
牙鲆精氨酸酶基因片段的克隆及序列分析
2016-02-08 05:22施婷婷施志仪
广东农业科学 2016年12期
刘 华,施婷婷,施志仪
(1.南宁富莱欣生物科技有限公司研发中心,广西 南宁 530003;2.上海海洋大学生物技术研究中心,上海 200090)
牙鲆精氨酸酶基因片段的克隆及序列分析
刘 华1,施婷婷1,施志仪2
(1.南宁富莱欣生物科技有限公司研发中心,广西 南宁 530003;2.上海海洋大学生物技术研究中心,上海 200090)
根据GenBank中虹鳟、大菱鲆、斑马鱼及其他生物的精氨酸酶基因序列设计并合成一对引物,并提取变态期牙鲆仔鱼总RNA作为模板进行RT-PCR,得到一条cDNA片段。将产物连接到T载体中,转化、扩增重组质粒。提取大肠杆菌中的重组质粒进行DNA测序。将DNA测序所得到的基因序列翻译成蛋白序列和已知的大菱鲆、鲤鱼、虹鳟、斑马鱼的精氨酸酶基因序列进行同源性分析。结果显示,利用RTPCR的方法扩增出一条284 bp的目的基因片段,比对结果发现,其与大菱鲆、鲤鱼、斑马鱼以及虹鳟精氨酸酶基因蛋白序列的同源性分别为96%、85%、84%和82%。对蛋白序列进行生物信息学分析发现,整个蛋白序列中无信号肽,无糖基化位点,无跨膜结构域,但含有一个典型的arginase结构。在N-端第38~44、57~64、71~76、89~94区段有β-折叠中心;第7~18、50~52、81~87区段可能形成α螺旋区域。Arg蛋白N端第37~45和第74~77区段为优势抗原表位区域。
牙鲆;精氨酸酶;克隆;序列分析
精氨酸酶(Arginase)是在细菌、酵母、植物、无脊椎动物和脊椎动物中普遍存在的一种酶,该酶不仅参与机体尿素循环,还与机体病理过程密切相关。在植物和排氨动物中精氨酸酶位于线粒体中,而在排尿动物中精氨酸酶位于细胞质中。目前,精氨酸酶基因在香鱼、虹鳟和鲤鱼等少数硬骨鱼类中被克隆和鉴定。健康香鱼 Arg-II mRNA主要在肝、脑和单核/巨噬细胞中表达,感染鳗弧菌后,Arg-II mRNA表达显著上调,Arg-II与香鱼抗病原感染的免疫反应紧密相关[1]。肝细胞癌(hepatocellular carcinoma,HCC)中Arg-1表达降低,Arg-1与HCC分化、转移及复发呈显著相关,提示Arg-1对HCC发生、进展发挥负性调控作用,Arg-1可作为评估患者预后的参考指标[2]。迄今为止,在蛋白质结构数据库(Protein Data Bank,PDB)中还没有关于鱼类精氨酸酶晶体结构的报道。目前研究主要集中在精氨酸酶的酶学性质和催化机制。
目前,国内外尚未见有对牙鲆精氨酸酶的研究,因此我们对牙鲆精氨酸酶基因部分序列进行了克隆和表达研究,研究结果将为重组牙鲆精氨酸酶的表达提供可靠的基因材料,同时也为进一步研究该基因的结构和功能及与酶激活因子之间的作用方式奠定基础。
1 材料与方法
1.1 试验材料
供试牙鲆仔鱼,购自山东良种示范基地,运回实验室海水暂养,水温控制在21(±0.5)℃。取27日龄变态期牙鲆仔鱼50条,分别去掉肠,液氮速冻,立即进行总RNA提取。
其他主要试验材料包括TRIzol Reagent、Taq酶、RNasin、M-MLV逆转录酶、Oligo(dT)18 primer、pUcm-T Vector、DNA Marker;主要仪器设备包括冷冻离心机、PCR仪、移液枪、电泳仪。
1.2 试验方法
1.2.1总RNA提取 根据TRizol试剂法抽提牙鲆仔鱼总RNA,按其产品说明书进行操作提取。
1.2.2设计引物与反转录-聚合酶链反应 从 GenBank数据库中查找虹鳟、大菱鲆、斑马鱼等生物的Arginase基因序列设计并合成聚合酶链反应5’端引物:5’-GGA AAC CTC CAC GGC CAG CC-3’(20 bp),3’端引物:5’-TCA AAG CTC AGG TGG AT-3’(17 bp)。依据试剂盒方法进行cDNA的合成和扩增。在0.2 mL EP中依次加入1 µL(1 µg/µL)总RNA、2 µL Oligo(dT)18 primer、20 U(0.5 µL)RNasin、2 µL dNTPs、4 µL 5×Reverse Transcriptase Buffer、10 U AMV逆转录酶、8.5 µL DEPC处理水,总体积20 µL。于室温下放10 min再转入42℃恒温槽温浴l h。在逆转录产物中吸取0.5 µL进行PCR,反应程序为94℃预变性1 min,再依据如下循环参数进行PCR反应:94℃ 45 s,51.1℃45 s,72℃ 45 s,35个循环后,72℃ 4 min。PCR反应结束后,吸取5 µL PCR产物进行1%琼脂糖凝胶电泳。
1.2.3牙鲆Arginase基因的克隆、鉴定和测序 Arginase基因片段通过电泳回收,吸取7 µL Arginase基因PCR纯化产物,依次加入1 µL pUcm-T Vector、1 µL Ligation Buffer、1 µL T4 DNA连接酶,总体积10 µL。连接反应条件:16℃,12 h。吸取2 µL反应终产物转化50 µL DH5α感受态细胞。已转化的感受态细胞在LB琼脂平板(含X-Gal)培养基上培养(37℃,20 h)。选取白色菌落接种于LB液体培养基进行培养,PCR法、酶切法进一步鉴定筛选阳性菌落,阳性菌送上海生物工程有限公司测序。双酶切体系为20 µL,分两管酶切,内切酶EcoRⅠ、XhoⅠ/BamHⅠ各1.5 µL,10×Buffer 2 µL,抽提的pUcm-arg284质粒15 µL,离心混匀后,37℃酶切过夜。
1.2.4生物信息学分析 利用SWISS-PROT、PROSITE、EMBL等蛋白数据库和DNAStar、Bioedit、ClustalX1.8、Mega2等软件及在线网络服务器分析牙鲆Arginase蛋白序列特征,并与GenBank中的物种序列进行同源性比较。
2 结果与分析
2.1 RT-PCR扩增结果
根据设计的目的基因上下游引物的活性温度,以50~55℃为温度梯度,以18S引物作阳性对照进行RT-PCR,将所得产物进行琼脂糖电泳,结果显示18S引物扩增出来的产物大小为315 bp,但其后500~750 bp区域有一条较弱的带,而目的基因引物扩增出来的产物大小约为280 bp(图1)。

图1 RT-PCR产物电泳结果
2.2 牙鲆Arginase基因cDNA片段的阳性克隆鉴定
从转化的DH5α进行菌液鉴定,见图2;菌株提取质粒DNA,利用目的基因的引物进行PCR,扩增得到一条与设计的目的基因长度相同的DNA片段,可以确定目的基因已经插入T载体,得到阳性克隆(图3)。EcoRⅠ、XhoⅠ/BamHⅠ双酶切均能切除280 bp左右大小的片段,酶切鉴定见图4,说明抽提的质粒含有插入目的片段。

图2 LB菌液PCR鉴定结果
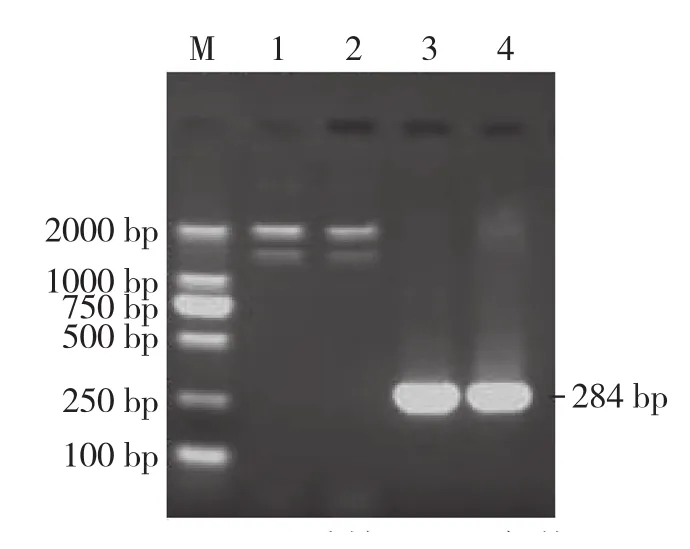
图3 重组质粒PCR鉴定结果

图4 重组子限制性内切酶酶切电泳结果

2.3 牙鲆Arginase基因cDNA片段测序结果
将提取的重组质粒送基因公司测序,以M13Primers为测序引物测序得到目的基因序列(284 bp):
将核苷酸测序结果输入EXPASY网站(http://www.expasy.org/tools/dna.html)进行翻译,得到该基因的编码氨基酸序列(94 aa):-GNLHGQPVAFMLKELQDKMPAIPGFSWVKPFL SSRDLVYIGLRDVDPGERHILKNLGIQYFTMRDI DRLGIQRVMEVTLDHLMARKQRPIHLSF-
应用NCBI网上的BLAST程序与GenBank、EMBL、DDBJ及PDB数据库上的序列进行同源性比较,结果表明,所克隆的牙鲆精氨酸酶与同属大菱鲆(Q71LX3,Scophthalmus maximus)同源性最高,其中与大菱鲆精氨酸酶基因的碱基序列及氨基酸序列的同源性分别为91%和96%;与鲫鱼(Q4A4U9,Cyprinus carpio)、斑马鱼(Q6PH54,Danio rerio)、虹鳟(Q7T1U6,Oncorhynchus mykiss)碱基序列及推测的氨基酸序列同源性分别为78%/85%、84%/84%和80%/82% 。与人(P78540,Homo sapiens human)的氨基酸序列同源性为72%;与红原鸡(XP_421191,Gallus gallus)的为72%。
GenBank搜索表明该片段是未登录的新基因,将所得到的目的基因的蛋白质序列在http://www.ncbi.nlm.nih.gov网站上进行BLAST分析,分析结果显示目的基因与奥尼罗非鱼、金头鲷、底鳉、斑马鱼精氨酸酶基因蛋白序列的同源性分别为85%、83%、81%、71%,并且目的基因蛋白序列具有典型的Arginase特征结构(图5),可以推断所克隆的序列为牙鲆精氨酸酶基因。

图5 Arginase特征结构
2.4 生物信息学分析
2.4.1Arginase结构特点剖析 运用Tmpred服务器剖析,没有出现跨膜区域;经亮氨酸富集核输出信号剖析,没有发现亮氨酸富集核输出信号;经信号肽剖析没有信号肽出现;经糖基化位点剖析该蛋白没有出现糖基化位点。
对牙鲆Arginase蛋白的内在扰乱区域进行分析(http://dis.embl.de/),结果显示,该蛋白没有Remark-465扰乱,但在N-端第42~50(LRDVDPGER)、56~66(LGIQY FTMRDI)和84~94(ARKQRPI HLSF)3个肽段内同时发现环热环扰乱和一卷曲扰乱,这3个肽段区域很不稳定(图6)。
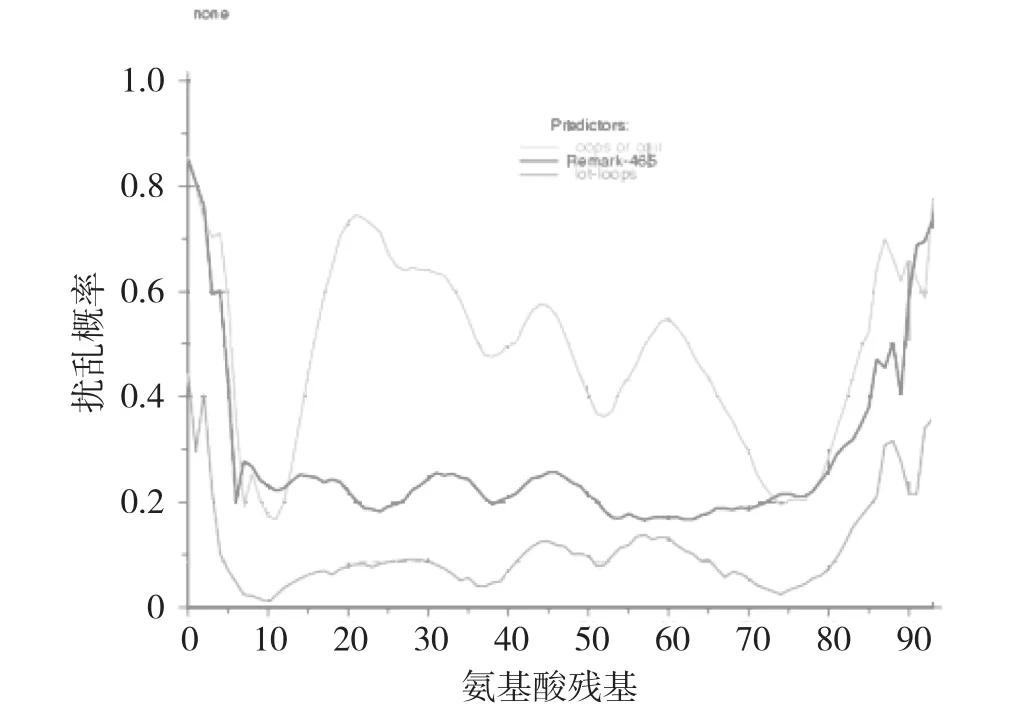
图6 牙鲆Arginase蛋白内在扰乱区域分析
2.4.2Arginase二级结构分析结果 对牙鲆Arginase蛋白二级结构中的α螺旋、β折叠和转角区域同时运用Chou-Fasman方法和Garnier-Robson方法分析,结果(图7、图8)发现,大部分区域重叠。α螺旋区域分析显示,牙鲆Arginase蛋白在N-端第7~18、50~52、81~87这3个肽段内形成共同区域;β折叠区域所在区段分析显示,牙鲆Arginase蛋白在N-端第38~44、57~64、71~76、89~94这4个肽段内形成共同区域;Coil卷曲所在区段分析显示,牙鲆Arginase蛋白在N-端2~5、33~34、45~47这3个肽段内形成共同区域。柔性区域运用Karplus-Schulz方法进行分析发现,柔性区域位于N-端第5~7、14~18、30~36、43~50、64~68、85~89这6个肽段内。
2.4.3Arginase抗原表位预测结果 运用Kyte-Doohttle方案、Jameson-Wolf方案和Emini方案分别预测氨基酸亲水性肽段(1~5,13~21,31~33,43~53,63~69,71~72,76~77,82~91)、抗原指数肽段(1~4,14~20,31~37,43~53,64~71,73~75,82~89)和表面可能性肽段(14~18,44~49,51~52,61~62,64~66,84~8),3个预测指数形成的共同区域均位于牙鲆Arginase蛋白N-端第14~18、44~49、51~52、64~66、84~88这5个肽段内(图9)。疏水性残基往往位于蛋白质分子的核心处,而亲水性残基则主要分布于蛋白质分子的表面,与水环境相互作用。抗原表位的选择是制备合成肽的关键,在这些共同肽段内或附近区域很可能是抗原表位的优势区域。

图7 牙鲆Arginase蛋白二级结构分析结果

图8 牙鲆Arginase蛋白二级结构模型分析结果
使用EMBOSS软件包中的ANTIGENIC程序预测Arginase抗原表位,结果如下(按打分排名从高到低依次排列):22~45(IPGFSWVKPFLSS RDLVYIGLRDV)1.135,5~13(GQPVAFMLK) 1.108,74~82(VMEVTLDHL)1.088,53~61(LKNLGIQYF)1.065。

图9 牙鲆Arginase抗原表位预测结果
用ProPred预测能与大多数MHCⅡ类等位结合的肽段(图10),预测到较好的结合肽分别是:69~77(38/51)、37~45(32/51)、63~68(25/51),括号内数字表示可结合的MHCⅡ类等位基因数/进行结合预测的全部MHCⅡ类等位基因数。
在MHCⅡ类结合肽和抗原表位的预测结果中发现,牙鲆Arginase蛋白37~45和74~77均出现在两次预测的结果中,表明它们可能既是抗原表位又是MHC结合肽。
2.4.4系统进化树的构建及同源性比较 利用GenBank数据库、SWISS-PROT数据库和EMBL数据库,搜索到以下14种生物的Arginase蛋白序列:大菱鲆(Q71LX3,Scophthalmus maximus)、虹鳟(Q7T1U6,Oncorhynchus mykiss)、斑马鱼(Q6PH54,Daniorerio)、鲤鱼(Q4A4U9,Cyprinus carpio)、牛蛙(AAA68073,Rana catesbeiana)、红原鸡(XP_421191,Gallus gallus)、兔(Q4VK78,Oryctolagus cuniculus)、羊(Q30DW3,Ovis aries)、褐家鼠(O08701,Rattus norvegicus)、马(Q6DTK8,Equus caballus)、小家鼠(Q8R1Q7,Mus musculus)、牛(Q58DL1,Bos taurus)、狗(XP_537488,Canis familiaris)、人(P78540,Homo sapiens human)。用ClustalX1.8进行序列比对,再用Mega2构建系统进化树。从构建的系统进化树(图11)中可看出,牙鲆精氨酸酶与大菱鲆、虹鳟、斑马鱼及鲤鱼的同源性高,与两栖类牛蛙同源性较低,而与哺乳类的人、马等同源性最低。

图10 牙鲆Arginase MHCⅡ类等位结合的肽段

图11 不同动物精氨酸酶基因构建的系统进化树
3 讨论
在玉米中,精氨酸酶是氮素循环中催化精氨酸生成鸟氨酸和尿素的重要酶,可促进氮素的再利用,提高氮素的利用效率[3]。在人体中,精氨酸是肝癌细胞繁殖必需的酵素,而精氨酸酶是一种天然酵素,具有分解精氨酸的功能,有助于抑制癌细胞生长,精氨酸酶1在食管癌患者外周血中的高表达与MDSCs水平和食管癌的发生、发展有着密切关系[4]。目前,国外实验室利用cDNA克隆技术已经获得了大菱鲆[5]、虹鳟[6]、斑马鱼[7]、罗氏沼虾[8]等多种鱼虾类的精氨酸酶部分序列或全长序列。现有的研究资料表明,多数鱼类精氨酸酶的分布有组织特异性,一般在肝脏中精氨酸酶含量最高。在大菱鲆中30日龄前精氨酸酶基因表达水平一直很低,30日龄后随日龄增加其表达量逐步上升,至47日龄(标志其底栖生活的开始)达最高峰,随后表达量逐步下降,57日龄后再次上升至较低水平,比较精氨酸酶基因与甲状腺激素受体TRα的表达模式,Oriane等[5]发现当TRα表达水平降低时,精氨酸酶基因表达水平开始升高,证实在大菱鲆胚胎后发育期间精氨酸酶基因和甲状腺激素受体TRα的表达模式呈负相关。
本试验用设计的引物和提取的变态期牙鲆仔鱼总RNA进行RT-PCR,获得了一条284 bp DNA片段并测序。同源性分析显示目的基因与大菱鲆精氨酸酶基因的碱基序列及氨基酸序列的同源性较高,分别为91%和96%;与鲫鱼(Q4A4U9,Cyprinus carpio)、斑马鱼(Q6PH54,Danio rerio)、虹鳟(Q7T1U6,Oncorhynchus mykiss)碱基序列及推测的氨基酸序列同源性分别为78%/85%、84%/84%和80%/82%。与人(P78540,Homo sapiens human)的氨基酸序列同源性为72%。证明本试验所得到的目的基因片段是目前尚未见报道的牙鲆精氨酸酶基因核苷酸序列的一部分。本研究为重组牙鲆精氨酸酶基因的表达提供了可靠的基因材料,同时也为进一步研究该基因的结构和功能奠定了基础。
[1] 丁斐斐,李长红,陈炯,等.香鱼精氨酸酶II 基因的cDNA 克隆及其表达与鳗弧菌感染的相关性[J].生物技术通报,2016,32(2):109-115.
[2] 顾春燕,肖锋,钱铮,等.精氨酸酶-1在肝细胞癌中的低表达及其临床意义[J].中国癌症杂志,2014(6):438-445.
[3] 张佩,杨小艳,邸宏,等.玉米精氨酸酶基因过表达载体的构建与遗传转化分析[J].玉米科学,2012(6):66-71.
[4] 高晶晶,王旭辉,段唐海,等.精氨酸酶1在食管癌患者外周血的表达及其临床意义[J].免疫学杂志,2012(11):58-61.
[5] Oriane M,Marilyne D,Gerard T,et al.Molecular cloning and developmental expression patterns of thyroid hormone receptors and T3 target genes in the turbot(Scophtalmus maximus)during post-embryonic development[J].Gen.Comp.Endocrinol,2004,135:345-357.
[6] Wright P A,Campbell A,Morgan R L,et al.Dogmas and controversies in the handling of nitrogenous wastes:Expression of arginase TypeⅠ and Ⅱ genes in rainbow trout:influence of fasting on liver enzyme activity and mRNA levels in juveniles[J].The Journal of Experimental Biology,2004,207:2033-2042.
[7] Song H D,Sun X J,Deng M,et al.Hematopoietic gene expression profile in zebrafish kidney marrow[J].Proc.Natl.Acad.Sci.USA,2004(101):16240-16245.
[8] 田荣,许婷,潘晓艺,等.罗氏沼虾幼体精氨酸激酶基因的cDNA克隆及其在双顺反子病毒感染后的表达特征[J].水生生物学报,2016(5):908-913.
(责任编辑 崔建勋)
Cloning and sequencing of gene fragments of Paralichthys olivaceus arginase
LIU Hua1,SHI Ting-ting1,SHI Zhi-yi2
(1.R&D Center of Nanning Fresh-life Bio-technology Co.,Ltd,Nanning 530003,China;2.Research Center of Biotechnology,Shanghai Fisheries University,Shanghai 200090,China)
A pair of primers was designed and synthesized according to the conservative nucleotide sequence(GenBank)of Scophthalmus maximus,Cyprinus carpio,Oncorhynchus mykiss,Danio rerio,et al.Total RNA from Paralichthys olivaceus was extracted by using TRIzol Reagent.With the designed primers,a gene fragment was obtained using RT-PCR method.The PCR products were purified and then cloned into pUcm-T vector followed by sequencing.The results showed that a 284 bp gene fragment was obtained.Contrasted with other species,the gene sequence obtained in this experiment showed 96%,85%,84% and 82% homology with S.maximus,C.carpio,D.rerio,O.mykiss,respectively.Through sequencing the gene fragment of Arg by bioinformatics technique,we found that Arg had no SignalP,no O-glycosylation sites,and no transmembrane(TM)region,but had a character structure of arginase.There was a center of β-sheet in the N-terminal N0.38-44,57-64,71-76,89-94.And the N-terminal No.7-18,50-52,81-87,may be the Alpha regions of Arg protein.The N-terminal No.37-45,74-77,were the predominant epitopes.
Paralichthys olivaceus;arginase;molecular cloning;sequence analysis
S917
A
1004-874X(2016)12-0108-07
10.16768/j.issn.1004-874X.2016.12.019
2016-08-22
国家自然科学基金(30271017);国家高等教育博士项目科研基金(20040264001)
刘华(1977-),男,硕士,工程师,E-mail:luckyliuh@126.com
施志仪(1954-),男,博士,教授,E-mail:314481590@qq.com
刘华,施婷婷,施志仪.牙鲆精氨酸酶基因片段的克隆及序列分析[J].广东农业科学,2016,43(12):108-114.
猜你喜欢
黑龙江水产(2022年6期)2022-12-03
北方农业学报(2022年3期)2022-08-08
中国渔业质量与标准(2021年5期)2021-11-20
小天使·二年级语数英综合(2021年8期)2021-08-16
河北渔业(2019年11期)2019-12-11
中国水产(2019年3期)2019-03-25
中成药(2017年6期)2017-06-13
海洋科学(2017年10期)2017-02-09
中国组织化学与细胞化学杂志(2016年4期)2016-02-27
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
