
长江流域棉花黄萎病菌的致病力多样性和遗传多样性分析
2016-03-16 09:00金利容万鹏黄薇杨晓华
湖北大学学报(自然科学版) 2016年2期
金利容,万鹏,黄薇,杨晓华
(湖北省农业科学院植保土肥研究所,湖北 武汉 430079)
长江流域棉花黄萎病菌的致病力多样性和遗传多样性分析
金利容,万鹏,黄薇,杨晓华
(湖北省农业科学院植保土肥研究所,湖北 武汉 430079)
摘要:棉花黄萎病是棉花上的重要病害,严重威胁着棉花的产量.从长江流域4个省即湖北省、湖南省、江西省和安徽省采集300多个菌株,选择各省不同县市的67个菌株,对这67个供试菌株进行培养性状的鉴定、致病力的测定以及遗传多样性的分析,结果表明,在67个供试菌株中,就致病力类型而言,强致病力类型、中等致病力类型和弱致病力类型分别占35.82%、43.28%和17.91%.就培养性状类型而言,菌核型、中间型和菌丝型分别占38.81%、22.39%和38.81%.菌株的致病力和菌落培养类型均与地理位置没有相关性,但菌株的致病力与培养性状类型之间存在一定的相关性,强致病菌株大多数为菌核型,而弱致病菌株则以菌丝型为主.利用筛选出的15对RAPD引物对供试菌株DNA扩增并进行遗传多样性的分析结果表明,遗传多样性与菌株来源地无显著相关性,同时,菌株的遗传多样性与菌株致病力和培养性状也没有显著相关性.
关键词:长江流域; 棉花黄萎病菌; 培养性状;致病力多样性; 遗传多样性
0引言

目前,DNA 分子标记技术已成为病原菌遗传多样性、致病力变异等研究的重要手段.RAPD(random amplified polymorphic DNA)全称为随机扩增多态性DNA,即利用不同的随机引物对基因组DNA进行扩增,所扩增的DNA片段的多态性反映了基因组相应区域的多态性,具有简单快速,所需DNA的量少等特点.
我们在长江流域4个不同的省份即江西省、安徽省、湖南省和湖北省的各个地区采集黄萎病病株,分离300多个菌株并进行单孢纯化,根据不同的县市和乡镇选取其中的67个菌株,对其进行培养性状类型的鉴定、致病力的测定以及遗传多样性的分析,从而明确长江流域棉花黄萎病菌致病力多样性、菌落形态多样性以及遗传变异性,并探讨它们相互之间的关系.
1材料与方法
1.1供试菌种的分离和纯化供试菌种分别为从长江流域4个省份湖北省、湖南省、江西省和安徽省各个县市的城镇和乡村的杂交棉上采集的病株,进行分离和单孢纯化后得到的菌株300多株,根据不同的县市和乡镇选择67个菌株作为供试菌株.
1.3供试菌种致病力的测定用于致病力测定的品种为抗病品种鲁棉28、耐病品种豫棉21和感病品种冀棉11.供试菌株包括67个菌株和2个对照菌株,其中对照菌株V991b为华中农业大学张献龙老师提供,属强致病力菌株,培养性状类型为菌核型.对照菌株AY为中国农科院棉花所朱荷琴老师提供,属安阳中等致病力菌株,培养性状类型为菌核型.
根据调查结果,计算病情指数.

表1 致病类型划分标准
1.5引物筛选和PCR扩增PCR反应所用随机引物购自上海生工生物技术工程公司.本试验选用70种随机引物,对这70种引物用4种菌株DNA进行筛选,选取出能扩增出清晰条带的随机引物,为本试验的RAPD分析扩增引物.反应体系总量为25 μL,模板DNA 1 μL,引物1 μL(浓度为10.0 μmol/L),dNTP 0.5 μL(浓度为25.0 mmol/L),TaKaRa Taq酶(1 u/μL)1 μL,PCR缓冲液2.5 μL,加超纯水至25 μL.扩增循环为94 ℃预变性3 min,94 ℃变性45 s,36 ℃退火1 min,72 ℃延伸3 min,40个循环,72 ℃延伸10 min.
1.6结果检测和分析扩增产物用1.5%的琼脂糖凝胶进行电泳,利用凝胶成像系统照相并保存.记录图谱中的条带,出现即为“1”,无条带即为“0”,制成图表用于数据处理,利用Ntsys软件计算各个菌株之间的相似系数,以UPGMA法进行聚类并生成树状图.
1.7特异性引物的扩增采用已报道的一对特异性引物[5]:上游引物D1(5’-CATGTTGCTCTGTT-GAACTGG-3’)和下游引物D2(5’-GACACGGTATCTTTGCTGCTGAA-3’),引物序列由北京鼎国生物公司合成.
PCR反应体系总量为25 μL,反应体系为:模板DNA 1 μL,上游/下游引物1 μL(浓度为10.0 μmol/L),dNTP 0.5 μL(浓度为25.0 mmol/L),Taq酶(1 u/μL)1 μL,PCR缓冲液2.5 μL,加超纯水至25 μL.扩增循环为94 ℃预变性5 min,94 ℃变性1 min,56 ℃退火1 min,72 ℃延伸3 min,30个循环,74 ℃延伸10 min.对以上菌株DNA进行特异性扩增,检测其结果.
2结果与分析
2.1供试菌种培养性状和致病力的鉴定对各供试菌株培养性状和致病力的测定结果进行归纳总结,所得结果见表2.
对表2的数据进行统计分析,结果发现,强、中等和弱致病力菌株在4个省份均匀分布,菌核型、菌丝型和中间型3种培养类型也在4个省份均匀分布.67个菌株划分为3个致病力类型,属强致病力类型的有24个,占35.82%,在这24个强致病力菌株中包括13个菌核型,8个中间型,3个菌丝型,主要以菌核型为主;属中等致病力类型的有29个,占43.28%, 在这29个中等致病力菌株中包括11个菌核型,5个中间型,13个菌丝型;属弱致病力类型有14个,占 17.91%,在这14个弱致病力菌株中无菌核型,2个中间型,12个菌丝型,以菌丝型为主.
2.2棉花黄萎病菌DNA的提取DNA提取试剂盒提取了69个菌株的DNA,经检测电泳条带清晰.

表2 供试菌株名称、来源、培养性状类型以及致病力类型的鉴定结果
续表2
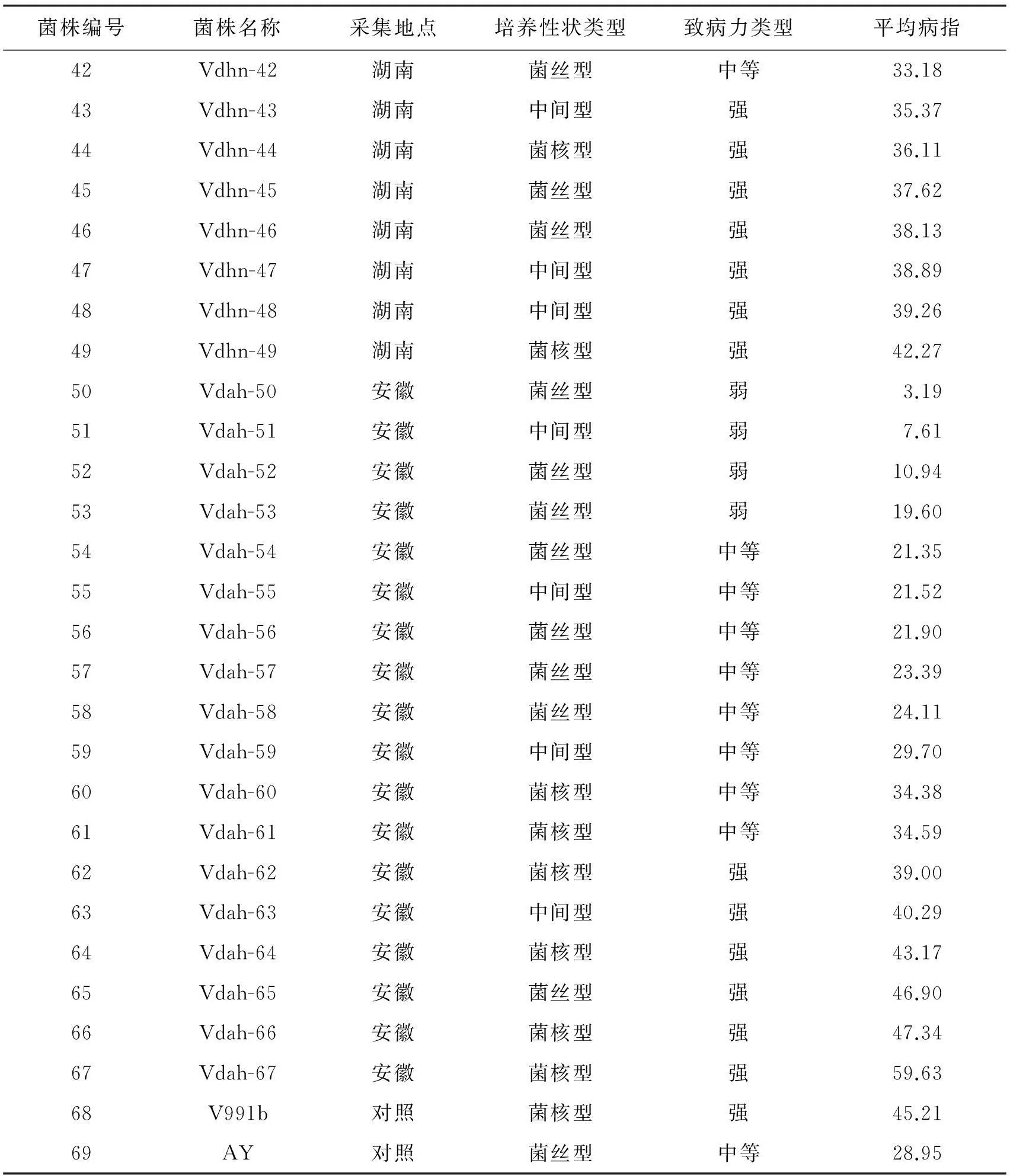
菌株编号菌株名称采集地点培养性状类型致病力类型平均病指42Vdhn-42湖南菌丝型中等33.1843Vdhn-43湖南中间型强35.3744Vdhn-44湖南菌核型强36.1145Vdhn-45湖南菌丝型强37.6246Vdhn-46湖南菌丝型强38.1347Vdhn-47湖南中间型强38.8948Vdhn-48湖南中间型强39.2649Vdhn-49湖南菌核型强42.2750Vdah-50安徽菌丝型弱 3.1951Vdah-51安徽中间型弱 7.6152Vdah-52安徽菌丝型弱10.9453Vdah-53安徽菌丝型弱19.6054Vdah-54安徽菌丝型中等21.3555Vdah-55安徽中间型中等21.5256Vdah-56安徽菌丝型中等21.9057Vdah-57安徽菌丝型中等23.3958Vdah-58安徽菌丝型中等24.1159Vdah-59安徽中间型中等29.7060Vdah-60安徽菌核型中等34.3861Vdah-61安徽菌核型中等34.5962Vdah-62安徽菌核型强39.0063Vdah-63安徽中间型强40.2964Vdah-64安徽菌核型强43.1765Vdah-65安徽菌丝型强46.9066Vdah-66安徽菌核型强47.3467Vdah-67安徽菌核型强59.6368V991b对照菌核型强45.2169AY对照菌丝型中等28.95
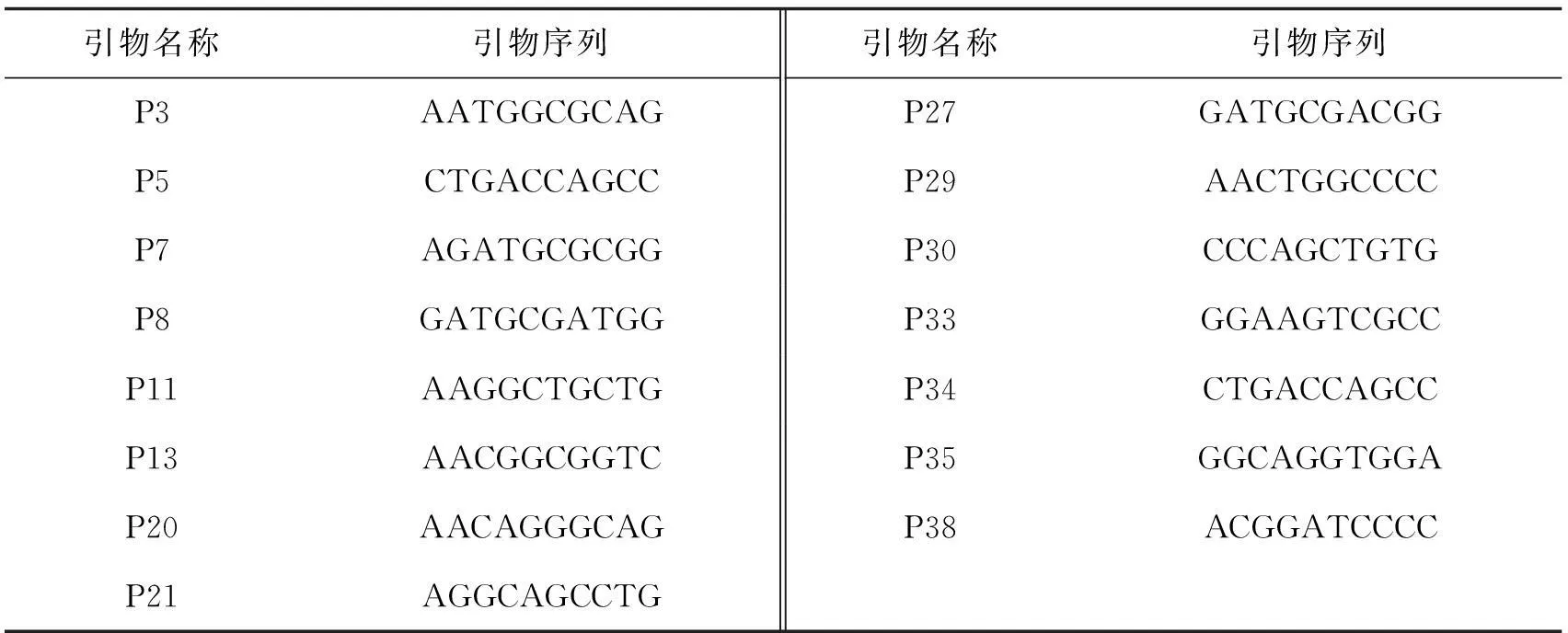
表3 筛选出的随机引物序列
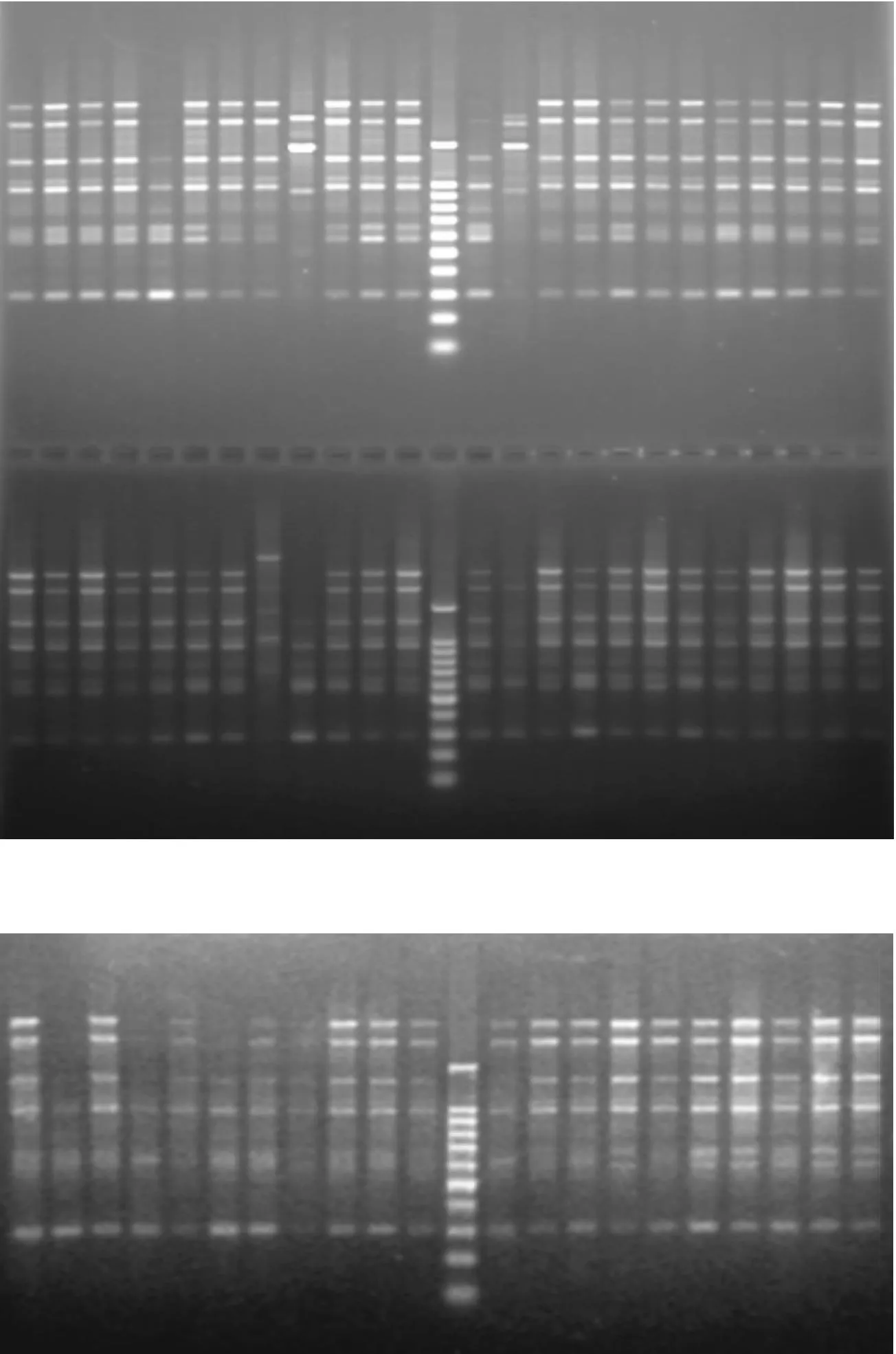
图1 引物P20对菌株DNA扩增产物的电泳图
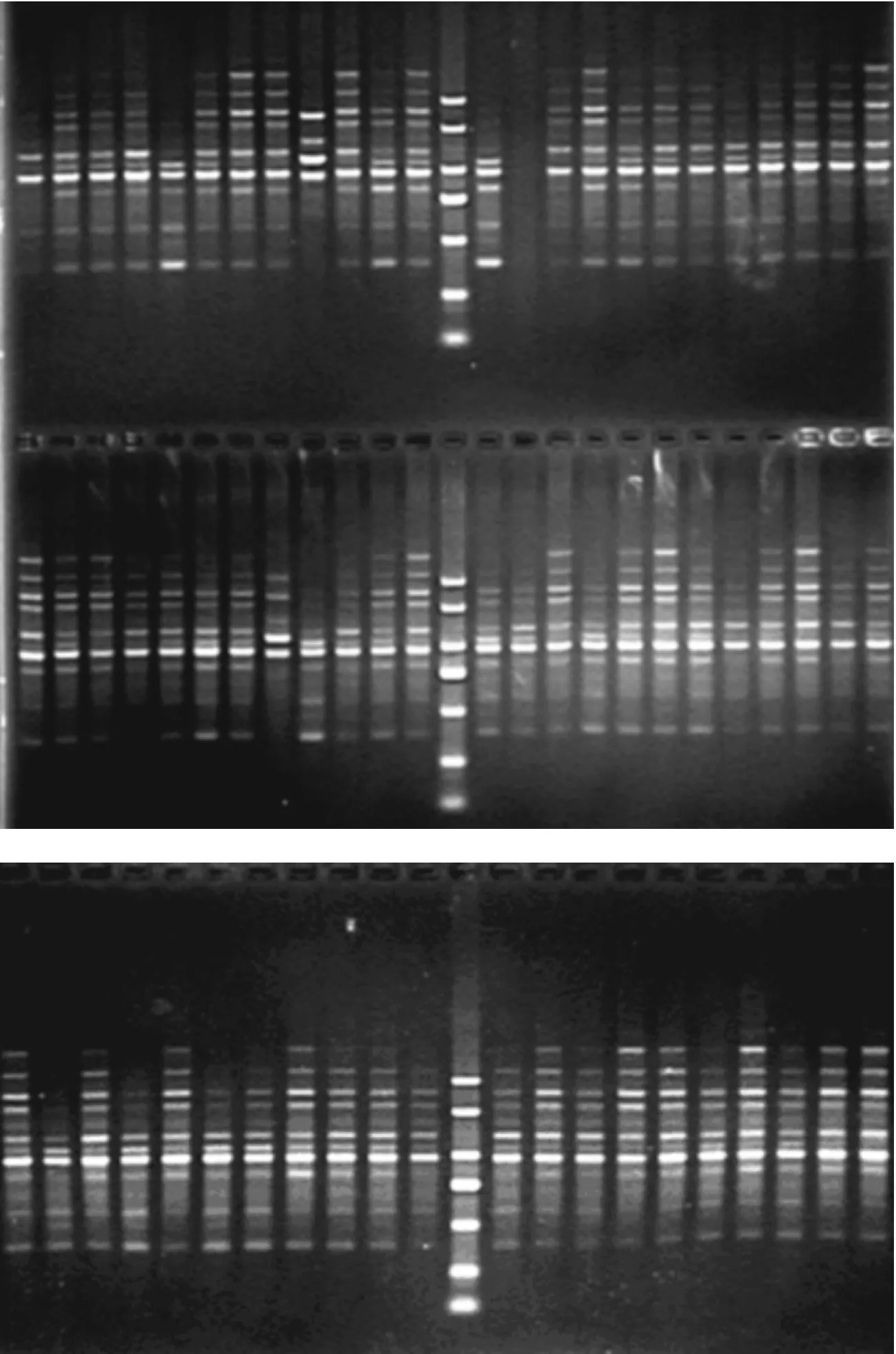
图2 引物P11对菌株DNA扩增产物的电泳图

图3 供试黄萎病菌RAPD聚类分析树状

图4 特异性引物D1/D2扩增的电泳图谱 图4中所有的菌株顺序都是按照表2中的菌株顺序 从左至右,从上至下. 正中间条带为DNA Marker. 图4中的Marker的大小从下至上依次为100、 250、 500、 750、 1 000、 2 000、 3 000
2.4棉花黄萎病菌的RAPD分析从电泳图谱上可以较为直观地看出供试菌株的扩增产物表现出较为丰富的多态性,见图1.不同地区和同一地区的黄萎病菌株均表现出遗传上的多态性.用ntsys软件对数据进行UPGMA法聚类分析,建立树状图(见图3).
从聚类分析树状图(树状图中的数字对应表2中的菌株编号)可以看出, 69个菌株在阈值0.69处被分为2个RAPD Group(简称为RG),3个菌株Vdjx-9(9)、Vdjx-14(14)和Vdhn-32(32)被归到RG1;其余66个菌株被归到RG2, RG2又分为2个亚组,其中Vdjx-5(5)、Vdhn-33(33)、Vdah-50(50)和Vdjx-13 (13)被分到一个亚组,其他62个菌株均集中在另一个亚组.对照菌株V991b(68)和AY(69)也在其中.在RG1中,Vdjx-9为菌丝型,中等致病力菌株;Vdjx-14为菌核型,强致病力菌株;Vdhn-32为菌核型,弱致病力菌株,这3个菌株均属于不同的培养性状类型和致病力类型.而在RG2中被分出来的一个亚组里面的4个菌株Vdjx-5、 Vdhn-33、 Vdah-50和 Vdjx-13也同样属于不同的培养性状类型和致病力类型.
2.5特异性引物的扩增国外报道过一对特异性引物,对于落叶型大丽轮枝菌菌株,该对引物可扩增出一段550 bp的片段,非落叶性大丽轮枝菌则不能被扩增[5].对所有供试菌株进行扩增的结果是所有供试菌株都能扩增出550 bp的DNA片段.其结果如图4.
3讨论
棉花黄萎病的病原菌大丽轮枝菌是一种变异性较强的菌,表现为呈现出不同的菌落的培养形态的差异,致病力的分化以及DNA序列上的差异.从致病力的强弱来看,国外一般分为落叶型强致病力菌系和非落叶型弱致病力菌系[6];国内一般分为强致病力菌系、中等致病力菌系和弱致病力菌系[7].笔者采用本实验室研发出来的一种裸苗浸根接种技术对67个来自长江流域4个不同省份湖北省、湖南省、江西省和安徽省的菌株进行致病力的测定,这种方法将基质育苗和浸根接种相结合,具有简单易操作和周期短等优点.
大丽轮枝菌的菌株不仅在致病力上存在分化,其菌落培养性状的类型也会出现不同程度的分化,根据宋晓轩等的描述,棉花黄萎病菌菌落培养类型有3种——菌核型、中间型和菌丝型[8],菌核型表现产生大量黑色微菌核,中间型产生少量微菌核,菌丝型则表现产生旺盛的白色菌丝.对67个菌株致病力的测定结果表明,强致病力类型、中等致病力类型和弱致病力类型分别占35.82%、43.28%和17.91%.同时,对这67个供试菌株的培养性状类型进行分类表明,属菌核型有26个,占38.81%;属中间型的有15个,占22.39%;属菌丝型的有26个,占38.81%.将致病力鉴定结果和培养性状类型鉴定结果进行比较分析发现, 24个强致病力菌株中有13个为菌核型菌株,以菌核型为主,占54.17%,菌丝型则仅有3个,占12.5%; 14个弱致病力菌株中有12个为菌丝型菌株,以菌丝型为主,占85.71%,菌核型则没有.各种致病力类型和培养类型在各个省份分布较为均匀,说明菌株的致病力和菌落培养类型与地理位置之间并没有相关性,然而,菌株的致病力与菌落培养类型之间存在一定的相关性,强致病菌株大多数为菌核型,而弱致病菌株则以菌丝型为主.菌核型能够产生大量微菌核,菌丝型则不能,而微菌核是一种抵抗不良环境的休眠结构,推断产生大量微菌核的菌株更能够适应有机体内的环境,能够更好的在植物体内定殖和生长.
利用筛选出的15对RAPD引物对所有供试菌株DNA扩增并进行遗传多样性的分析,通过聚类分析我们发现,不同地区和同一地区的菌株均呈现出一定的遗传多样性,证明遗传多样性与菌株来源地无明显相关性.对致病力和菌落培养类型的鉴定结果和遗传多样性分析结果进行比较分析,证明菌株的遗传多样性与菌株致病力和菌落形态也没有明显相关性,这与田新莉等[9-10]的报道有不一致的地方,他们认为RAPD分析与采用鉴别寄主法进行致病性研究间有一定的相关性,但是他们也指出并不是完全吻合.
Encarnacion Perez-Artes等[5]用落叶型菌系的一对特异性引物进行PCR扩增,落叶型菌系能扩增出550 bp的片段,非落叶型则不能扩增出片段.张莉等[11]利用这对特异性引物对新疆不同地区的菌株DNA进行扩增,其中只有3个新疆供试菌株能扩增出550 bp的片段.本文中用这对引物对供试菌株DNA进行扩增,发现所有的菌株都能够扩增出一段550 bp的片段.但是并不是所有的菌株都是落叶型的,推测由于黄萎病菌的变异性强,在逐渐变异的过程中基因会向优势种群靠近.
综上所述,长江流域黄萎病菌的遗传多样性与菌株来源、致病力以及培养性状在一定程度上没有相关性,但是致病力和菌落形态之间有一定的相关性,是否需要收集不同地区的更多材料来证实这一点,有待进一步的研究和探讨.
4参考文献
[1] 李生才,周运宁,郝赤,等.棉田有害生物综合治理[M].北京:中国农业科技出版社,1998:121-129.
[2] 金利容,万鹏,孔令甲,等.湖北省棉花黄萎病病菌致病力分化研究[J].棉花学报,2011,23(6):566-572.
[3] Tayor J W,Natvig D O.Isolation of fungal DNA[Z].Personal Communication,1998:1-16.
[4] 石磊岩,王波,文学.我国棉花黄萎病类型分化及培养特性研究[J].植物保护学报,1993,15(3):247-252.
[5] Encarnacion Perez-Artes, Maria D Garcia-Pedrajas, Jose Bejarano-Alcazar et al. Differentiation of cotton-defoliating and non defoliating pathotypes ofVerticilliumdahliaeby RAPD and specific PCR analyses[J]. European Journal of Plant Pathology, 2000, 106:507-517.
[6] Ramsay J R. RAPD-PCR identification of Verticillium dahliae isolate with differential pathogenicity on cotton[J]. Australian Journal of Agricultural Research, 1996, 47(5):681-693.
[7] 朱荷琴,冯自力,尹志新,等.我国棉花黄萎病菌致病力分化及ISSR指纹分析[J].植物病理学报,2012,42( 3):225-235.
[8] 宋晓轩,朱荷琴,郭金城. 棉花黄萎病(VerticilliumdahliaeKleb. )安阳菌系致病力分化研究[J].棉花学报,1997, 30(1): 13-18.
[9] 田新莉,李晖,赵宗胜,等. 新疆棉花黄萎菌不同致病类型的RAPD指纹分析[J].棉花学报,2001,13(6):346-350.
[10] 刘学堂,郭金城,张元恩,等. 中国主产棉区黄萎病菌的RAPD分析[J].华北农学报,1999,14(1):107-114.
[11] 张莉,段维军,李国英,等. 应用聚合酶链式反应鉴定新疆棉花落叶型黄萎菌[J].植物检疫,2004,18(5):266-268.
(责任编辑游俊)
The pathogenic diversity and the genetic diversity of cottonVerticilliumdahliaein the Yangtze River Basin
JIN Lirong, WAN Peng, HUANG Wei, YANG Xiaohua
(Institute of Plant Protection and Soil Science, Hubei Academy of Agricultural Sciences, Wuhan 430079,China)
Abstract:Cotton Verticillium wilt is an important disease in the world,and a serious threat to cotton production.The four provinces, Hubei Province, Hunan Province, Jiangxi Province and Anhui Province, lie in the Yangtze River Basin. More than three hundreds isolates were collected and 67 isolates were selected from different cities and counties as the tested strains, then the cultural character, the pathogenic diversity and genetic diversity of the 67 strains were analyzed. The results showed that the high virulent types, the medium virulent types and the weak virulent types accounted for 35.82%, 43.28% and 17.91% respectively.The microsclerotia types, the intermediate types and the hyphae types acounted for 38.81%, 22.39% and 38.81% respectively. Further analysis showed that there was no correlation between the pathogenicity, the cultural character and the strain origin. However, there were some correlations between the pathogenicity and the cultural character. Most of high virulent strains belonged to microsclerotia types and the weak virulent strains dominate hyphae types.Fifteen pairs of RAPD primers were gained for DNA amplification, cluster analysis showed that the genetic diversity had no significant correlation with the strain origin. Furthermore, the genetic diversity of the strains had no significant correlation with the pathogenicity and the cultural character.
Key words:the Yangtze River Basin; cotton Verticillium dahliae; culture character; the pathogenic diversity; the genetic diversity
中图分类号:S435.62
文献标志码:A
DOI:10.3969/j.issn.1000-2375.2016.02.007
文章编号:1000-2375(2016)02-0120-07
作者简介:金利容(1982-),女,助理研究员
基金项目:长江中游棉区棉花重大病虫害防控技术研究与集成示范项目(2012BAD19B05-006)资助
收稿日期:2015-07-28
猜你喜欢
疯狂英语·新悦读(2019年12期)2020-01-06
中国环境监察(2018年10期)2018-11-14
中国工程咨询(2017年4期)2017-01-31
长江蔬菜·学术版(2016年12期)2017-01-12
学习月刊(2016年14期)2016-07-11
现代园艺(2016年3期)2016-03-24
天津农业科学(2015年5期)2015-05-30
中国工程咨询(2014年11期)2014-02-16
