
威海小石岛人工鱼礁海域渔获物群落结构特征及其与环境因子相关性
2016-02-09 02:29王新萌唐衍力孙晓梅盛化香
海洋科学 2016年11期
王新萌, 唐衍力, 孙晓梅, 盛化香, 万 荣
(中国海洋大学 水产学院, 山东 青岛 266003)
威海小石岛人工鱼礁海域渔获物群落结构特征及其与环境因子相关性
王新萌, 唐衍力, 孙晓梅, 盛化香, 万 荣
(中国海洋大学 水产学院, 山东 青岛 266003)
为探究人工鱼礁投放对威海小石岛附近海域渔获物种类组成、渔获量变化、渔获物结构特征的影响, 作者于2012~2013年对威海小石岛人工鱼礁区及对照区的渔获物和环境因子进行了调查, 并采用聚类分析、单因子相似性分析及相似性百分比分析对渔获物结构特征进行分析, 采用典范对应分析(CCA)对礁区主要物种与环境因子的相关性进行分析。结果显示, 南、北礁区地笼网调查到渔获物种类数量高于对照区, 分别为24、31和18种。南、北礁区鱼类和甲壳类生物资源量高于对照区。南、北礁区全年资源量占优势的种类为日本蟳(Charybdis japonica)和大泷六线鱼(Hexagrammos otakii), 对照区为海刺猬(Glyptocidaris crenularis)和大泷六线鱼。聚类分析和单因子相似性分析表明, 渔获物组成结构在季节间差异显著(P<0.05), 在春季和夏季, 人工鱼礁区渔获物组成结构和对照区差异显著(P<0.05)。典范对应分析表明, 水温、叶绿素a和溶解氧是影响礁区主要渔获物种类分布的主要环境因子。投放人工鱼礁改善了威海小石岛礁区水域的渔获物群落结构, 并增加主要鱼类、甲壳类经济物种的渔获量。
人工鱼礁; 渔业资源; 群落结构; CCA; 环境因子
过度捕捞和环境污染导致了中国近海渔业资源的严重衰退[1-2], 而建设人工鱼礁是改善鱼类栖息地、修复渔业生态环境的有效措施之一[3-6]。人工鱼礁投放后产生的上升流能够将海底有机物及营养盐带入海水表层, 促进海洋浮游植物、藻类繁殖, 提高海洋初级生产力。研究发现, 人工鱼礁区的无机氮、磷酸盐、硅酸盐、有机碳、有机氮和有机磷含量均高于非礁区[7], 也有研究表明, 人工鱼礁投放后浮游植物生物量、叶绿素a及初级生产力均高于对照区和投礁前[8]。随着中国沿海人工鱼礁建设逐渐推广,近年来国内许多学者对人工鱼礁区进行了游泳动物资源调查与研究。对嵊泗、海州湾、三横山、俚岛、莱州湾和象山湾等海域的人工鱼礁渔业资源调查发现, 在海区投放人工鱼礁可以明显增加游泳动物资源种类和资源量[9-13], 并能吸引鱼类产卵群体[14]。国外研究发现, 人工鱼礁主要对岩礁性鱼类起到诱集作用[15]。通过对人工鱼礁区某些岩礁性鱼类运动特征的研究发现, 人工鱼礁能够为岩礁性鱼类提供遮蔽和摄食场所, 且鱼类在人工鱼礁的行为与其生理需求及捕食行为相关[16-17]。
国内对人工鱼礁浮游生物群落组成与环境因子相关性的研究较为丰富, 但对游泳动物与环境因子关系的研究相对较少。汪振华等[18]、吴忠鑫等[10]研究发现, 水温、深度能够影响人工鱼礁游泳动物群落变化。国外对此方面研究较多, 降水、河流、水的清澈度、盐度、人工鱼礁的结构复杂性以及礁体的空间分布都会对人工鱼礁区渔业资源产生影响[19-20]。因此, 研究人工鱼礁游泳动物结构与环境要素相关性, 对人工鱼礁的合理建设具有重要意义。
威海小石岛人工鱼礁区位于山东半岛的烟威渔场, 作者通过比较小石岛南、北礁区及其自然对照海区各季节地笼渔获物组成结构的差异, 探索环境因子及浮游动植物细胞密度对人工鱼礁区主要渔获物组成结构的影响, 对小石岛海域人工鱼礁的渔业资源修复效果进行评价。
1 材料与方法
1.1 调查时间与站位
2012年9月和12月、2013年5月和7月, 分别对威海小石岛人工鱼礁南礁区3个站位(S1、S2和S3)、北礁区3个站位(N1、N2和N3)和对照区3个站位(C1、C2和C3)渔业资源和海洋环境因子进行调查(图1)。对照区选择在不受人工鱼礁的流场影响的区域, 章守宇[21]认为人工鱼礁流场影响在水平尺度上不超过单体的50倍, 对照区站位离南礁区边缘最近为580 m, 离北礁区边缘最近距离为780 m, 均超过了鱼礁单体的50倍。
1.2 样品采集与保存
渔业资源调查网具为地笼网, 规格为每节长宽高分别为30、20和25 cm, 24节为一串, 每个站位设置5串, 每次调查的设置时间为24 h。渔获物被带回实验室进行种类鉴定、体长和体质量等生物学参数测定。调查的海洋环境因子为: 水温、盐度、pH、DO、COD、BOD5和叶绿素a。浮游动物、植物样品分别用浅水Ⅰ、Ⅲ型浮游生物网, 自底至表垂直拖网取得。样品的现场采集、保存、测定和分析等过程参照《海洋监测规范》(GB17378-2007)[22]和《海洋调查规范》(GB12763-2007)[23]进行。
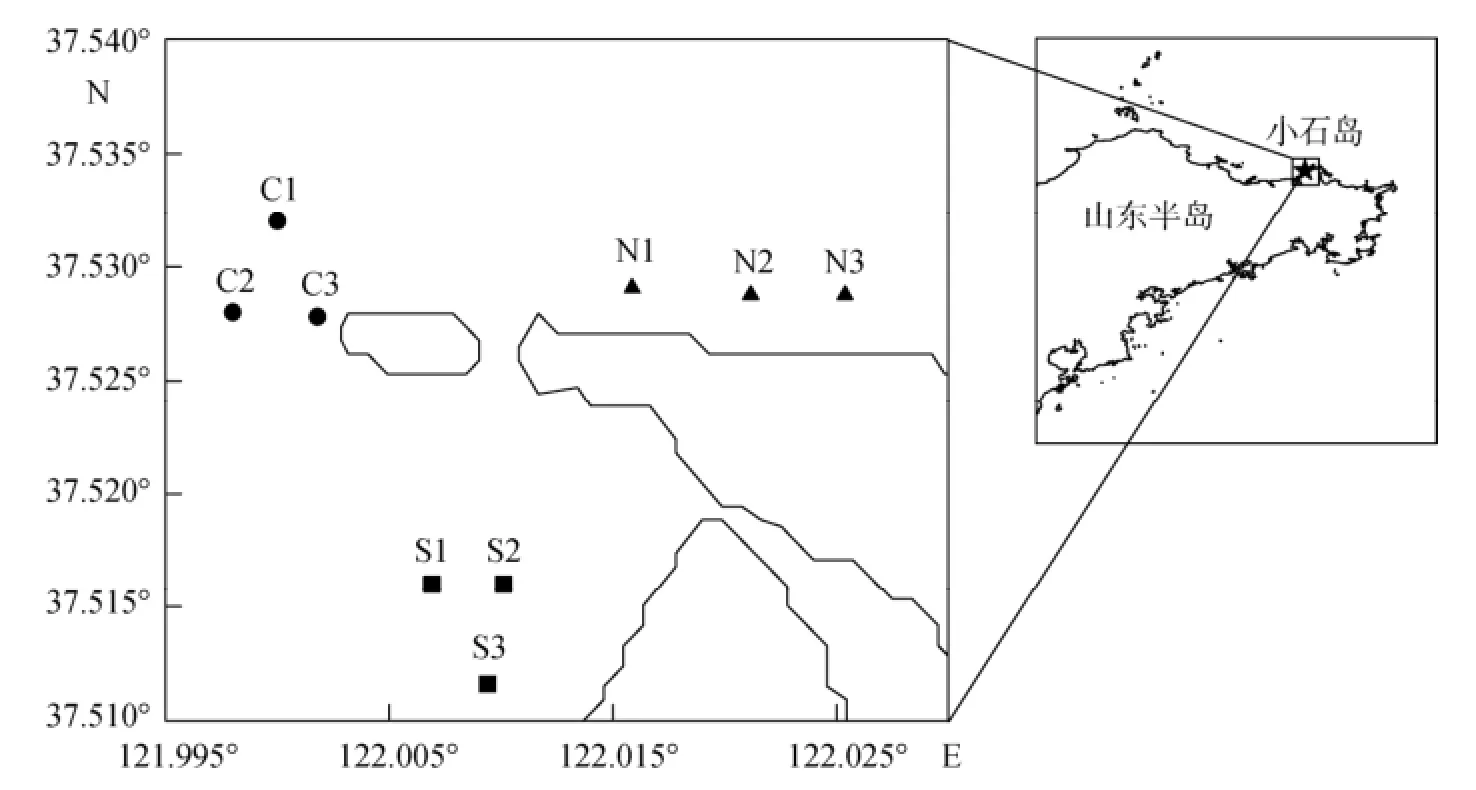
图1 小石岛南、北礁区及对照区调查站位图Fig. 1 Sampling stations in southern and northern reef areas and the control area of Xiaoshi Island
1.3 数据处理
将每站位设置的5串地笼网作为1网笼单元, 地笼网单位捕捞努力量渔获量(CPUE)定义为每天每网笼单元的渔获量, 单位为g/(d·网)[10]。小石岛离岸较近, 南北礁区分别位于小石岛两侧, 南礁区与北礁区海况不同, 因此南、北礁区CPUE分别计算。
采用Pinkas[24]相对重要性指数IRI确定渔获物优势种, IRI>1 000的种类为优势种, 公式如下:
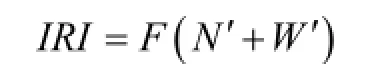
式中, N'为某一种类的尾数占总尾数的百分比; W'为某一种类的质量占总质量的百分比; F为某一种类出现的站数占调查总站数的百分比; 规定IRI >1 000的种类为优势种。
根据各物种的CPUE(g/(d·网))平方根转换计算的Bray Curtis相似性矩阵, 采用聚类分析研究南、北礁区和对照区群落结构。应用单因子相似性分析和相似性百分比分析计算组群之间差异的显著性并鉴别造成群落结构差异的物种。以上分析使用primer5.0软件完成[25-26]。
应用典范对应分析分析水温、盐度、pH、DO、叶绿素a、浮游动植物细胞密度与礁区主要渔获物的关系, 并做出主要渔获物种类与环境因子关系的排序图。以上分析由CANOCO4.5 软件完成[27-29]。
2 结果
2.1 渔获物种类组成
4次调查共获得渔获种类40种, 南礁区、北礁区和对照区的渔获物分别为24种、31种和18种。表1和表2列出了南、北礁区和对照区调查发现的鱼类和甲壳类。南、北礁区共调查到鱼类20种、甲壳类10种、棘皮动物3种、软体动物3种。南礁区调查到硬骨鱼类13种、甲壳类8种, 北礁区调查到硬骨鱼类19种、甲壳动物9种。对照区调查到硬骨鱼类10种、甲壳类5种、棘皮动物3种。
表3列出了地笼网的CPUE和主要物种。鱼礁区的主要渔获物为硬骨鱼类和甲壳动物, 而对照区的主要渔获物为硬骨鱼类和棘皮动物。大泷六线鱼(Hexagrammos otakii)是南、北礁区和对照区的主要渔获物, 在春季丰度较高。南、北礁区大泷六线鱼年均CPUE分别是对照区的1.36倍和4.13倍。日本(Charybdis japonica)是小石岛鱼礁区主要甲壳类经济物种, 年均CPUE分别是对照区的4.52倍和5.75倍。海刺猬(Glyptocidaris crenularis)为对照区主要渔获物, 夏季丰度较高, 占对照区年均CPUE的43.3%, 在鱼礁区渔获物中没有出现。这可能是由于地笼网能够为渔获物提供隐蔽场所[30], 会对海刺猬产生诱集作用, 导致对照区所捕获的海刺猬CPUE较高。而在鱼礁区, 海刺猬能够躲避在鱼礁内部或者缝隙中, 或者发生遮蔽行为, 将贝壳、石头、大型海藻碎片等吸附到反口面上[31-33], 地笼网对海刺猬的吸引作用相对降低, 因此海刺猬不易被地笼网捕获。

表1 鱼礁区和对照区鱼类生物名录Tab. 1 Fish species in the artificial reef areas and the control area
2.2 渔获物优势种
表4列出了不同季节各礁区渔获物的优势种,秋季南、北礁区的优势种均有日本和石鲽, 二者占南礁区秋季CPUE的比例为49.2%和7.5%, 占北礁区秋季CPUE比例为22.3%和2.1%, 北礁区优势种星康吉鳗占北礁区秋季CPUE的比例为61.0%。对照区秋季采集的渔获物小黄鱼、斑和赤鼻棱在此次地笼调查中出现。冬季南、北礁区和对照区渔获物重量远低于其他3个季节, 分别占南、北礁区和对照区全年渔获物总重量的10.2%、9.2%和3.6%。

表2 鱼礁区和对照区甲壳类生物名录Tab. 2 Crustacean species in the artificial reef areas and the control area

表3 鱼礁区和对照区的CPUETab. 3 CPUE of the artificial reef areas and the control area
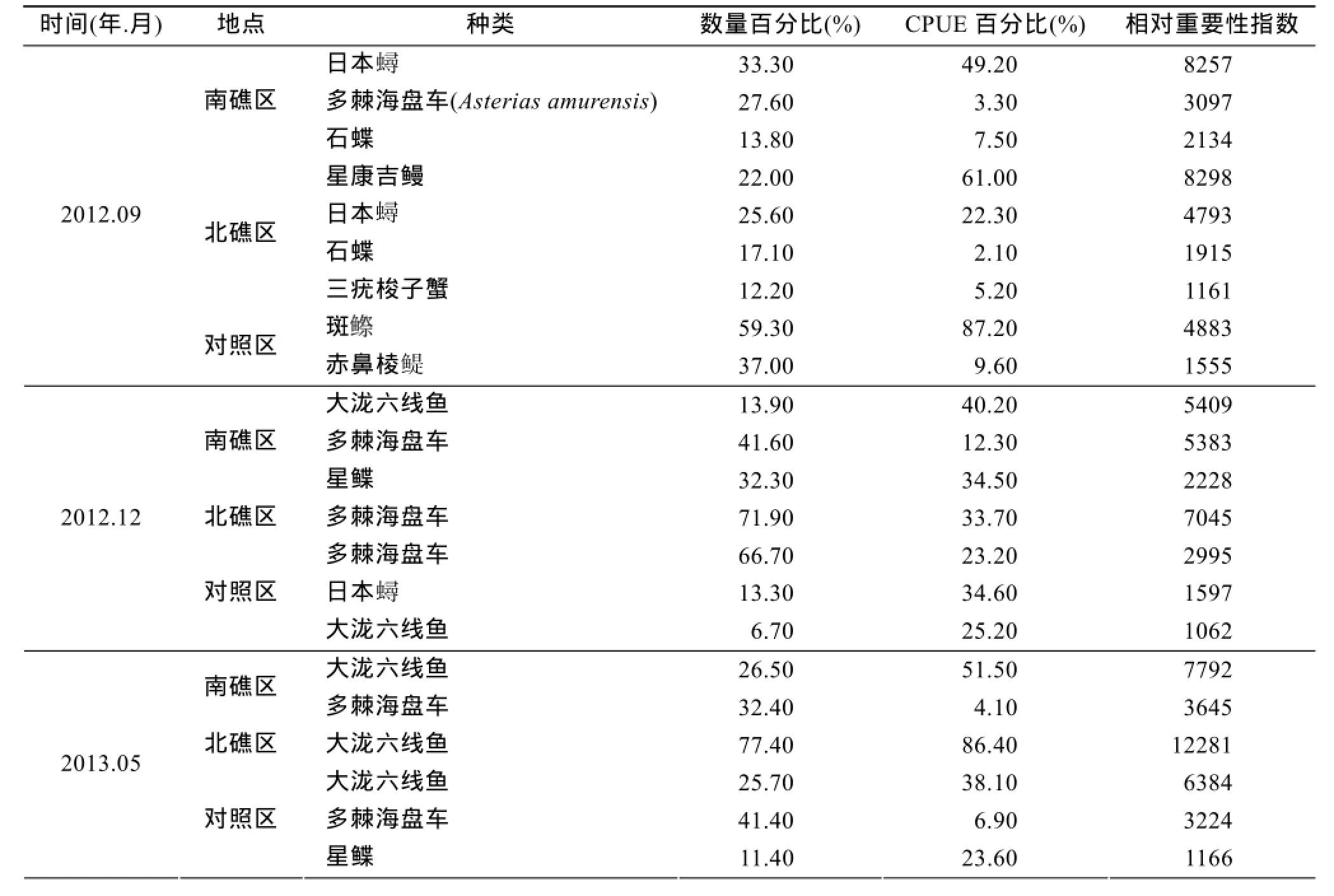
表4 渔获物优势种Tab. 4 Dominant species of catches

续表
大泷六线鱼为南、北礁区和对照区的春季的优势种, 分别占南、北礁区和对照区CPUE的比例为51.5%、86.4%和38.1%, 是春季鱼礁区渔获物的主要种类。许氏平鲉、大泷六线鱼和日本蟳均为南、北礁区夏季的优势种, 分别占南、北礁区渔夏季CPUE的比例为69.4%和88.8%, 对照区夏季的主要优势种为海刺猬, 占对照区夏季CPUE的比例为62.8%, 许氏平鲉、大泷六线鱼和日本蟳仅占对照区夏季CPUE的比例21.76%。

图2 鱼礁区和对照区聚类分析图Fig. 2 Cluster analysis (Bray-Curts) for the artificial reef areas and the control area
2.3 渔获物结构特征
图2为渔获样本聚类分析图, 渔获样本可分为4组, 秋季礁区组、冬季组、春夏礁区组和春夏对照区组。单因子相似性分析(ANOSIM)表明, 渔获物结构组间差异显著(P<0.05)。对照区在2012年秋季只有一个站位, 渔获物由小黄鱼、斑和赤鼻棱组成,这3种鱼类在南、北礁区的渔获物中未出现过, 因此秋季礁区渔获物组成结构与对照区具有差异, 但差异不显著(P=0.16>0.05)。冬季水表温度在5.23~5.40℃,渔获物较少, 因此鱼礁区和对照区渔获物结构没有明显差异。
表5列出了渔获物结构单因子相似性分析结果和对组间差异性贡献率达到前4位的物种。秋季礁区组和冬季组的平均相异性为66.85%。冬季组和春夏礁区组平均相异性为47.36%, 春夏礁区组与春夏对照区组平均相异性为48.60%。春季和夏季造成鱼礁区和对照区群落结构差异贡献率较高的物种为海刺猬、许氏平鲉、日本蟳和大泷六线鱼。海刺猬是春夏对照区组典型种, 日本蟳和许氏平鲉则是春夏礁区组的典型种。

表5 渔获物结构比较Tab. 5 Comparison of catch communities
2.4 鱼礁区主要渔获物丰度与环境因子的相关性
通过对鱼礁区渔获物和环境因子的关系进行排序, 探讨鱼礁区主要物种与环境因子的相关性。表6为各环境因子的平均值, 通过单因素方差分析, 表中环境因子在各季节间差异及其显著(P<0.01)。水温、盐度、pH、DO、COD、叶绿素a、浮游动物细胞密度、浮游植物细胞密度季节差异均极其显著(P<0.01)。
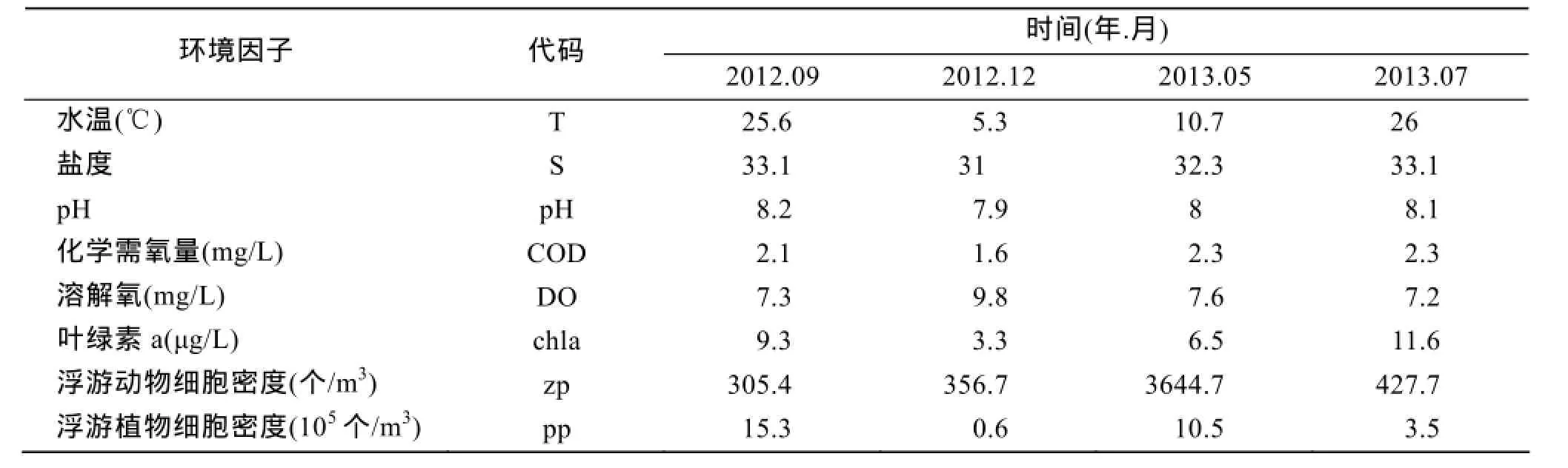
表6 鱼礁区环境因子和浮游生物Tab. 6 Environmental measurements and plankton in the artificial reef areas
图3为环境因子以及浮游生物对主要渔获物影响的CCA排序图。第一轴解释了物种变异的41.6%,第二轴解释了物种变异的26.7%。应用蒙特-卡洛置换检验方法对海水环境因子以及浮游动、植物生物量进行预筛选, 水温因子、叶绿素a、溶解氧含量对渔获物具有显著影响(P<0.05)。对物种变异解释比例依次为36.1%、22.5%和18.9%, 累积为77.5%。水温与第一轴相关性最高, 为-0.8416, 叶绿素a、溶解氧与第二轴相关性最高, 分别为-0.6717和0.6685。
由CCA排序图可知, 在调查海域, 水温与星康吉鳗、石鲽、许氏平鲉、褐菖和日本呈正相关关系, 与星鲽和大泷六线鱼呈负相关关系, 叶绿素a含量主要与石鲽、高眼鲽、许氏平、褐菖鲉和日本呈正相关关系。溶解氧浓度主要与星鲽表现出正相关关系, 冬季礁区溶解氧浓度显著高于其他季节(P<0.01), 而且星鲽为冷温性鱼类, 冬季溶解氧浓度较高的站位CPUE较高。
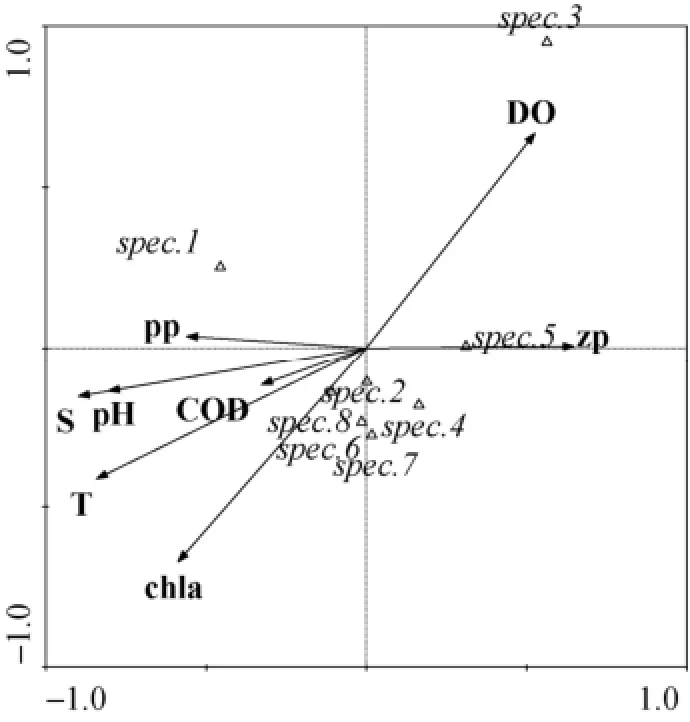
图3 鱼礁区主要渔获物的CCA排序Fig. 3 CCA ordination of the main catches of the artificial reef areas
3 讨论
鱼礁区调查到的渔获种类数和资源量均高于对照区, 南、北礁区渔获种类数分别为对照区的1.44倍和1.89倍, 南、北礁区的鱼类、软体动物和甲壳动物的资源量分别为对照区的1.37倍和2.40倍。表明投放人工鱼礁对渔业生物资源的诱集和恢复具有积极影响, 使附近海域鱼类种群丰富和多元化。这也与三横山人工鱼礁区、獐子岛海洋牧场、荣成俚岛人工鱼礁区的资源调查结果一致[10-11,34]。人工鱼礁产生的上升流能够将底部营养盐带入表层, 能够促进海洋浮游植物、藻类等繁殖, 从而促进其他海洋生物的增殖; 礁体产生的背涡流通常流速很小, 能够为海洋动物提供庇护, 吸引海洋动物前来[35]。南、北礁区地理位置存在差异, 根据刘彦等[36]研究, 礁体产生的最大上升流速度会随着来流速度的增加而增大, 北礁区流速较快, 会产生较大的最大上升流, 从而形成较好的流场效应, 这可能是造成北礁区资源量高于南礁区的原因之一。
2009年春季在小石岛附近人工鱼礁的调查显示,礁区仅存在大泷六线鱼、许氏平鲉和褐菖鲉3种鱼类[37]。本次调查结果显示2013年春季人工鱼礁区出现的鱼类种类数多于2009年春季调查调查结果, 这表明, 随着时间的推移, 人工鱼礁区聚集的鱼类种类数目增加。2013年春季在人工鱼礁区共发现9种鱼类, 除了大泷六线鱼、许氏平鲉和褐菖鲉, 还有石鲽、星鲽和高眼鲽等岩礁鱼类。随着海底生态环境的修复, 礁区渔业资源结构也会不断改善, 人工鱼礁对资源的修复是一个漫长的过程。
地笼网被放置在海底, 目标渔获物大多是底栖生物,如底层鱼类和甲壳动物等。硬骨鱼类和甲壳动物是南、北礁区渔获物的主要组成部分, 棘皮动物次之, 软体动物最低, 并且鱼类渔获物主要为底层鱼类, 有大泷六线鱼、石鲽、许氏平鲉等。这与徐浩等[38]在莱州朱旺港人工鱼礁区调查一致, 可能与使用地笼网进行调查有关。
南、北礁区和对照区春季的主要经济鱼种为大泷六线鱼, 并且礁区大泷六线鱼的春季CPUE远高于对照区。冬季黄海北部水温较低, 经济鱼虾远离近岸,对照区鱼类仅有少量大泷六线鱼40.41 g/(d·网), 在南、北礁区尚存在星鲽、裸虾虎鱼、花鲈等物种。威海小石岛鱼礁区的经济类甲壳动物为日本日本是南、北礁区夏季的主要优势种, CPUE远高于对照区。
威海小石岛鱼礁区地笼网渔获物组成结构季节间差异明显, 按季节变化可分为秋季礁区组、冬季组、春夏礁区组和春夏对照区组。在春季和夏季, 鱼礁区群落结构与对照区差异明显, 南礁区和北礁区渔获物群落结构差异性不明显, 表明礁区之间, 较礁区与对照区之间具有更加相似的群落结构。造成春夏鱼礁区与春夏对照区差异的主要分歧种为海刺猬和日本蟳,其中海刺猬是夏季对照区的典型种。棘皮动物海刺猬会因为光照、捕食者等因素影响发生遮蔽行为, 人工鱼礁及附着在其上的藻类等能够为海刺猬提供遮蔽物或掩蔽物, 可能会使得海刺猬较难被捕获。有研究表明长期生活在没有遮蔽物情况下的海刺猬遮蔽强度相应较低[31], 对照区日本等捕食者较少, 可能会导致海刺猬遮蔽行为相对较少, 并且地笼网能够为渔获物提供隐蔽场所, 会对对照区海刺猬产生诱集作用,从而使得对照区渔获物中出现较多的海刺猬。
以往有研究表明水深、水温和盐度是影响鱼类分布的主要限制因素[10]。小石岛鱼礁区平均水深约为10 m,变化范围不大, 因此未对水深进行排序分析。威海小石岛鱼礁区鱼类分布主要受水温、叶绿素a和溶解氧浓度影响。CCA排序图显示星鲽和大泷六线鱼与水温成负相关, 在鱼礁区, 星鲽出现在冬季和春季, 冬季CPUE较高, 大泷六线鱼全年均有出现, 春季CPUE最高, 夏季和秋季CPUE较低, 与星鲽和大泷六线鱼为冷温性鱼类相一致。石鲽分布与水温成微弱正相关, 在鱼礁区, 石鲽CPUE在春季和秋季较高, 秋季CPUE最高,与石鲽在秋季和早春产量较大一致。许氏平鲉为近海底层鱼类, 营附礁性生活, 研究显示2012年9月和2013年7月份丰度高于冬季和春季, 可能是因为许氏平鲉在春季产卵后, 在礁区育肥, 为越冬储存能量[34]。研究发现星鲽与溶解氧浓度呈正相关关系, 冬季主要出现在礁区溶解氧较高的站位, 表明在人工鱼礁区,星鲽的分布受温度和溶解氧浓度影响。王小荟[39]研究发现冷温性鱼类分布与底层溶解氧浓度相关, 张迎秋等[40]也研究发现大头鳕(Gadus macrocephalus)等冷温性鱼类多分布在低温季节深水高溶解氧区。海水中叶绿素a是浮游植物进行光合作用的主要色素, 夏秋季节叶绿素a浓度较高, 也会造成人工鱼礁区海洋初级生产力升高, 进而造成其他海洋动物丰度增高, 因此叶绿素对渔获物物种变异解释程度较高。而且叶绿素a的浓度受到海水温度的影响, 在人工鱼礁区变化特征与水温一致, 表现出明显的季节变化特征, 夏季高于秋季高于春季高于冬季, 因此与许氏平鲉等夏秋季节CPUE较高的渔获物表现出相关性。
[1] 徐宾铎, 金显仕, 梁振林. 秋季黄海底层鱼类群落结构的变化[J]. 中国水产科学, 2003, 10(2): 148-154. Xu Binduo, Jin Xianshi, Liang Zhenlin. Changes of demersal fish community structure in the Yellow Sea during the autumn[J]. Journal of Fishery Sciences of China, 2003, 10(2): 148-154.
[2] 董婧, 刘海映, 许传才, 等. 黄海北部近岸鱼类的群落结构[J]. 大连水产学院学报, 2004, 19(2): 132-137. Dong Jing, Liu Haiying, Xu Chuancai, et al. Community structure of fish in the coast of northern Yellow Sea[J]. Journal of Dalian Fisheries University, 2004, 19(2): 132-137.
[3] Gao Q F, Shin P K S, Xu W Z, et al. Amelioration of marine farming impact on the benthic environment using artificial reefs as biofilters[J]. Marine Pollution Bulletin, 2008, 57(6-12): 652-661.
[4] Fukunaga A, Bailey-Brock J H. Benthic infaunal communities around two artificial reefs in Mamala Bay, Oahu, Hawaii[J]. Marine Environmental Research, 2008, 65(3): 250-263.
[5] Santos M N, Monteiro C C. Comparison of the catch and fishing yield from an artificial reef system and neighbouring areas off Faro (Algarve, south Portugal)[J]. Fisheries Research, 1998, 39(1): 55-65.
[6] Pickering H, Whitmarsh D, Jensen A. Artificial reefs as a tool to aid rehabilitation of coastal ecosystems: investigating the potential[J]. Marine Pollution Bulletin, 1999, 37(8-12): 505-514.
[7] Falcão M, Santos M N, Drago T, et al. Effect of artificial reefs (southern Portugal) on sediment-water transport of nutrients: Importance of the hydrodynamic regime[J]. Estuarine, Coastal and Shelf Science, 2009, 83(4): 451-459.
[8] 张硕, 朱孔文, 孙满昌. 海州湾人工鱼礁区浮游植物的种类组成和生物量[J]. 大连水产学院学报, 2006, 21(2): 134-140. Zhang Shuo, Zhu Kongwen, Sun Manchang. Species composition and biomass variation in phytoplankton in artificial reef area in Haizhou Bay[J]. Journal of Dalian Fisheries University, 2006, 21(2): 134-140.
[9] 孙习武, 张硕, 赵裕青, 等. 海州湾人工鱼礁海域鱼类和大型无脊椎动物群落组成及结构特征[J]. 上海海洋大学学报, 2010, 19(4): 505-513. Sun Xiwu, Zhang Shuo, Zhao Yuqing, et al. Community structure of fish and macroinvertebrates in the artificial reef sea area of Haizhou Bay[J]. Journal of Shanghai Ocean University, 2010, 19(4): 505-513.
[10] 吴忠鑫, 张磊, 张秀梅, 等. 荣成俚岛人工鱼礁区游泳动物群落特征及其与主要环境因子的关系[J]. 生态学报, 2012, 32(21): 6737-6746. Wu Zhongxin, Zhang Lei, Zhang Xiumei, et al. Nekton community structure and its relationship with main environmental variables in Lidao artificial reef zones of Rongcheng[J]. Acta Ecologica Sinica, 2012, 32(21): 6737-6746.
[11] 汪振华, 章守宇, 王凯, 等. 三横山人工鱼礁区鱼类和大型无脊椎动物诱集效果初探[J]. 水产学报, 2010, 34(5): 751-759. Wang Zhenhua, Zhang Shouyu, Wang Kai, et al. A preliminary study on fish and macroinvertebrate enhancement in artificial reef area around Sanheng Isle, Shengsi, China[J]. Journal of Fisheries of China, 2010, 34(5): 751-759.
[12] 赵静, 章守宇, 汪振华, 等. 嵊泗人工鱼礁海域鱼类和大型无脊椎动物群落结构特征[J]. 中国水产科学, 2010, 17(5): 1045-1056. Zhao Jing, Zhang Shouyu, Wang Zhenhua, et al. Analysis on community structure and diversity of fish and macroinvertebrate in Shengsi artificial reef area[J]. Journal of Fishery Sciences of China, 2010, 17(5): 1045-1056.
[13] 廖一波, 曾江宁, 寿鹿, 等. 象山港人工鱼礁投放对大型底栖动物群落结构的影响[J]. 海洋与湖沼, 2014,45(3): 487-495. Liao Yibo, Zeng Jiangning, Shou Lu, et al. Impact of artificial reef on macrobenthic community structure in Xiangshan Bay[J]. Oceanologia et Limnologia Sinica, 2014, 45(3): 487-495.
[14] 高东奎, 赵静, 张秀梅, 等. 莱州湾人工鱼礁区及附近海域鱼卵和仔稚鱼的种类组成与数量分布[J]. 中国水产科学, 2014, 21(2): 369-381. Gao Dongkui, Zhao Jing, Zhang Xiumei, et al. Species composition and distribution patterns of ichthyoplankton within and outside artificial reefs in Laizhou Bay[J]. Journal of Fishery Sciences of China, 2014, 21(2): 369-381.
[15] Addis D T, Patterson W F III, Dance M A, et al. Implications of reef fish movement from unreported artificial reef sites in the northern Gulf of Mexico[J]. Fisheries Research, 2013, 147(10): 349-358.
[16] Henderson M J, Fabrizio M C, Lucy J A. Movement patterns of summer flounder near an artificial reef: Effects of fish size and environmental cues[J]. Fisheries Research, 2014, 153(5): 1-8.
[17] Abecasis D, Bentes L, Lino P G, et al. Residency, movements and habitat use of adult white seabream (Diplodus sargus) between natural and artificial reefs[J]. Estuarine, Coastal and Shelf Science, 2013, 118: 80-85.
[18] 汪振华, 章守宇, 王凯. 三横山鱼礁生境鱼类和大型无脊椎动物群落特征[J]. 生态学报, 2010, 30(8): 2026-2035. Wang Zhenhua, Zhang Shouyu, Wang kai. Fish and macroinvertebrates community structure in artificial habitat around Sanheng Isle, Shengsi, China[J]. Acta Ecologica Sinica, 2010, 30(8): 2026-2035.
[19] Godoy E A S, Almeida T C M, Zalmon I R. Fish assemblages and environmental variables on an artificial reef north of Rio de Janeiro, Brazil[J]. Ices Journal of Marine Science, 2002, 59(5): S138-S143.
[20] Zalmon I R, Saleme De Sá F, Neto E J D, et al. Impacts of artificial reef spatial configuration on infaunal community structure — Southeastern Brazil[J]. Journal of Experimental Marine Biology and Ecology, 2014, 454(5): 9-17.
[21] 章守宇, 张焕君, 焦俊鹏, 等. 海州湾人工鱼礁海域生态环境的变化[J]. 水产学报, 2006, 30(4): 475-480. Zhang Shouyu, Zhang Huanjun, Jiao Junpeng, et al. Change of ecological environment of artificial reef waters in Haizhou Bay[J]. Journal of Fisheries of China, 2006, 30(4): 475-480.
[22] 中国国家标准化管理委员会. GB/T17378-2007, 海洋监测规范[S]. 北京: 中国标准出版社, 2007. Standardization Administration of the People’s Republic of China. GB/T12763-2007, Specifications for Marine Monitoring[S]. Beijing: Standards Press of China, 2007.
[23] 中国国家标准化管理委员会. GB/T12763-2007, 海洋调查规范[S]. 北京: 中国标准出版社, 2007. Standardization Administration of the People’s Republic of China. GB/T12763-2007, Specifications for Oceanographic Survey[S]. Beijing: Standards Press of China, 2007.
[24] Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, bluefin tuna and bonito in Californian waters[J]. Fish Bull, 1971, 152: 5-10.
[25] Burt J A, Feary D A, Cavalcante G, et al. Urban breakwaters as reef fish habitat in the Persian Gulf[J]. Marine Pollution Bulletin, 2012, 72(2): 342-350.
[26] 周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J]. 青岛海洋大学学报(自然科学版), 2003, 33(1): 58-64. Zhou Hong, Zhang Zhinan. Rationale of the multivariate Statistical Software PRIMER and its app; ication in benthic community ecology[J]. Journal of Ocean University of Qingdao, 2003, 33(1): 58-64.
[27] Lepx J, Smilauer P. Multivariate Analysis of Ecological Data Using CANOCO[M]. UK: Cambridge University Press, 2003: 151-165.
[28] 赖江山. 生态学多元数据排序分析软件Canoco5介绍[J]. 生物多样性, 2013, 21(6): 765-768. Lai Jiangshan. Canoco 5: a new version of an ecological multivariate data ordination program[J]. Biodiversity Science, 2013, 21(6): 765-768.
[29] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004. Zhang Jintun. Quantitative Ecology[M]. Beijing: Science Press, 2004.
[30] 孙满昌. 海洋渔业技术学[M]. 北京: 中国农业出版社, 2005. Sun Manchang. Marine Fisheries Technology[M]. Beijing: China Agriculture Press, 2005.
[31] James D W. Diet, movement, and covering behavior of the sea urchin Toxopneustes roseus in rhodolith beds in the Gulf of California, México[J]. Marine Biology, 2000, 137(5-6): 913-923.
[32] Dumont C P, Drolet D, Deschênes I, et al. Multiple factors explain the covering behaviour in the green sea urchin, Strongylocentrotus droebachiensis[J]. Animal Behaviour, 2007, 73(6): 979-986.
[33] 常亚青, 李云霞, 罗世滨, 等. 不同实验生态环境对海刺猬遮蔽行为的影响[J]. 生态学报, 2013, 33(9): 2754-2760. Chang Yaqing, Li Yunxia, Luo Shibin, et al. Effects of different ecological environments in the laboratory on the covering behavior of the urchin Glyptocidaris crenularis[J]. Acta Ecologica Sinica, 2013, 33(9): 2754-2760.
[34] 陈勇, 杨军, 田涛, 等. 獐子岛海洋牧场人工鱼礁区鱼类资源养护效果的初步研究[J]. 大连海洋大学学报, 2014, 29(2): 183-187. Chen Yong, Yang Jun, Tian Tao, et al. The enhancement effect of fishery resources on artificial reefs in marine ranching area in Zhangzi Island[J]. Journal of Dalian Ocean University, 2014, 29(2): 183-187.
[35] 崔勇, 关长涛, 万荣, 等. 人工鱼礁流场效应的数值模拟与仿真研究[J]. 系统仿真学报, 2009, 21(23):7393-7396. Cui Yong, Guan Changtao, Wan Rong, et al. Research and numerical simulation on features of flow field around artificial reefs[J]. Journal of System Simulation, 2009, 21(23): 7393-7396.
[36] 刘彦, 赵云鹏, 崔勇, 等. 正方体人工鱼礁流场效应试验研究[J]. 海洋工程, 2012, 30(4): 103-108. Liu Yan, Zhao Yunpeng, Cui Yong, et al. Experimental study of the flow field round cube artificial reef[J]. The Ocean Engineering, 2012, 30(4): 103-108.
[37] 焦金菊, 潘永玺, 孙利元, 等. 人工鱼礁区的增殖鱼类资源效果初步研究[J]. 水产科学, 2011, 30(2): 79-82. Jiao Jinju, Pan Yongxi, Sun Liyuan, et al. Effect of artificial reefs on fish multiplication[J]. Fisheries Science, 2011, 30(2): 79-82.
[38] 徐浩, 曾晓起, 顾炎斌, 等. 人工鱼礁对山东莱州朱旺港海区游泳动物的群落结构及季节变化的影响[J].中国海洋大学学报(自然科学版), 2012, 42(5): 47-54. Xu Hao, Zeng Xiaoqi, Gu Yanbin, et al. The effect of artificial reefs of community structure and seasonal variation of nektons in Zhuwang Habor, Laizhou, Shangdong[J]. Periodical of Ocean University of China, 2012, 42(5): 47-54.
[39] 王小荟. 海州湾主要鱼种的空间分布及其与环境因子的关系[D]. 青岛: 中国海洋大学, 2013. Wang Xiaohui. Spatial distribution of dominant fish species in Haizhou Bay and their relationships with environmental factors[D]. Qingdao: Ocean University of China, 2013.
[40] 张迎秋, 许强, 徐勤增, 等. 海州湾前三岛海域底层鱼类群落结构特征[J]. 中国水产科学, 2016, 23(1): 156-168. Zhang Yingqiu, Xu Qiang, Xu Qinzeng, et al. Demersal fish community structure around the Qiansan Islets in Haizhou Bay[J]. Journal of Fishery Sciences of China, 2016, 23(1): 156-168.
Received: Nov. 23, 2015
Catch community structure characteristics and their relationships with environmental factors in the Xiao Shidao artificialreef sea area of Weihai
WANG Xin-meng, TANG Yan-li, SUN Xiao-mei, SHENG Hua-xiang, WAN Rong
(College of Fisheries, Ocean University of China, Qingdao 266003, China)
Artificial reef; fishery resource; community structure; CCA; environmental factors
Fishery resources and environmental elements in the Xiao Shidao artificial reef area and its control area were investigated during 2012-2013 to identify the influence of artificial reefs on species as well as catch community structure characteristics, which were analyzed using the CLUSTER, one-way ANOSIM, and SIMPER methods. The correlation between the dominant species and environmental factors was analyzed using CCA methods. The cage net survey results showed that the number of species in the southern and northern reef areas was higher than that in the control areas 18, 24, and 31. The dominant fish species throughout the year in the southern and northern reef areas were Charybdis japonica and Hexagrammos otakii. The dominant species in the control area were Glyptocidaris crenularis and Hexagrammos otakii. The size of the fish and crustacean catches in the southern and northern reef areas were higher than that in the control area. CLUSTER and ANOSIM analyses indicated significant differences in catch community structures both among seasons (P<0.05) and between the artificial reef areas and the control area during spring and summer (P<0.05). CCA analysis showed that the major environmental factors affecting the distribution of dominant species in reef areas were water temperature, chlorophyll, and dissolved oxygen. In conclusion, construction of artificial reefs ameliorated the catch community structure of the Xiao Shidao artificial reef areas and increased the catch abundance of the dominant economic fish and crustacean species.
S953.1
A
1000-3096(2016)11-0034-10
10.11759//hykx20151123001
(本文编辑: 谭雪静)
2015-11-23;
2016-05-25
海洋公益性行业专项(201305030); 公益性行业(农业)科技专项(201203018)
[Foundation: Marine Public Welfare Industry, No.201305030; Special Public Welfare Industry (Agriculture) Research, No. 201203018]
王新萌(1990-), 女, 山东临沂, 硕士研究生, 主要从事渔业资源研究, 电话: 15165252397, E-mail: wangxinmeng9012@163.com;唐衍力, 通信作者, 电话: 0532-82031076, E-mail: tangyanli@ouc.edu.cn
猜你喜欢
中国水产(2021年6期)2021-07-01
海洋通报(2020年5期)2021-01-14
农家之友(2020年9期)2020-04-21
浙江海洋大学学报(自然科学版)(2019年5期)2019-03-21
渔业现代化(2018年6期)2018-12-28
珠江水运(2018年21期)2018-12-20
海洋渔业(2018年5期)2018-10-26
中国海洋大学学报(自然科学版)(2015年8期)2015-03-18
