26 μmol/L-1、Mn 9.15 μmol/L、Zn 0.77 μmol /L、Cu 0.32 μmol /L、Mo 0.39 μmol/L,以EDTA-Fe(20.06 μmol/L)为Fe源。此外,营养液中均加入双氰胺(7 μmol/L)作为硝化抑制剂。营养液直接浇灌在基质中,苗期、开花坐果期和收获期每隔2 d浇灌一次营养液,每桶每次用量500 mL,一穗果膨大期和二穗果膨大期每隔1 d浇灌一次营养液,每桶每次用量500 mL。
1.2 测定项目及方法
分别于苗期、开花坐果期、一穗果膨大期、二穗果膨大期、收获期随机选取番茄3株,将样株去根,茎、叶、果分离,105 ℃下烘30 min杀青,然后于75 ℃烘干至恒质量称重。粉碎并充分混匀烘干样品,过0.5 mm筛,用H2SO4-H2O2消煮[14],采用AA3型流动分析仪测定消煮液中全氮含量。各器官氮累积量=各器官含氮量×各器官干物质量;所有器官氮累积量相加得到地上部植株氮累积量;地上部植株含氮量=地上部植株氮积累量/地上部植株干物质量。
果实成熟后按处理采摘,每个处理取3个重复,每次收获时将各计产处理分别称取果实鲜重,计算产量。
1.3 相关指标计算方法
1.3.1临界氮浓度
临界氮浓度的计算方法有2种。第1种是根据Justes等[15]1994年提出的临界氮浓度的定义及计算方法,步骤如下:①数据分组。作物生长对氮素的反应可以分为2种情形,一是随着施氮量的增加作物地上部生物量显著增加,表明作物生长受氮素不足的限制;二是尽管氮素吸收量可能增加,但地上部生物量并未出现显著增加,即作物生长不受氮素制约。对不同施氮处理的每次取样数据进行差异显著性分析,通过分析对作物生长受氮素营养限制与否的施氮水平进行分类。②对2组数据的地上部生物量(X)和对应的氮浓度值(Y)分别进行回归曲线拟合,每次取样的临界氮浓度由上述2类曲线的交点的纵坐标决定。第2种方法是Herrmann和Taube[16]为解决相邻的氮素处理水平结果相近、显著性不高而提出的,对生物量和含氮量之间的关系以分段函数拟合。而本试验不同处理间差异显著,且各取样时期数据未呈现较好的分段函数特性,因此采用第1种方法建模。
1.3.2临界氮浓度稀释曲线
依据Lemaire和Salette等[17]1984年提出的临界氮浓度与地上部生物量关系的方程式,临界氮浓度稀释曲线模型为:
%Nc=acDW-bmax
(1)
式中:%Nc为临界氮浓度值;ac为基质栽培番茄地上部生物量为1 g/株时的临界氮浓度;DWmax为番茄地上部生物量最大值,g/株;b为决定临界氮浓度稀释曲线斜率的统计学参数。
1.4 氮素吸收模型和地上部氮累积亏缺模型
番茄植株的氮吸收量与累积地上部最大生物量之间的关系可用如下公式表示:
Nupt=10NcDWmax
(2)
将式(1)代入式(2)得到番茄的临界氮吸收模型:
Nuptc=10acDW1-bmax
(3)
由式(2)可推出番茄的地上部氮累积亏缺模型:
Nand=Nna-Nuptc
(4)
式中:Nna为不同氮肥浓度下实际氮积累量,mg/株;Nuptc为临界氮浓度条件下的氮积累量,mg/株;Nand为氮亏缺量,若Nand>0,说明氮积累过量,反之则氮积累不足。
1.5 氮营养指数(NNI)
为了进一步明确作物的氮素营养状况,Lemaire等提出了氮素营养指数(N nutrition index,NNI)的概念,可用如下公式来表示:
NNI=Nt/Nc
(5)
式中:Nt为地上部生物量氮浓度的实测值,mg/g;Nc为根据临界氮浓度稀释曲线模型求得的在相同地上部生物量时的氮浓度值,mg/g。
NNI可以直观反映作物体内的氮营养状况,NNI小于1,则氮素不足;等于1,恰好合适;大于1,则氮营养过剩。
1.6 数据处理与分析
由于番茄产量、地上部生物量、氮累积量随生育期变化较大,为了量化产量、地上部生物量、氮累积量与NNI间的关系,将不同氮源下的这些指标分别进行处理:
RY=Ya/Ym
(6)
RNupt=Na/Nm
(7)
RDW=DWa/DWm
(8)
式中:RY为相对产量;Ya为各处理实测产量;Ym为平均产量;RNupt为相对氮累积量;Na为各处理实际氮累积量;Nm为平均氮累积量;RDW为相对地上部生物量;DWa为各处理实测地上部生物量;DWm为平均地上部生物量。
数据采用DPS软件进行试验统计分析,方差分析使用最小显著差异法(LSD)进行,所有数据采用OriginPro8.5作图。
2 结果与分析
2.1 基质栽培番茄临界氮浓度稀释曲线模型的建立
2.1.1不同浓度硝铵比下番茄地上部生物量与氮浓度变化规律
分析不同浓度硝铵比下番茄地上部生物量和相对应的氮浓度值(见表1和图1)。结果表明,不同氮源处理下,地上部生物量随生育进程而增加,且在N75+A25 (1/2)s处理下达到最大值153.80 g/株;同一氮源同一取样时期,番茄地上部生物量随着施氮浓度的增加,表现为先增加后降低的趋势,并以(1/2)s处理最高,说明过量施氮没有有效提高地上部生物量。由图1可知,不同氮源下地上部生物量中的氮浓度随着氮肥浓度的增加而增加,随生育进程而降低,说明番茄地上部氮浓度存在稀释现象。而在相同氮肥浓度下, N75+A25番茄地上部生物量的氮浓度值均高于N100和N50+A50处理对应的氮浓度值。
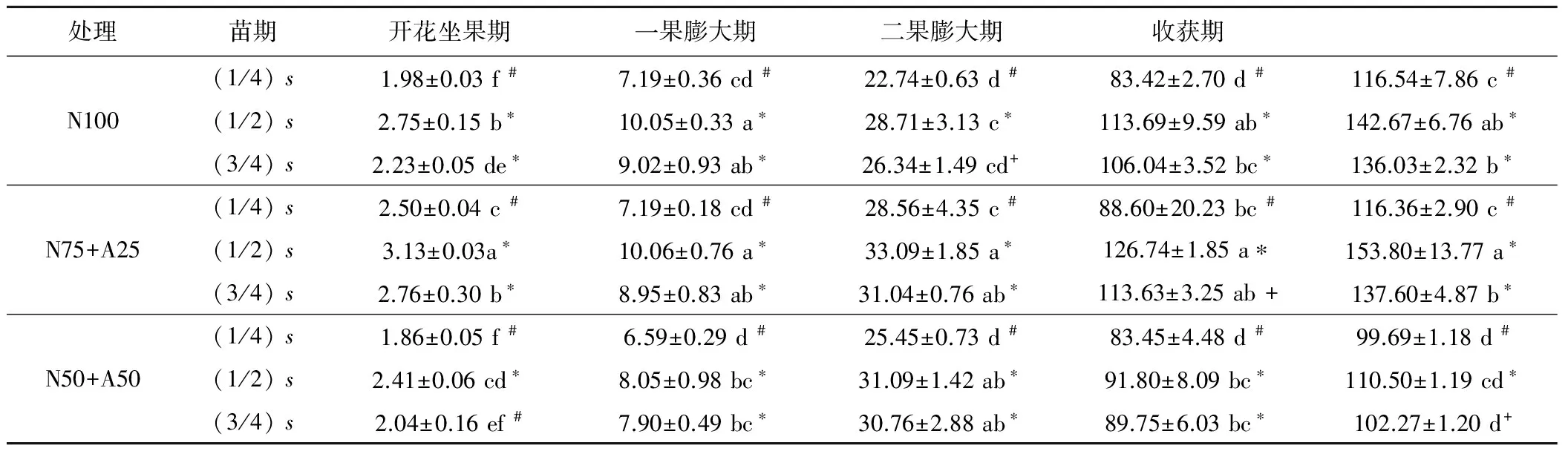
表1 不同浓度硝铵比对番茄地上部生物量的动态累积的影响Tab.1 Effects of different concentrations of nitrate and ammonium ratios on dynamic accumulation of soilless tomato shoot biomass
注:表中数据为平均值±标准误差;同列不同小写字母表示在0.05水平上差异显著;#表示数据为限氮组;*表示数据为非限氮组;+表示该数据与限氮和非限氮数据均不显著。
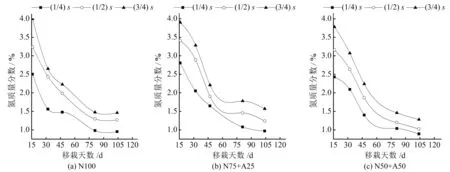
图1 不同浓度硝铵比下基质栽培番茄地上部生物量氮浓度的动态变化Fig.1 Dynamic of nitrogen concentration in aboveground biomass of soilless tomato under different concentrations of nitrate and ammonium ratios
根据Justes等[15]提出的临界氮浓度的计算方法,对各生育期不同浓度硝铵比下番茄的地上部生物量进行LSD方差分析(见表1)。根据分析结果将数据分为限氮(标记为#)和非限氮(标记为*)2组,即每个取样日的最低生物量等级的为限氮组,每个取样日最高生物量等级的为非限氮组,与限氮组和非限氮组差异均不显著(标记为+,即一果膨大期的N100 (3/4)s、二果膨大期的N75+A25 (3/4)s、收获期的N50+A50 (3/4)s处理的数据)的数据点予以舍去。从不同氮源下地上部生物量的方差分析结果可以看出,非限氮组的数据点均来自(1/4)s浓度的取样值,限氮组的数据点几乎全部为(1/2)s和(3/4)s处理的取样值。
2.1.2临界氮浓度稀释曲线的建立
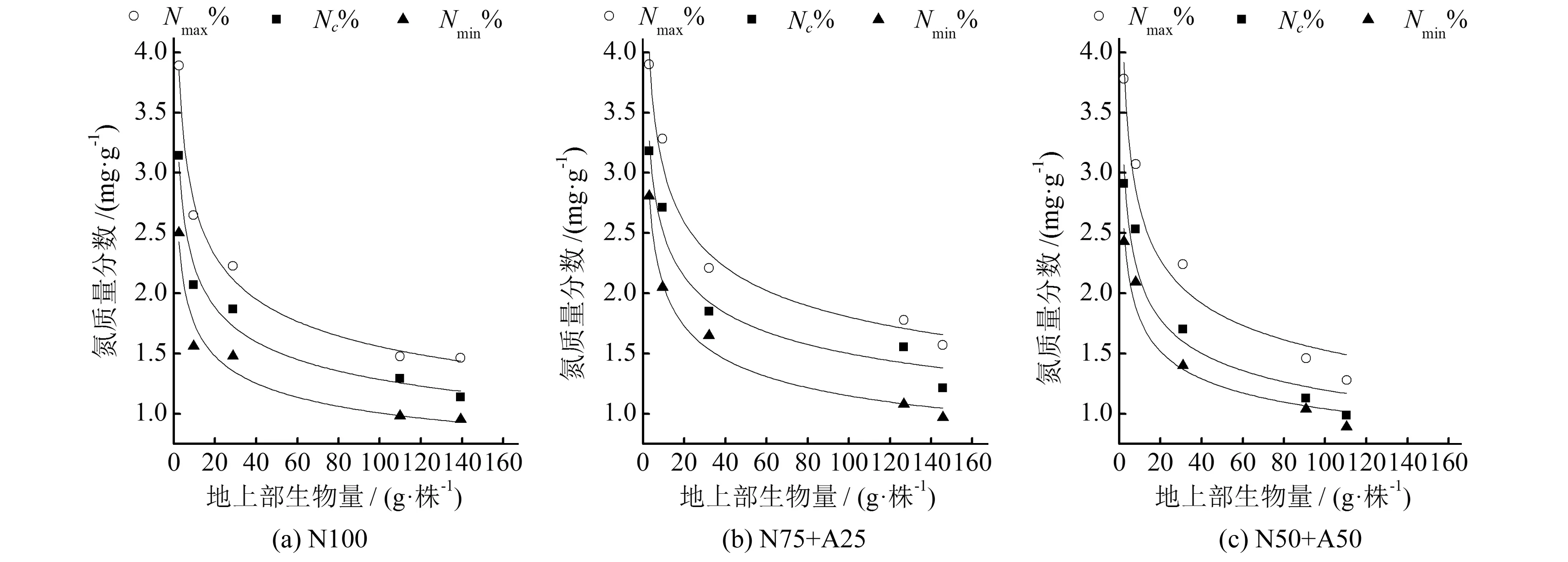
根据式(1)确定的临界氮浓度值与对应的地上部最大生物量进行拟合,得到番茄临界氮浓度稀释曲线模型。根据式(3)计算得到氮素吸收模型(见表2)。通过番茄地上部生物量与氮浓度值的对比发现,在相同地上部生物量的情况下,氮浓度值有很大的变异性,利用每个取样日所观察到氮浓度的最大值、最小值可得到番茄最大、最小氮浓度稀释模型,同样符合模型(1),模型参数见表2。
从表2可以看出,不同氮源下番茄临界、最高、最低氮浓度稀释模型形式一致,但参数不同。同一氮源下模型的曲线斜率b值相同或相近,而不同氮源间模型斜率差异较大,表现为N75+A25 表2 番茄临界氮浓度稀释模型和氮素吸收模型参数值Tab.2 Parameters of nitrogen dilution model and nitrogen uptake model for tomato 2.2.1基于氮营养指数(NNI)的基质栽培番茄氮营养诊断 为了更精确地反映番茄植株氮素是否适宜,氮营养指数被用来定量评估番茄体内的氮素状况。由图3可知,不同浓度硝铵比下的NNI变化趋势一致,都呈现出一定程度的波动性。番茄移栽后32~84 d是番茄开花坐果期至二果膨大期,(1/2)s和(3/4)s处理的NNI总体呈现下降的趋势,可能是由于地上部生物量的快速增加,促使植株对氮肥需求量增加。N100、N75+A25和N50+A50 3种氮源处理下,随着施氮水平的提高NNI的值不断上升,且(1/4)s处理的NNI都小于1,表明番茄受到了氮素不足的制约,(3/4)s处理的NNI都大于1,表明番茄氮营养过剩;(1/2)s处理下,3种氮源的NNI分别为1.003~1.180、0.99~1.070、1.040~1.110,N50+A50的适宜施氮水平在(1/4)s和(1/2)s之间变化,N100下(1/2)s施肥水平与适宜施氮量最为接近,但仍略高于需求量,可见N100下适宜的施肥水平应该是略低于(1/2) s浓度,N75+A25 (1/2)s处理的NNI始终最接近1或在1附近变化,表明此时的施氮量较为适宜。 图2 基质栽培番茄地上部生物量的氮浓度稀释曲线Fig.2 Nitrogen concentration dilution curve in aboveground biomass of soilless tomato 图3 不同浓度硝铵比下基质栽培番茄氮营养指数动态变化Fig.3 Dynamics of the NNI for soilless tomato under different concentrations of nitrate and ammonium ratios 2.2.2基于氮累积亏缺的基质栽培番茄氮营养诊断 N100、N75+A25和N50+A50 3种氮源处理下,随着施氮浓度的提高,氮亏缺值均逐渐减小(见表3)。由表3可知,在番茄生长过程中氮亏缺值呈现一定的动态变化,将不同浓度硝铵比下实测氮亏缺值与最佳氮营养状况之间的偏离程度进行统计分析,各处理下动态氮亏缺值与最佳营养状况间的RMSE,N100分别为0.551、0.151、0.272 g/株,N75+A25分别为0.597、0.068、0.286 g/株,N50+A50分别为0.312、0.169、0.295 g/株,得到N75+A25 (1/2)s的动态氮亏缺值与最佳氮营养状况的RMSE最小,说明硝铵比75∶25,施肥水平(1/2)s为最佳的施肥组合。 由图4可知,3种氮源处理下番茄氮营养指数和氮亏缺值呈极显著正相关关系,拟合度分别为0.903、0.920和0.890[式(9)、(10)、(11)]。整个生育期中,氮营养指数从0.7到1时,根据公式(9)、(10)、(11)得到N100、N75+A25和N50+A50分别需要累积0.539、0.618和0.409 g/株氮素,说明N75+A25氮源组合需要比N100和N50+A50积累更多的氮素才能满足作物正常生长的需要。 表3 不同浓度硝铵比下基质栽培番茄氮亏缺值的动态变化Tab.3 Dynamics of the accumulative nitrogen deficit for soilless tomato under different concentrations of nitrate and ammonium ratios 氮营养指数和氮亏缺值定量关系: N100:Nand=1.796NNI-1.925R2=0.920** (9) N75+A25:Nand=2.059NNI-2.193R2=0.920** (10) N50+A50:Nand=1.362NNI-1.411R2=0.818** (11) 图4 番茄氮营养指数与氮亏缺值的定量关系Fig.4 Quantitative relationship between NNI and accumulative nitrogen deficit for tomato 为了评价NNI在预测植株氮素盈亏上的可行性,分别研究了相对氮累积量(RNupt)、相对地上部生物量(RDW)和相对产量(RY)与NNI之间的关系(见图5~7)。结果表明,相对地上部生物量和产量均随NNI的增加呈先升高后降低的趋势,RDW~NNI、RY~NNI均表现为二次曲线关系,除N50+A50下相对产量与NNI为显著二次相关外,其他均达极显著水平。而相对氮累积量随着NNI的增加而增加,RNupt~NNI表现为线性相关关系,拟合度均达极显著水平。本研究条件下,各氮源下番茄的相对地上部生物量和相对产量达到最大值的NNI值,在N100处理下分别为1.085、1.085,在N75+A25处理下分别为1.079、1.051,在N50+A50处理下分别为1.137、1.120,均处在1~1.14。以上分析表明,NNI和相对氮累积量、相对地上部生物量、相对产量之间关系密切,利用氮营养指数定量评估番茄体内的氮营养状况是可靠的。 图5 番茄氮营养指数(NNI)与相对地上部生物量(RDW)的关系Fig.5 Relationship between NNI and relative aboveground biomass in tomato 图6 番茄氮营养指数(NNI)与相对氮累积吸收量(RNupt)的关系Fig.6 Relationship between NNI and relative total N uptake amount in tomato 图7 番茄氮营养指数(NNI)与相对产量(RY)的关系Fig.7 Relationship between NNI and relative yield (RY) in tomato 本研究中番茄地上部生物量的氮浓度值随着施肥量的增加而增加,随生育进程稀释的现象可能是由于植株生长过程中叶片的相互遮阴以及茎、果实和叶柄的比例增大有关[20],这与Justes[21]和王新[7]的研究结果一致。国内外学者构建了不同作物的临界氮浓度稀释模型,Justes[21]等得到冬小麦临界氮浓度稀释曲线模型%Nc=5.35DW-0.442max,Caludia M[22]等构建加工马铃薯的临界氮浓度稀释曲线模型%Nc=5.30DW-0.42max,Tei F[23]构建加工番茄临界氮浓度稀释曲线模型%Nc=4.53DW-0.327max,上述模型形式与Greenwood[24]等提出的模型形式一致,临界氮含量与地上部生物量之间均表现为幂函数关系。本研究所构建的不同氮源下基质栽培番茄地上部生物量和临界氮浓度值间的模型形式符合Greenwood[21]等的假设:%Nc=3.836DW-0.237max(N100)、%Nc=4.145DW-0.221max(N75+A25)、%Nc=3.470DW-0.247max(N50+A50),但参数不同,原因可能是:①不同作物和品种的根、茎、叶形态及其内在生理生态机制不同造成了相应的氮浓度稀释模型参数差异较大;②由于施氮方式、栽培条件及品种的不同造成了模型参数的差异,王新[7]等采用里格尔“87-5”番茄品种,膜下滴灌栽培条件直接施入氮肥的方式构建加工番茄的临界氮浓度稀释曲线模型,Tei F基于大田条件构建了加工番茄临界氮浓度稀释曲线模型,模型仅适用于移栽后40 d到成熟期。而本研究采用桶栽基质栽培、浇灌营养液的栽培模式,构建了不同氮源下番茄临界氮浓度稀释模型,能够更精确地量化氮肥的施入量和作物的氮素营养状况,使模型更具机理性。 基于作物临界氮浓度,Lemaire等[25]提出的NNI的概念,具有明确的生物学意义,可以动态诊断番茄氮营养状况。本研究中运用NNI对不同氮源下番茄氮营养状况进行诊断,得到与其他指标确定的适宜施肥量相同的结论,表明NNI可以较好地反应番茄生育期的氮营养状况,量化了番茄在生长过程中的养分供应量。通过进一步分析发现,NNI和番茄氮亏缺值、相对氮累积量、相对地上部生物量和产量间显著相关,说明利用NNI评价植株氮营养状况的方法是可靠的。 (1)本文依据不同浓度硝铵比营养液基质栽培试验资料,建立了3个氮源下番茄临界氮浓度稀释模型%Nc=3.836DW-0.237max(N100)、%Nc=4.145DW-0.221max(N75+A25)、%Nc=3.470DW-0.247max(N50+A50),3个模型的参数ac和曲线斜率b均不相同,表明不同氮源处理番茄有其独立的氮浓度稀释曲线。 (2)基于临界氮浓度构建的氮营养指数和氮亏缺模型对基质栽培番茄生育期的氮营养状况诊断结果一致,以N75+A25 (1/2)s最适宜。 (3)氮营养指数能直观反映番茄生育期的氮素营养状况,与氮亏缺值、相对地上部生物量、相对氮累积量和相对产量间呈显著相关性,可为基质栽培番茄精确合理化施用氮肥提供理论指导。 [1] Haraldsen T K, Asdal A, Grasdalen C, et al. Nutrient balances and yields during conversion from conventional to organic cropping systems on silty loam and clay soils in Norway [J]. Biological Agriculture and Horticulture, 2000,17:229-246. [2] Lemaire G, Gastal F, Salette J. Analysis of the effect of N nutrition on dry matter yield of a sward by reference topotential yield and optimum N content [C]∥ Proceedings of the 16th International Grassland Congress. Nice, France, 1989:179-180. [3] Yue S C, Meng Q F, Zhao R F, et al. Critical nitrogen dilution curve for optimizing nitrogen management of winter wheat production in the North China Plain [J]. Agronomy Journal, 2012,104:523-529. [4] Ziadi N, Belanger G, Claessens A, et al. Determination of a critical nitrogen dilution curve for spring wheat [J]. Agronomy Journal, 2010,102:241-250. [5] Ziadi N, Brassard M, Bélanger G, et al. Critical nitrogen curve and nitrogen nutrition index for corn in eastern Canada [J]. Agronomy Journal, 2008,100: 271-276. [6] Sheehy J E, Dionora M J A, Mitchell P L, et al. Critical nitrogen concentrations: implications for high-yielding rice (Oryza sativa L.) cultivars in the tropics [J]. Field Crops Research, 1998,59:31-41. [7] 王 新, 马富裕, 刁 明,等. 滴灌番茄临界氮浓度、氮素吸收和氮营养指数模拟[J]. 农业工程学报, 2013,29(18):99-108. [8] van Oosterom E J, Carberry P S. Critical and minimum N contents for development and growth of grain sorghum [J]. Field Crops Research, 2001,70(1):55-73. [9] Colnenne C, Meynard J M, Reau R. Determination of a critical nitrogen dilution curve for winter oilseed rape [J]. Annals of Botany, 1998,81(2):311-317. [10] Lemaire G, Jeuffroy M H, Gastal F. Diagnosis tool for plant and crop N status in vegetative stage theory and practices for crop N management [J]. European Journal of Agronomy, 2008,28:614-624. [11] Hu D W, Sun Z P, Li T L, et al. Nitrogen nutrition index and its relationship with N use efficiency, tuber yield, radiation use efficiency, and leaf parameters in potatoes [J]. Journal of Integrative Agriculture, 2014,13(5):1 008-1 016. [12] Christos A D. Nitrogen nutrition index and its relationship to N use efficiency in linseed [J]. European Journal of Agronomy, 2011,34:124-132. [13] 梁效贵,张经廷,周丽丽, 等. 华北地区夏玉米临界氮稀释曲线和氮营养指数研究[J]. 作物学报, 2013,39(2):292-299. [14] 鲁如坤. 土壤农业化学分析方法[M]. 北京:中国农业科技出版社, 2000:302-316. [15] Justes E, Mary B, Meynard J M, et al. Determination of a critical nitrogen dilution curve for winter wheat crops [J]. Annals of Botany, 1994,74:397-407. [16] Herrmann A, Taube F. The range of the critical N dilution curve for maize can be extended until silage maturity [J]. Agronomy Journal, 2004,96:1 131-1 138. [17] Lemaire G, Salette J. Croissance estivale en matiere seche de peuplements de fetuque elevee (Festuca arundinacea Schreb) et de dactyle (Dactylisglomerata L.) danslouest de la France: I. Etude en conditions de nutrition azotee et dalimentation hydrique non limitantes [J]. Agrono-mie, 1987,7(6):373-380. [18] Dong C X, Shen Q R, Wang G. Tomato growth and organic acid changes in response to partial replacement of NO3-N by NH+4-N [J]. Pedosphere, 2004,14(2):159-164. [19] Yan F, Schubert S, Mengel K. Effect of low root medium pH on net proton release, root respiration, and root growth of corn (Zeamays L.) and broad bean (Viciafaba.) [J]. Plant Physiological, 1992,99:415-421. [20] Roger N, Yves D, Nicolas T. A mathematical model for nitrogen demand quantification and a link to broccoli (Brassica oleracea var. italica) glutamine synthetase activity [J]. Plant Science, 2003,165:483-496. [21] Justes E, Mary B, Meynard J M, et al. Determination of a critical nitrogen dilution curve for winter wheat crops [J]. Annals of Botany, 1994,74:397-407. [22] Claudia M, Giletto & Hernán E, Echeverría. Critical nitrogen dilution curve for processing potato in Argentinean Humid Pampas [J]. American Journal of Potato Research, 2012,89(2):102-110. [23] Tei F, Benincasa P, Guiducci M. Critical nitrogen concentration in processing tomato[J]. European Journal of Agronomy, 2002,18(1):45-55. [24] Greenwood D J, Gastal F, Lemaire G. Growth rate and %Nof field grown crops: theory and experiments [J]. Annals of Botany, 1991,67(2):181-190. [25] Lemaire G, Gastal F. N uptake and distribution in plant canopies: diagnosis of the nitrogen status in crops[R]. Springer Berlin Heidelberg, 1997:3-43.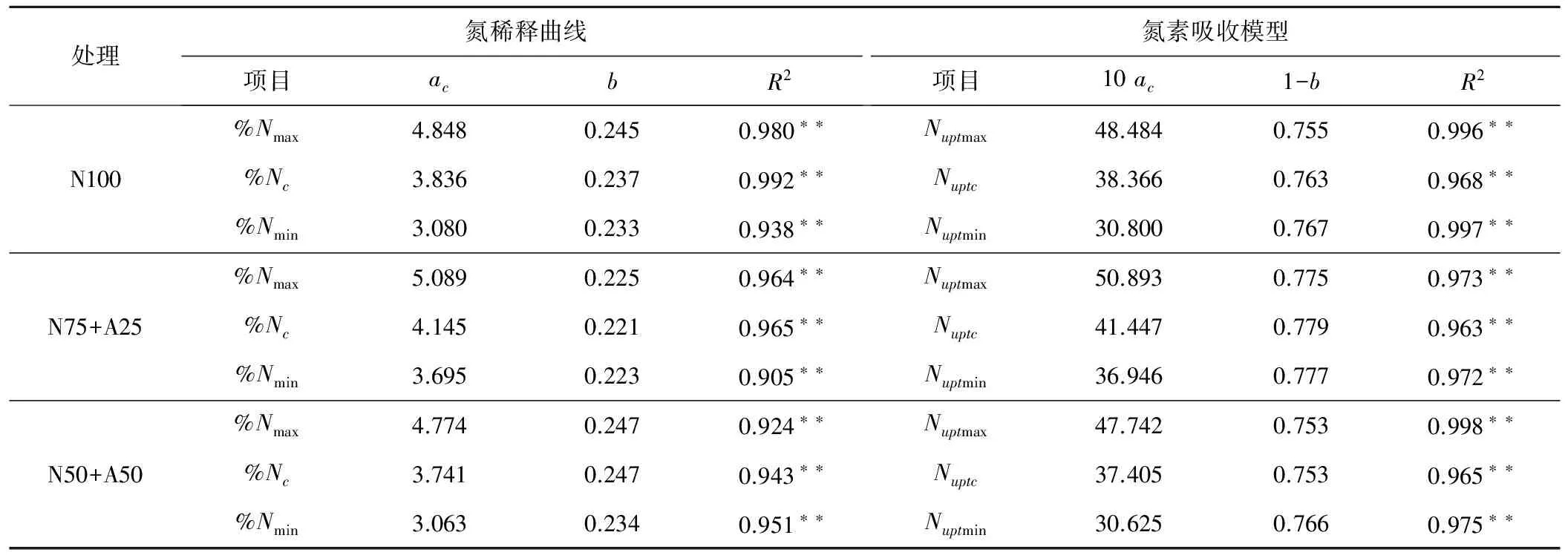
2.2 不同浓度硝铵比下番茄氮营养诊断
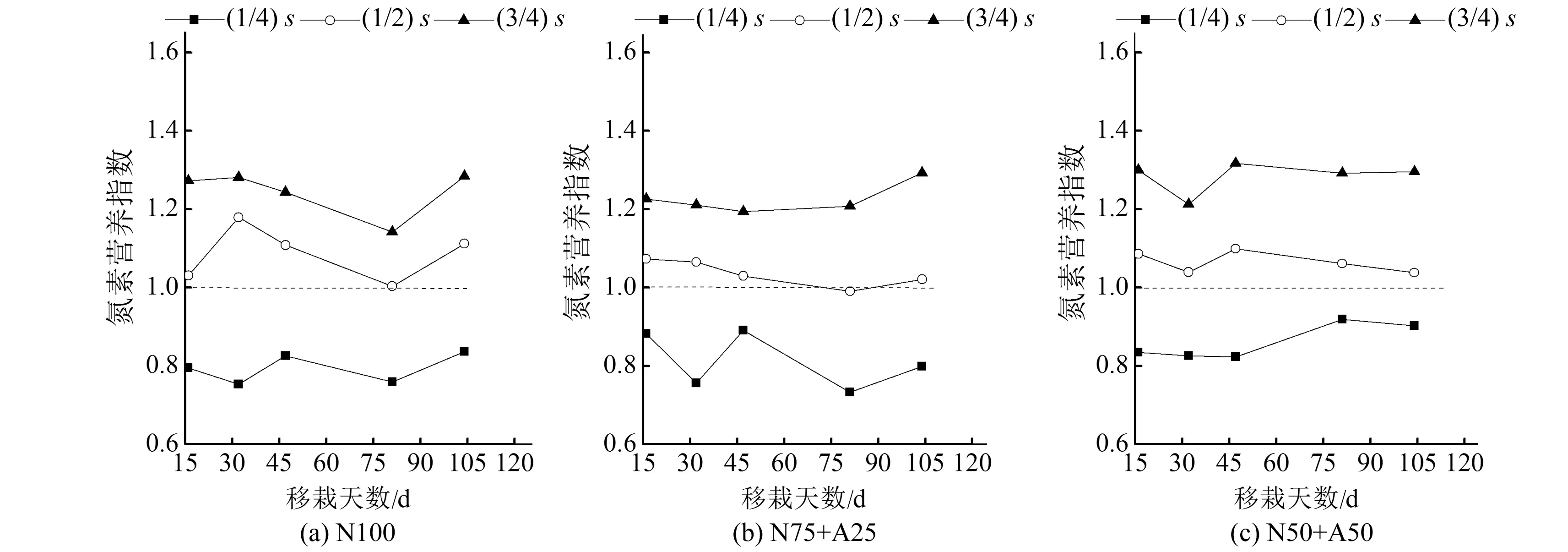

2.3 氮营养指数与相对氮累积量、相对地上部生物量和相对产量间的关系
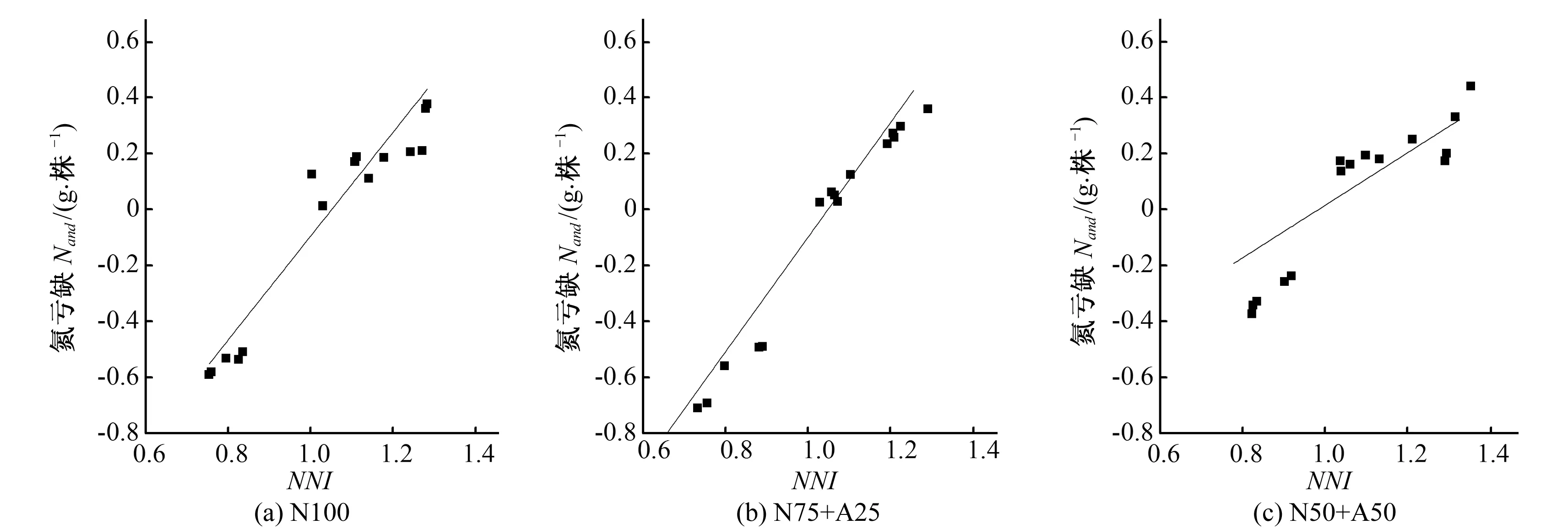

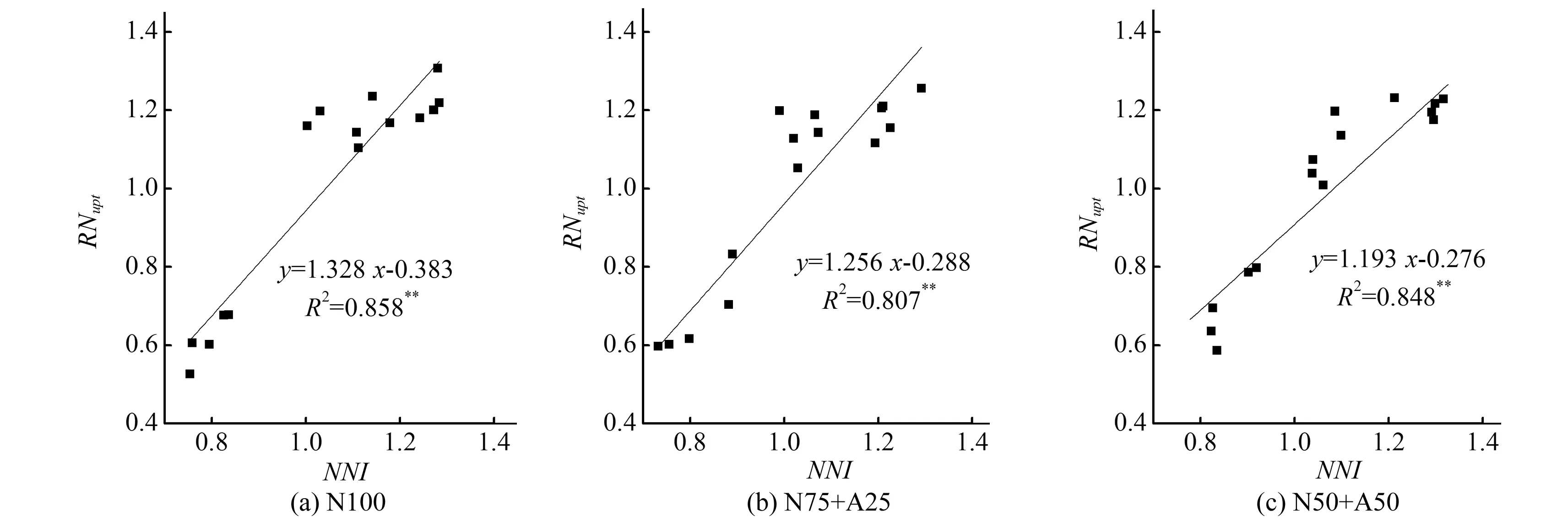

3 讨 论
4 结 论
猜你喜欢
中国农资(2021年10期)2021-12-14
山东林业科技(2019年2期)2019-06-03
中国酿造(2016年12期)2016-03-01
化工管理(2015年3期)2015-03-23
中国酿造(2014年9期)2014-03-11
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年3期)2011-10-24
植物营养与肥料学报(2011年6期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
