
春季和夏季渤海湾近岸海域大型砂壳纤毛虫的群落结构
2016-02-09 02:29:18马玉艳王群山张武昌
海洋科学 2016年11期
于 莹, 马玉艳, 陈 卫, 王群山, 张武昌
(1. 天津渤海水产研究所 渔业资源与生态环境研究室, 天津 300457; 2. 国家海洋局 天津海洋环境监测中心站, 天津 300457; 3. 中国科学院 海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071)
春季和夏季渤海湾近岸海域大型砂壳纤毛虫的群落结构
于 莹1, 马玉艳2, 陈 卫1, 王群山1, 张武昌3
(1. 天津渤海水产研究所 渔业资源与生态环境研究室, 天津 300457; 2. 国家海洋局 天津海洋环境监测中心站, 天津 300457; 3. 中国科学院 海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071)
为了解渤海湾砂壳纤毛虫丰度、生物量及种类组成, 于2011年春季和夏季在渤海湾天津近岸海域进行砂壳纤毛虫群落的采样调查。两个季节砂壳纤毛虫丰度、生物量、优势种及群落结构均有显著的不同。春季砂壳纤毛虫丰度的范围为0~1608个/L, 生物量为0.00~20.03 µg /L; 夏季砂壳纤毛虫丰度的范围为2~313个/L, 生物量为0.03~4.04 µg /L。共鉴定出砂壳纤毛虫5属15种, 其中拟铃虫属(Tintinnopsis)种类最多。春季的主要优势种为运动类铃虫 (Codonellopsis mobilis, 优势度为0.79), 夏季的主要优势种为斯氏拟铃虫 (Tintinnopsis schotti, 优势度为0.28)和诺氏薄铃虫 (Leprotintinnus nordqvisti, 优势度为0.28)。春季砂壳纤毛虫群落的物种丰富度、Shannon指数和Pielou指数均明显低于夏季。
砂壳纤毛虫; 丰度; 生物量; 群落结构; 渤海湾
砂壳纤毛虫普遍存在于海湾、河口、陆架及大洋中, 是浮游纤毛虫和微型浮游动物的重要组成类群;其主要摄食浮游植物、鞭毛虫等, 在海洋浮游生态系统的物质循环和能量流动中发挥重要的作用[1-2]。迄今,世界海区记录的砂壳纤毛虫近900种, 中国海区共记录140余种[3]。与我国其他海区相比, 渤海砂壳纤毛虫生态学的研究起步较晚, 资料较少, 主要集中在渤海中部[4-5]和莱州湾[6-7]。目前对渤海砂壳纤毛虫生态学的研究出现断层, 近10年来很少有渤海砂壳纤毛虫生态学的相关报道, 渤海砂壳纤毛虫生态学的研究急需开展。
渤海湾属于渤海三大海湾之一, 位于渤海西部,是一个典型的半封闭性海湾, 是许多经济鱼虾类的产卵场和索饵场, 对黄渤海的渔业经济意义重大。关于渤海湾浮游植物和浮游动物的群落结构的研究很多[8-9]。但是渤海湾浮游纤毛虫的资料尚少。本文于2011年5月和8月在渤海湾天津近岸海域进行砂壳纤毛虫丰度和生物量的调查, 为渤海湾浮游生态系统研究及渔业资源的发展提供基础数据和参考资料。
1 材料与方法
于2011年春季 (5月)和夏季 (8月)在渤海湾天津近岸海域5个断面共20个站位进行砂壳纤毛虫样品的采集 (图1)。在各站位采用浮游生物Ⅲ型网 (网口面积0.1 m2, 网孔直径76 μm)由底至表垂直拖网取样, 样品用5%福尔马林液固定后带回实验室。在Olympus SZX16体视显微镜下观察计数。测量虫体的体长、体宽, 按最接近的几何形状 (如柱体、球体和锥体)计算体积, 生物量由体积乘转换系数 (0.19 pg/µm3, 以碳计)[10]得来。砂壳纤毛虫根据文献[3, 11-12]鉴定到种。
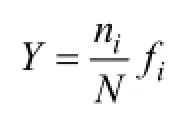
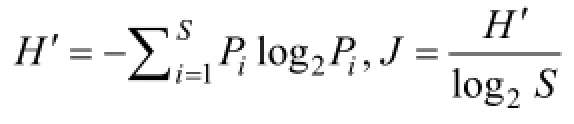
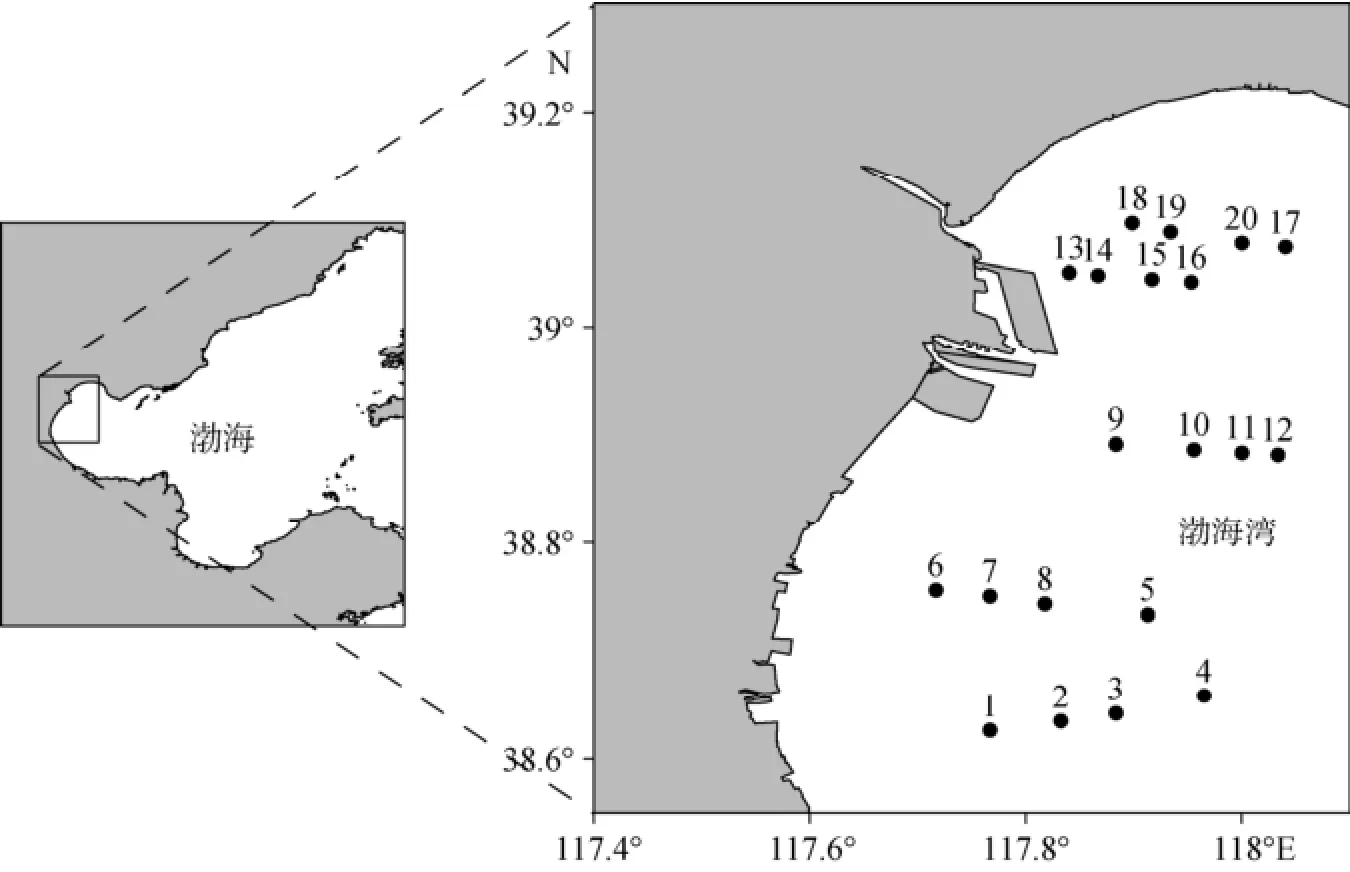
图1 调查站位分布图Fig. 1 Location of sampling stations in Bohai Bay
2 结果
2.1 砂壳纤毛虫丰度和生物量
春季砂壳纤毛虫的平均丰度为(371±497)个/L,范围为0~1608个/L, 最大值出现在6号站, 主要是运动类铃虫, 其丰度占砂壳纤毛虫丰度的89.6%;砂壳纤毛虫的平均生物量为(3.60±5.56)µg/L, 范围为0.00~20.03 µg/L, 最大值出现在6号站, 主要是日本拟铃虫和运动类铃虫, 其生物量分别占砂壳纤毛虫生物量的52.8%和46.3%。砂壳纤毛虫丰度和生物量的高值区均主要分布在西南部近岸海区 (离岸距离<4~5 km), 北部及远岸海区值较低 (图2)。
夏季砂壳纤毛虫的平均丰度为(83±90)个/L, 范围为2~313个/L, 最大值出现在14号站, 主要是诺氏薄铃虫和斯氏拟铃虫, 其丰度分别占砂壳纤毛虫丰度的39.3%和32.8%; 砂壳纤毛虫的平均生物量为(0.95±1.10)µg/L, 范围为0.03~4.04 µg /L, 最大值出现在6号站, 主要是斯氏拟铃虫, 其生物量占砂壳纤毛虫生物量的84.1%。砂壳纤毛虫丰度和生物量的高值区均主要分布在西北及西南部近岸 (离岸距离<4~5 km), 远岸海区值较低 (图3)。
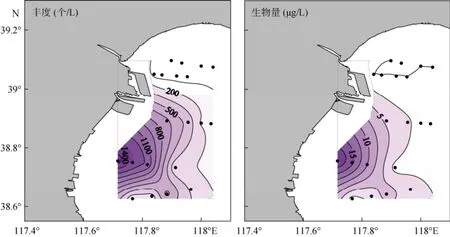
图2 春季砂壳纤毛虫丰度和生物量的水平分布Fig. 2 Horizontal distribution of tintinnid abundance and biomass in spring
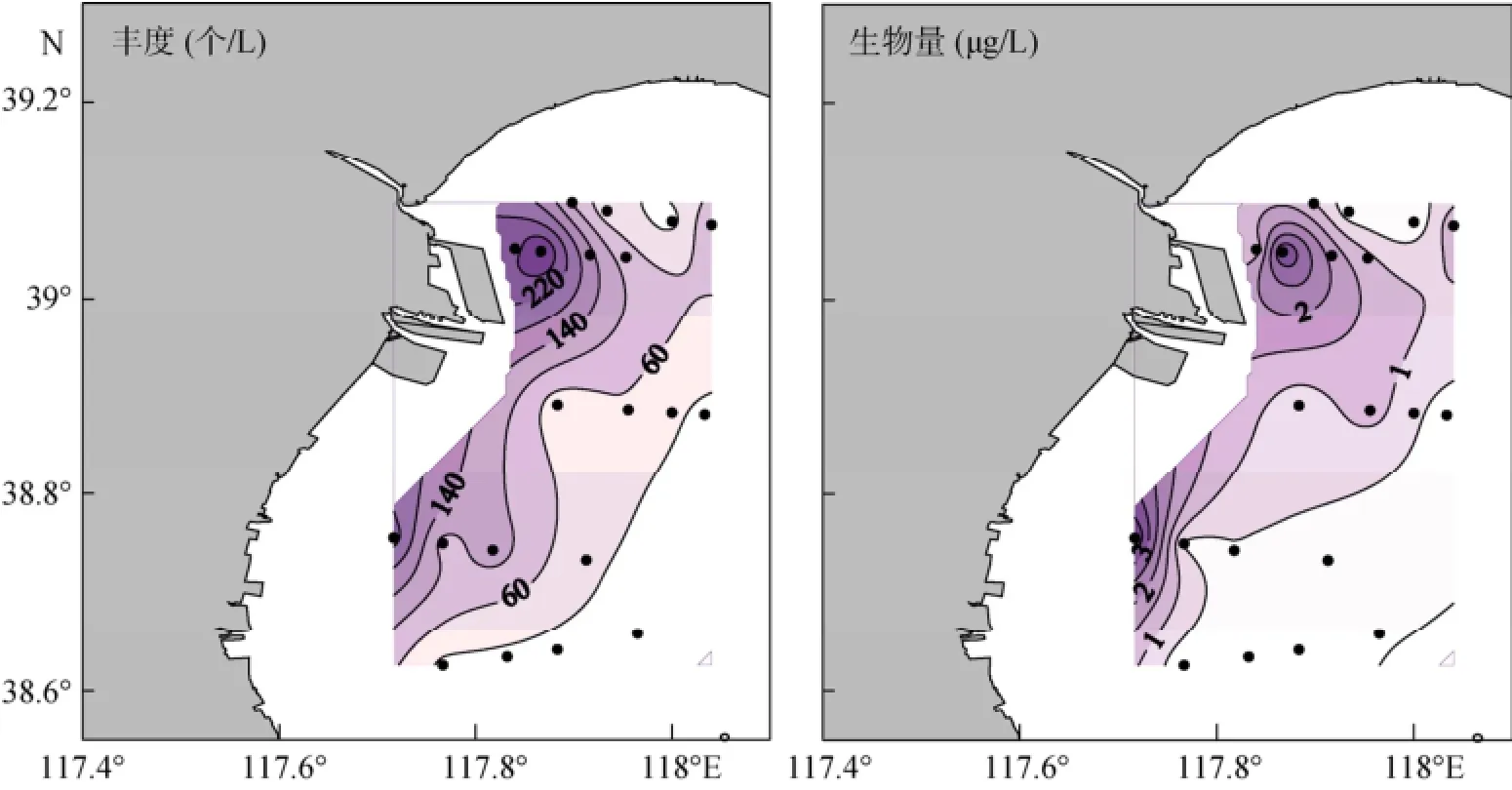
图3 夏季砂壳纤毛虫丰度和生物量的水平分布Fig. 3 Horizontal distribution of tintinnid abundance and biomass in summer
2.2 种类组成
春季和夏季共鉴定出砂壳纤毛虫5属15种, 其中拟铃虫属 (Tintinnopsis)种类最多, 为9种 (表1)。爱氏网纹虫 (Favella ehrenbergii)、原始筒壳虫(Tintinnidium primitivum)、厦门拟铃虫 (Tintinnopsis amoyensis)和最小拟铃虫 (Tintinnopsis minima)是渤海的新纪录种。
春季运动类铃虫 (Codonellopsis mobilis)占绝对优势 (优势度为0.79), 其平均丰度为(346±457)个/L,范围为0~1440 个/L, 其丰度占砂壳纤毛虫丰度的平均值为(95.5±5.8)%; 运动类铃虫的平均生物量为(2.23±2.95)µg /L, 范围为0.00~9.28 µg /L, 其生物量占砂壳纤毛虫生物量的平均值为(83.7±20.4)%。运动类铃虫丰度和生物量的最大值均出现在6号站, 其高值区主要分布在西南部近岸, 北部及远岸海区值较低 (图4)。
夏季的主要优势种为斯氏拟铃虫 (Tintinnopsis schotti, 优势度为0.28)和诺氏薄铃虫 (Leprotintinnus nordqvisti, 优势度为0.28)。

表1 春季和夏季鉴定的砂壳纤毛虫的种类Tab. 1 List of tintinnid species in Bohai Bay during spring and summer
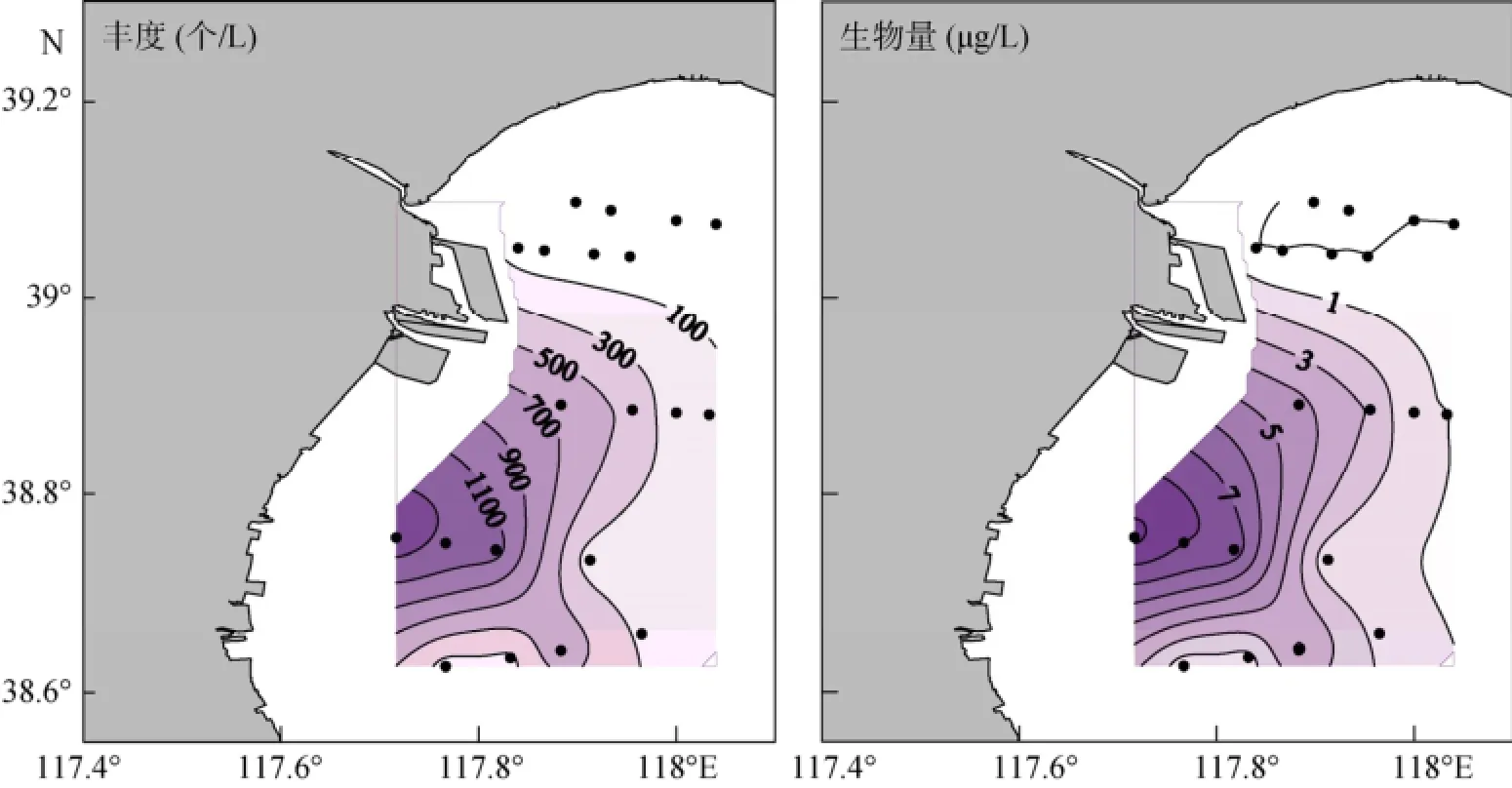
图4 春季运动类铃虫丰度和生物量的水平分布Fig. 4 Horizontal distribution of Codonellopsis mobilis abundance and biomass in spring
斯氏拟铃虫的平均丰度为(23±36)个/L, 范围为1~136个/L; 斯氏拟铃虫的平均生物量为(0.59± 0.89)µg /L, 范围为0.02~3.40 µg /L。斯氏拟铃虫丰度和生物量的最大值均出现在6号站, 其高值区主要分布在西北及西南部近岸, 远岸海区值较低 (图5)。
诺氏薄铃虫的平均丰度为(27±45)个/L, 范围为0~170 个/L; 诺氏薄铃虫的平均生物量为(0.08± 0.12)µg/L, 范围为0.00~0.47 µg/L。诺氏薄铃虫丰度和生物量的最大值均出现在13号站, 其高值区主要分布在西北部近岸, 南部及远岸海区值较低 (图6)。
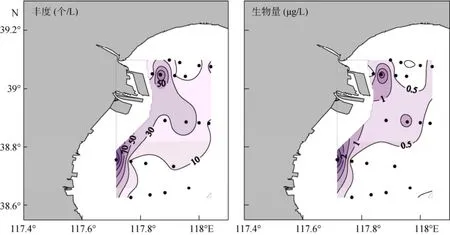
图5 夏季斯氏拟铃虫丰度和生物量的水平分布Fig. 5 Horizontal distribution of Tintinnopsis schotti abundance and biomass in summer
2.3 群落特征
春季砂壳纤毛虫群落Shannon指数的平均值为0.24±0.27, 最大值为0.90, 出现在4号站; Pielou指数的平均值为0.11±0.15, 最大值为0.57, 出现在4号站。Shannon指数和Pielou指数的水平分布模式类似,高值区主要出现在南部海区, 北部值较低 (图7)。
夏季砂壳纤毛虫群落Shannon指数的平均值为1.76±0.44, 最大值为2.46, 出现在8号站; Pielou指数的平均值为0.64±0.16, 最大值为0.95, 出现在4号站。Shannon指数和Pielou指数的水平分布模式类似, 高值区主要出现在北部和南部海区, 中部值较低 (图8)。
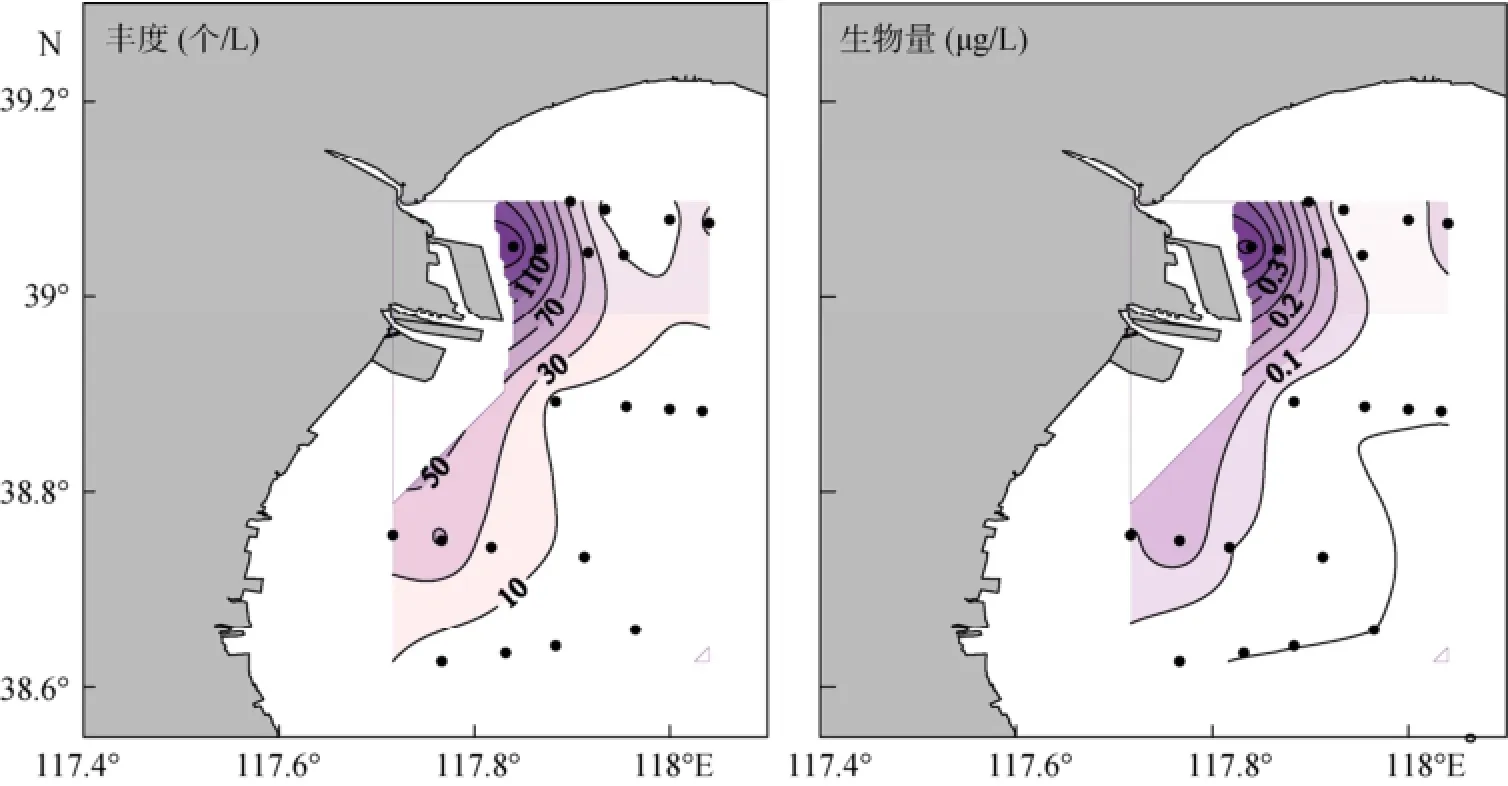
图6 夏季诺氏薄铃虫丰度和生物量的水平分布Fig. 6 Horizontal distribution of Leprotintinnus nordqvisti abundance and biomass in summer
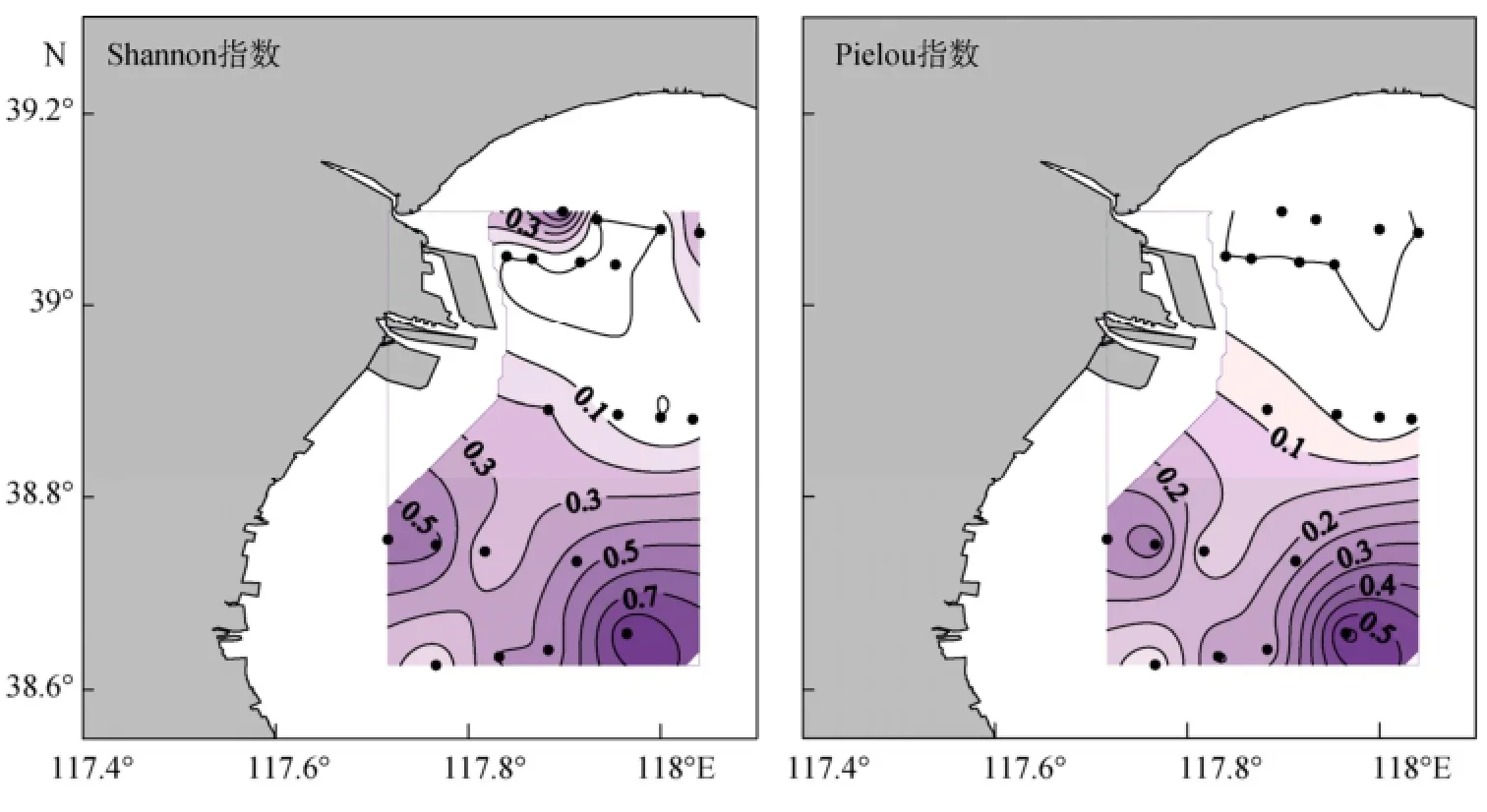
图7 春季砂壳纤毛虫群落Shannon指数和Pielou指数的水平分布Fig. 7 Horizontal distribution of tintinnid Shannon index and Pielou index in spring
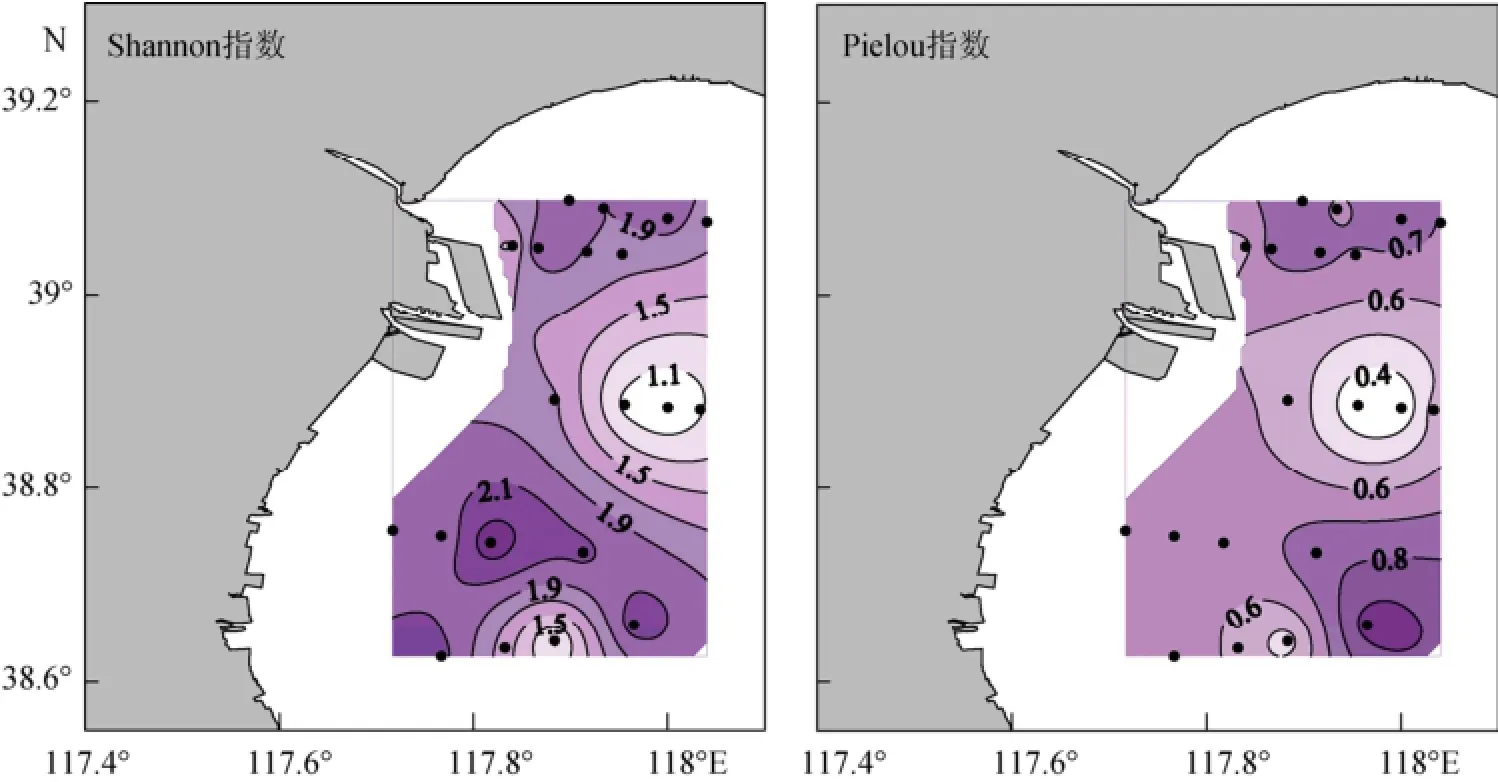
图8 夏季砂壳纤毛虫群落Shannon指数和Pielou指数的水平分布Fig. 8 Horizontal distribution of tintinnid Shannon index and Pielou index in summer
3 讨论
3.1 丰度和生物量
本研究中春季和夏季砂壳纤毛虫丰度和生物量均表现出近岸较高、远岸较低的趋势。这与大多数温带近岸海区的分布规律一致。如Ise Bay[16]及Hiroshima Bay[17], 砂壳纤毛虫的丰度和生物量在湾内比湾外高。这可能与其饵料的主要组成—浮游植物的分布有关[16,18]。渤海湾天津近岸海域浮游植物丰度的高值区基本出现在近岸[19-20], 从而为砂壳纤毛虫提供较高水平的饵料。
3.2 运动类铃虫
春季渤海湾砂壳纤毛虫几乎全是运动类铃虫,其丰度占砂壳纤毛虫丰度的95%以上, 是春季的绝对优势种, 这种现象在渤海以前的研究中[21-22]已有报道。可见, 运动类铃虫是春季渤海优势度较大的砂壳纤毛虫。
此外, 运动类铃虫在黄海[23-24]、东海[25]和南海[26-27]都有分布。各海区运动类铃虫一般在5~8月丰度较高; 且运动类铃虫最大丰度从北到南逐渐降低。渤海湾运动类铃虫的最大丰度要远远高于其他海区(表2)。

表2 不同海区运动类铃虫的最大丰度Tab. 2 Maximum abundance of Codonellopsis mobilis in China seas
3.3 群落结构
本研究中春季鉴定的砂壳纤毛虫的种类数远远小于夏季, 这与胶州湾[23]、桑沟湾[24]、莱州湾[6]等海湾的研究结果类似, 砂壳纤毛虫的种类数一般在温度较高的夏季较高。此外, 与莱州湾[6]相似, 渤海湾春季砂壳纤毛虫群落的Shannon指数和Pielou指数均明显低于夏季, 这可能表明春季砂壳纤毛虫群落稳定性较差, 夏季砂壳纤毛虫群落则相对稳定。
[1] Azam F, Fenchel T, Field J G, et al. The ecological role of water column microbes in the sea[J]. Marine Ecology Progress Series, 1983, 10: 257-263.
[2] Pierce R W, Turner J T. Plankton studies in Buzzards Bay, Massachusetts, USA, IV. Tintinnids, 1987-1988[J]. Marine Ecology Progress Series, 1994, 112: 235-240.
[3] 张武昌, 丰美萍, 于莹, 等. 砂壳纤毛虫图谱[M]. 北京: 科学出版社, 2012: 1-499. Zhang Wuchang, Feng Meiping, Yu Ying, et al. Tintinnids map[M]. Beijing: Science Press, 2012: 1-499.
[4] Zhang W, Wang R. Short time dynamics of ciliate abundance in the Bohai Sea (China)[J]. Chinese Journal of Oceanology and Limnology, 2002, 20(2): 135-141.
[5] Zhang W, Wang R. Rapid changes in stocks of ciliate microzooplankton associated with a hurricane in the Bohai Sea (China)[J]. Aquatic Microbial Ecology, 2000, 23(1): 97-101.
[6] 陈雪, 张武昌, 吴强, 等. 莱州湾大型砂壳纤毛虫群落季节变化[J]. 生物多样性, 2014, 22(5): 649-657. Chen Xue, Zhang Wuchang, Wu Qiang, et al. Laizhou Bay change large tintinnids seasonal[J]. Biodiversity, 2014, 22(5): 649-657.
[7] 张武昌, 王荣, 王克. 1997年7月一航次中莱州湾自由生纤毛虫和桡足类幼虫的分布[J]. 海洋科学, 2002, 26(9): 20-21. Zhang Wuchang, Wang Rong, Wang Ke. A cruise in July 1997 in the Gulf of Laizhou free of planktonic ciliates and copepod larvae distribution of[J]. Marine Sciences, 2002, 26(9): 20-21.
[8] 周然, 彭士涛, 覃雪波, 等. 渤海湾浮游植物与环境因子关系的多元分析[J]. 环境科学, 2013, 34(3): 864-873. Zhou Ran, Peng Shitao, Qin Xuebo, et al. Multivariate analysis phytoplankton and environmental factors ofBohai Bay[J]. Environmental Science, 2013, 34(3): 864-873.
[9] 高文胜, 刘宪斌, 张秋丰, 等. 渤海湾近岸海域浮游动物多样性[J]. 海洋科学, 2014, 38(4): 55-60. Gao Wensheng, Liu Xianbin, Zhang Qiufeng, et al. The diversity of zooplankton in the coastal waters of Bohai Bay[J]. Marine Science, 2014, 38(4): 55-60.
[10] Putt M, Stoecker D K. An experimentally determined carbon: volume ratio for marine “oligotrichous” ciliates from estuarine and coastal waters[J]. Limnology and Oceanography, 1989, 34(6): 1097-1103.
[11] 尹光德. 胶州湾砂壳纤毛虫之初步调查[J]. 山东大学学报, 1952, 2(1): 36-56. Yin Guangde. A preliminary investigation on Jiaozhou Bay tintinnids[J]. Journal of Shandong University, 1952, 2(1): 36-56.
[12] Hada Y. The fauna of Akkeshi Bay IV. The pelagic ciliata[J]. Journal of the Faculty of Science, Hokkaido Imperial University, Series VI, Zoology, 1937, 4(5): 143-216.
[13] 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 1989, 8(4): 13-15. Xu Zhaoli, Chen Yaqu. East the Yellow Sea sub dominant species in autumn animal gathered strength and mackerel SCAD fishery related[J]. Journal of Ecology, 1989, 8(4): 13-15.
[14] Shannon C E, Weaver W. The Mathematical Theory of Communication[M]. Urbana: University of Illinois Press. 1949: 1-114.
[15] Pielou E C. The measurement of diversity in different types of biological collections[J]. Journal of Theoretical Biology, 1966, 13: 131-144.
[16] Uye S I, Nagano N, Shimazu T. Abundance, biomass, production and trophic roles of micro- and net-zooplankton in Ise Bay, central Japan, in winter[J]. Journal of Oceanography, 2000, 56(4): 389-398.
[17] Kamiyama T. The impact of grazing by microzooplankton in northern Hiroshima Bay, the Seto Inland Seam, Japan[J]. Marine Biology, 1994, 119(1): 77-88.
[18] Buck K R, Garrison D L, Hopkins T L. Abundance and distribution of tintinnid ciliates in an ice edge zone during the austral autumn[J]. Antarctic Science, 1992, 4(1): 3-8.
[19] 尹翠玲, 张秋丰, 崔健, 等. 2008-2012年渤海湾天津近岸海域夏季浮游植物组成[J]. 海洋科学进展, 2013(4): 527-537. Yin Cuiling, Zhang Qiufeng, Cui Jian, et al. Phytoplankton composition in Bohai Bay Tianjin coastal area in summer from 2008 to 2012[J]. Advances in Marine Science, 2013, 31(4): 527-537.
[20] 马玉艳, 张秋丰, 徐玉山, 等. 渤海湾基础生物资源现状及其变化趋势[J]. 海洋环境科学, 2013, 32(6): 845-850. Ma Yuyan, Zhang Qiufeng, Xu Yushan, et al. The present situation and trend of the basic biological resources in the Bohai Bay[J]. Marine Environmental Science, 2013, 32 (6): 845-850.
[21] 张武昌, 孙军, 孙松. 渤海1999年4月运动类铃虫的平面分布[J]. 海洋科学, 2004, 28(12): 67-69. Zhang Wuchang, Sun Jun, Sun Song. April 1999 Bohai sports bollworm distribution[J]. Marine Sciences, 2004, 28 (12): 67-69.
[22] 张武昌, 王荣. 渤海微型浮游动物及其对浮游植物的摄食压力[J]. 海洋与湖沼, 2000, 31(3): 252-258. Zhang Wuchang, Wang Rong. Bohai animal and microzooplankton grazing pressure on the phytoplankton[J]. Oceanologia et Limnologia Sinica, 2000, 31(3): 252-258.
[23] 于莹, 张武昌, 赵楠等. 胶州湾浮游纤毛虫丰度和生物量的周年变化[J]. 海洋与湖沼, 2011, 42(5): 690-701. Yu Ying, Zhang Wuchang, Zhao Nan et al. Jiaozhou Bay planktonic ciliate abundance and biomass of the annual variation of[J]. Oceanologia et Limnologia Sinica, 2011, 42 (5): 690-701.
[24] 于莹, 张武昌, 蒋增杰等. 桑沟湾浮游纤毛虫丰度和生物量分布的季节变化[J]. 海洋学报, 2013, 35(3): 215-224. Yu Ying, Zhang Wuchang, Jiang Zengjie. The Sanggou Bay planktonic ciliate abundance and biomass distribution of the seasonal variation of[J]. Acta oceanologica Sinica, 2013, 35 (3): 215-224.
[25] Yu Y, Zhang W, Feng M, et al. Differences in the vertical distribution and response to freshwater discharge between aloricate ciliates and tintinnids in the East China Sea[J]. Journal of Marine Systems, http: // dx.doi.org/10.1016/j.jmarsys.2015.02.005.
[26] 张翠霞, 张武昌, 肖天. 2007年10月南海北部浮游纤毛虫的丰度和生物量[J]. 生态学报, 2010, 30(4): 0867-0877. Zhang Cuixia, Zhang Wuchang, Xiao Tian. The abundance and biomass of planktonic ciliates in October 2007 in the northern South China Sea[J]. Journal, Ecological, 2010, 30 (4): 867-877.
[27] 于莹, 张武昌, 蔡昱明等. 冬季和夏季南海北部浮游纤毛虫的分布特点[J]. 海洋与湖沼, 2014, 45(4): 839-847. Yu Ying, Zhang Wuchang, Cai Yuming. The winter and summer distribution characteristics of planktonic ciliates in the northern South China Sea[J]. Oceanologia et Limnologia Sinica, 2014, 45 (4): 839-847.
[28] Yu Y, Zhang W, Wang S, et al. Abundance and biomass of planktonic ciliates in the sea area around Zhangzi Island, Northern Yellow Sea[J]. Acta Ecologica Sinica, 2013, 33: 45-51.
[29] Zhang C, Zhang W, Xiao T, et al. Meso-scale spatial distribution of large tintinnids in early summer in southern Yellow Sea[J]. Chinese Journal of Oceanology and Limnology, 2008, 26(1): 81-90.
Received: Jan. 9, 2016
Community structure of large tintinnids (Ciliophora, Tintinnida) in Bohai Bay during spring and summer
YU Ying1, MA Yu-yan2, CHEN Wei1, WANG Qun-shan1, ZHANG Wu-chang3
(1. Laboratory of Fisheries Resources and Ecological Environment, Tianjin Bohai Sea Fisheries Research Institute, Tianjin 300457, China; 2. Tianjin Marine Environmental Monitoring Center Station, SOA, Tianjin 300457, China; 3. Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China)
tintinnid; abundance; biomass; community structure; Bohai Bay
The community structure and seasonal dynamics of tintinnids in Bohai Bay were investigated during spring and summer, 2011. Tintinnid abundance, biomass, and community structure were different in spring and summer. In spring, tintinnid abundance ranged from 0 to 1608 ind./L, and the biomass was in the range of 0.00-20.03 µg/L; in summer, tintinnid abundance ranged from 2 to 313 ind./L, and the biomass was in the range of 0.03-4.04 µg/L. Tintinnid abundance and biomass were mainly concentrated in the coastal areas during spring and summer. Fifteen tintinnid species were identified, nine of which were in the genus Tintinnopsis. The most dominant species were Codonellopsis mobilis in spring and Tintinnopsis schotti and Leprotintinnus nordqvisti in summer. Species richness, the Shannon index, and the Pielou index were all higher in summer than in spring.
Q958.8
A
1000-3096(2016)11-0057-08
10.11759/hykx20150422002
(本文编辑: 梁德海)
2015-04-22;
2015-09-30
中国科学院海洋生态与环境科学重点实验室开放课题(KLMEES201307); 天津市水产局青年科技项目(J2014-04)
[Foundation: Key Laboratory of marine ecology and environmental science, Chinese Academy of Sciences, Key Laboratory of open project, No.KLMEES201307, Tianjin Municipal Bureau of Fisheries Science and technology project, No.J2014-04]
于莹(1986-), 女, 山东荣成人, 工程师, 博士, 主要从事海洋浮游动物生态学, 电话: 022-59886527, E-mail: yuyingxlf001@ 163.com; 张武昌, 通信作者, 研究员, 博士, E-mail: wuchangzhang@ 163.com
猜你喜欢
工会博览(2022年17期)2022-07-15 07:25:22
海洋通报(2022年2期)2022-06-30 06:06:44
生态学报(2022年9期)2022-05-26 10:03:04
矿产勘查(2020年5期)2020-12-19 18:25:11
科技进步与对策(2020年12期)2020-07-03 07:04:56
华中师范大学学报(人文社会科学版)(2020年3期)2020-06-17 09:02:16
钻井液与完井液(2019年4期)2019-10-10 01:57:10
岷峨诗稿(2019年4期)2019-04-20 09:02:10
渔业科学进展(2018年1期)2018-04-03 08:45:38
童话世界(2017年34期)2017-12-04 09:09:48
