
三江源区高寒草甸植物多样性的海拔分布格局
2016-01-27 07:11:38卢慧丛静刘晓王秀磊唐军李迪强张于光
草业学报 2015年7期
卢慧,丛静,刘晓,王秀磊,唐军,李迪强,张于光*
(1.中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态重点实验室,北京100091;2.中央民族大学
生命与环境科学学院,北京100081;3.中南大学资源加工与生物工程学院,湖南 长沙410083)
三江源区高寒草甸植物多样性的海拔分布格局
卢慧1,2,丛静1,3,刘晓1,王秀磊1,唐军1,李迪强1,张于光1*
(1.中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态重点实验室,北京100091;2.中央民族大学
生命与环境科学学院,北京100081;3.中南大学资源加工与生物工程学院,湖南 长沙410083)
摘要:三江源地区是我国受气候变化影响最大且最敏感的区域之一。研究高寒草甸植物多样性海拔分布格局及其与环境的关系,能为预测气候变化对植物多样性的影响和响应提供科学依据。本研究以青海三江源地区的6个样地/海拔梯度共78个样方的高寒草甸样地为研究对象,了解高寒草甸植物多样性和群落结构,分析海拔分布格局及其与环境的关系,旨在以空间替代时间的实验系统来揭示植物对气候变化的响应。统计分析发现,6个高寒草甸样地共有植物21科、51属、74种,且在中海拔和高海拔地区,高寒草甸以藏嵩草、高山嵩草、矮嵩草等莎草科植物占优势,低海拔地区高寒草甸则以针茅、早熟禾、垂穗披碱草等禾本科植物为优势种群。方差分析表明,随着海拔的降低,高寒草甸群落的物种多样性和物种丰富度均呈“单峰”分布格局,而均匀度指数逐渐升高。DCA分析发现,高寒草甸植物群落沿着海拔梯度呈现有规律的变化,具有从高寒草甸向高寒草原化草甸的逐渐过渡的特征,海拔梯度明显的影响了植物群落结构和多样性。此外,CCA结果表明,土壤pH值是影响高寒草甸植物群落分布格局的主要因素,土壤含水量、土壤温度、土壤有机碳、碱解氮含量与全钾含量是影响植物群落分布格局的关键因子。综上所述,我们推测气候变化极有可能影响三江源地区高寒草甸植物群落结构与分布格局。
关键词:三江源区;高寒草甸;植物多样性;海拔梯度;气候变化
DOI:10.11686/cyxb2014347
Lu H, Cong J, Liu X, Wang X L, Tang J, Li D Q, Zhang Y G. Plant diversity patterns along altitudinal gradients in alpine meadows in the Three River Headwater Region, China. Acta Prataculturae Sinica, 2015, 24(7): 197-204.
卢慧,丛静,刘晓,王秀磊,唐军,李迪强,张于光. 三江源区高寒草甸植物多样性的海拔分布格局. 草业学报, 2015, 24(7): 197-204.
http://cyxb.lzu.edu.cn
收稿日期:2014-08-21;改回日期:2014-12-10
基金项目:国家自然科学基金项目(No.31370145)和中央级公益性科研院所基本科研业务费专项(CAFRIFEEP201410)资助。
作者简介:卢慧(1985-),女,壮族,湖南长沙人,在读博士。E-mail: susanluhui@163.com
通讯作者*Corresponding author. E-mail: yugzhang@sina.com.cn
Abstract:The Three River Headwater region of Qinghai province, also known as the Qinghai-Tibetan Plateau is one of the regions in China likely to be most significantly affected by climate change. Hence, understanding the relationship between plant community distribution patterns and environment factors could afford scientific evidence for predicting the effect of climate change on plant diversity. In this research, six sites at a range of elevations and including 78 alpine meadow plots in the Three River Headwater region were surveyed to study the relationship between plant community structure and the environment. We aimed to explore the potential plant responses to climate change using a strategy of space-for-time substitution. At the six sites a total of 74 plant species belonging to 21 families and 51 genera were identified. Cyperaceous plants such as Kobresia schoenoides, Kobresia pygmaea and Kobresia humilis were the dominant species in the middle-high-elevation area, while plants of the Poaceae such as Stipa capillata, Poa annua and Elymus nutans covered the most of low elevation area. Species diversity index and richness index of the alpine meadow sites showed a unimodal pattern, and species evenness showed no significant differences along with the altitudinal gradient across the six sites. Detrended correspondence analysis indicated that species composition and diversity of plant communities changed continuously with decreasing elevation from alpine meadow to alpine steppe meadow. In addition, canonical correspondence analysis showed that soil pH was the main factor affect species diversity patterns in alpine meadow. Meanwhile, soil moisture, temperature, organic carbon content, available nitrogen content and total potassium content were key factors affect the distribution patterns of plant communities. On the basis of these observations, we predict that climate change is very likely to change plant community structure in the Three River Headwater Region.
Plant diversity patterns along altitudinal gradients in alpine meadows in the Three River Headwater Region, China
LU Hui1,2, CONG Jing1,3, LIU Xiao1, WANG Xiu-Lei1, TANG Jun1, LI Di-Qiang1, ZHANG Yu-Guang1*
1.InstituteofForestEcology,EnvironmentandProtection,ChineseAcademyofForestry,KeyLaboratoryofForestEcologyandEnvironmentStateForestryAdministration,Beijing100091,China; 2.CollegeofLifeandEnvironmentSciences,MinzuUniversityofChina,Beijing100081,China; 3.SchoolofMineralProcessingandBioengineering,CentralSouthUniversity,Changsha410083,China
Key words: the Three River Headwater Region; alpine meadow; plant diversity; elevation gradient; climate change
生物多样性沿环境梯度的变化格局一直是生物多样性研究的一个重要议题,海拔梯度的变化综合了温度、湿度和光照等多种环境因子,因此,在一定程度上能够显著地影响植物种群的空间分布格局[1]。大量研究表明,植物多样性随着海拔梯度的变化常呈现出以下几种分布规律[2-7]:随海拔梯度先升高后降低(单峰分布格局)、先降低后升高、随海拔升高而降低、随海拔升高而递增和无规律性。近年来,许多学者对森林、草原、湿地环境下的植物群落进行了分布格局与环境解释的研究[8-11],对于高寒草甸的研究多集中于基本结构、外貌特征、生产力以及物质循环、能量流动等方面[12-15],而对高寒草甸群落分布格局的影响因子的研究报道相对较少。
研究物种多样性随海拔梯度的变化在一定程度上可以用于探讨物种多样性对气候变化的响应与适应[16]。以往的研究多集中在控制实验上进行且周期较短的野外观测研究,而沿着环境梯度样带和野外实验相结合的研究比较缺乏。由海拔变化引起的自然温度梯度作为气候变化的替代实验系统,包括了温度、湿度和光照等多种环境因子的梯度效应,以空间代替时间,是评价气候变化对陆地生态系统可能影响的经济实用的方法,近年来被广泛应用,并取得了颇有说服力的结果[17]。
三江源地区是黄河、长江和澜沧江等的发源地,是生物多样性资源宝库,对我国江河中下游地区以及东南亚国家的生存与发展起着重要的生态安全保障作用[18]。高寒草甸是三江源最主要的植被类型,在各植被类型中分布面积最大,为171662 km2[19]。在全球气候变化背景下,尤其是在一些人为干扰因素的影响下,高寒草甸已呈现明显的退化态势。本研究以高寒草甸植被为研究对象,运用统计学方法分析样地调查得到的物种数量、盖度、物种高度和环境因子数据,并用除趋势对应分析(detrended correspondence analysis,DCA)和典范对应分析(canonical correspondence analysis,CCA)两种排序方法分析了高寒草甸群落物种多样性的海拔分布格局及其影响因子,试图为预测气候变化对植物多样性影响和响应提供科学依据。
1材料与方法
1.1 研究区概况
三江源地区位于青藏高原腹地,青海省的南部。该地区地形复杂,自然环境类型多样,生物多样性丰富。该地区属于典型高原大陆性气候,年均气温在-5.6~3.8℃之间,极端最低气温为-48℃,极端最高气温为28℃,年均降水量在 262.2~772.8 mm之间,年蒸发量在 730~1700 mm之间[19]。高寒草甸是由耐寒的多年生草本植物为主或有高寒灌丛参与形成、以矮草草群占优势的一种草地植被类型,是在高原亚寒带与寒带寒冷半干旱的气候条件下发育形成[20],广泛分布于海拔3800~5200 m的山顶、滩地、山地阳坡。由于植物种类随环境的变化而发生变化,可以将其分为高寒嵩草草甸、高寒杂类草草甸、高寒草原化草甸和沼泽化草甸[19]。
1.2 样地选择和取样方法
选择青海省三江源地区的玛多县、兴海县等地的典型高寒草甸植被作为研究区域(97°40′22″-100°05′27″ E,34°08′16″-35°56′06″ N,表1),于2012年8月进行采样,沿海拔3220~4790 m以每相差约300 m设立一块样地,共设立6块样地。每块样地的坡度、坡向、人为干扰情况尽可能一致。6块样地沿海拔梯度分别命名为SJY-4790、SJY-4480、SJY-4140、SJY-3880、SJY-3490、SJY-3220,其中,样地SJY-3880附近有采矿活动,受到的人为干扰相对较多,如踩踏、采药等。样地基本情况见表1。为了在考虑空间异质性的同时降低实际地形的取样难度,采样时对巢式取样[21]的方法进行了简化,进行了“L”形取样。即在每块样地设立1个200 m×200 m的网格,以网格内任意一个角为起点,在水平和垂直方向上距离分别为5,10,20,50,100和200 m处设置1 m×1 m的样方,每块样地内设立13个1 m×1 m样方,6块样地共78个样方。在每个1 m×1 m的样方内采用对角线取样法采集土壤样品,取样深度为0~10 cm,均匀混合后低温保存带回实验室用于土壤理化性质分析。同时记录采样地点的经纬度、地形和土壤温度[22](10 cm深度)等。
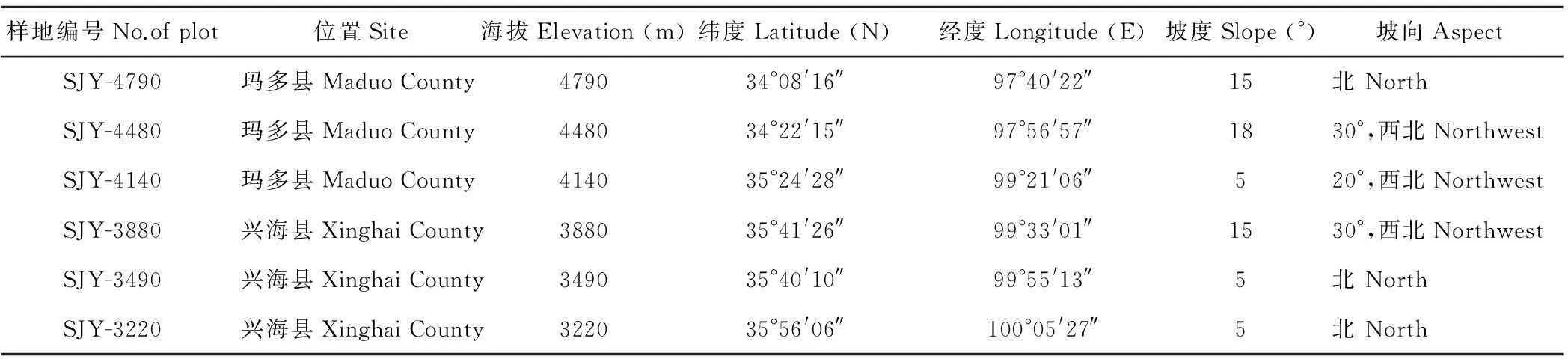
表1 6个样地基本情况
1.3 物种多样性和土壤理化性质测定
在每个样方中,调查植物种类、多度、高度、盖度等指标,计算重要值。物种多样性的测定采用物种丰富度指数(S)、Shannon-Wiener多样性指数(H′)和Pielou指数(J)表征[23]。其中,H′=-∑PilnPi;J=H′/lnS。式中,Pi为i物种的相对重要值,即(相对高度+相对多度)/2,S为群落中的总物种数。
土壤有机碳(SOC)用重铬酸钾氧化-分光光度法测定[24],pH值用酸度计法测定[25],土壤含水量、全磷(TP)、全钾(TK)、全硫(TS)采用常规方法测定[25],碱解氮(AN)用碱解-扩散法测定[24],速效磷(AP)和速效钾(AK)用盐酸和硫酸溶液浸提,等离子发射光谱法测定[24]。
1.4 数据分析
植物群落中物种多样性指数均采用Excel 2010进行统计计算;采用SPSS 18.0软件,对植物群落各指标进行单因素方差分析(one-way ANOVA),比较各样方的差异;并将物种重要值转换成78×74矩阵,用除趋势对应分析(DCA)进行排序;并将该矩阵与78×10的环境因子矩阵结合,用典范对应分析(CCA)研究群落分布格局与环境因子的关系。DCA和CCA均使用生物统计学软件R软件中的Vegan软件包进行统计分析。采用SigmaPlot 12.5软件作图。
2结果与分析
2.1 植物群落分布特征
在6个样地中共有植物21科、51属、74种(表2),其中,以样地SJY-4140的物种数最多,有14科30属37 种;样地SJY-3880物种数最少,有8科16属20种。样地SJY-4790海拔最高,为4790 m,位于巴颜喀拉山山顶,为藏嵩草草甸,主要优势种有藏嵩草、高山嵩草,植物共有12科31属30种,其中莎草科与禾本科植物重要值分别为54.3和24.2。样地SJY-4480也位于巴颜喀拉山,海拔为4480 m,是以藏嵩草为建群种的高寒草甸,该样地共有植物11科27属36种,该样地莎草科植物重要值为43.2,禾本科植物重要值为17.8。样地SJY-4140为高山嵩草草甸,海拔4140 m,主要优势种是高山嵩草和矮嵩草,植物共有14科30属37种,其中莎草科植物重要值为31.1,禾本科植物重要值为18.1。样地SJY-3880海拔为3880 m,为高山嵩草草甸,主要优势种为高山嵩草和针茅,该样地植物共有8科16属20种,其中莎草科植物重要值为41.1,禾本科植物重要值为26.7。样地SJY-3490海拔为3490 m,为早熟禾-针茅草原化草甸,优势种为早熟禾、针茅等禾本科植物,该样地植物共有10科20属25种,其中莎草科植物重要值为17,禾本科植物重要值为31.1。样地SJY-3220海拔为3220 m,为垂穗披碱草草甸,其优势种为垂穗披碱草和针茅,该样地植物共有12科21属23种,其中莎草科植物重要值为5.1,禾本科植物重要值为45.8。
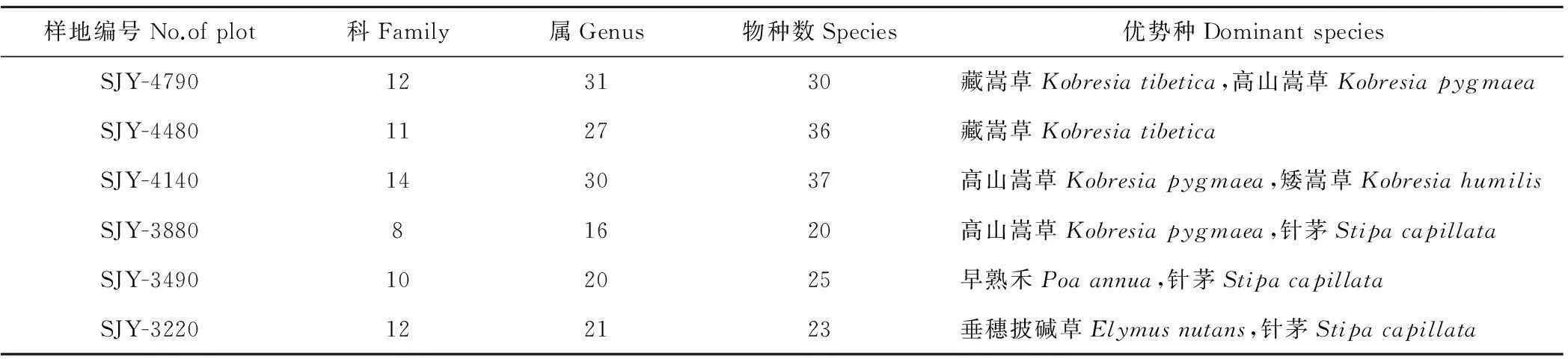
表2 植物群落调查情况
2.2 植物群落物种多样性
根据样方调查得到的植物种类、株数、高度等数据,计算Shannon-Wiener多样性指数、Pielou均匀度指数和物种丰富度指数,从表3可以看出,6个样地植物群落多样性差异明显。Shannon-Wiener多样性指数从大到小依次为SJY-4480>SJY-4140>SJY-3490>SJY-3220>SJY-4790>SJY-3880,物种丰富度从大到小依次为SJY-4480>SJY-4140>SJY-4790>SJY-3490>SJY-3220>SJY-3880,二者均在群落SJY-3880出现最低值,这可能与该样地较多的人为干扰有关(踩踏、采药等)。随着海拔的降低,除SJY-3880以外的5个样地的Shannon-Wiener指数与物种丰富度指数均呈现先增加后减少的趋势,中海拔的多样性和丰富度相对较高;Pielou指数在样地SJY-3880明显低于其他5个样地,而其他5个样地则沿海拔降低而逐渐升高。
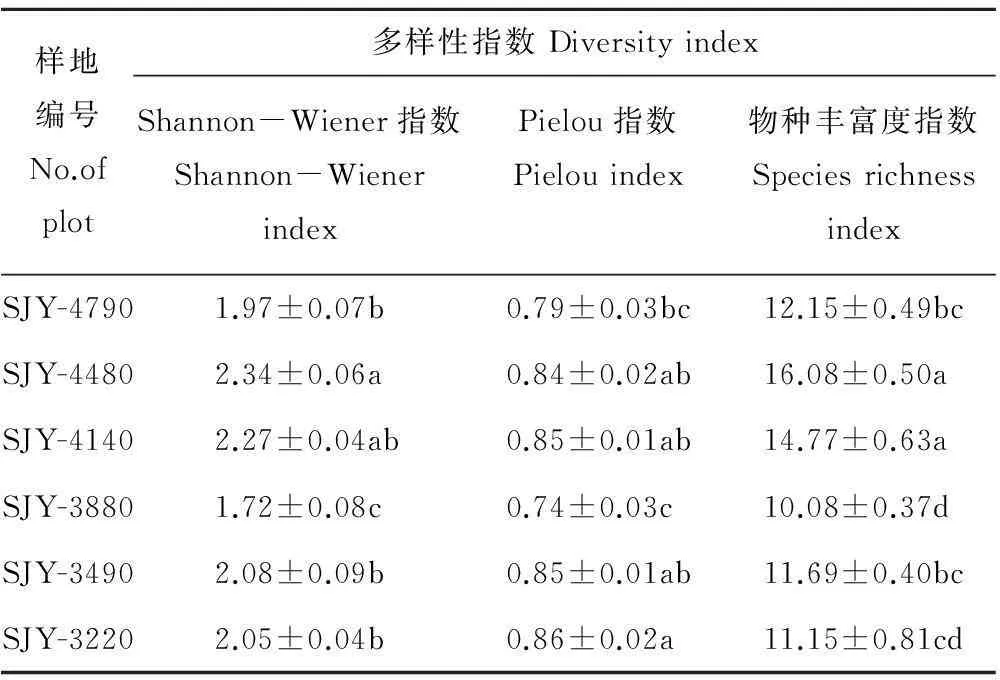
表3 6个样地植物群落多样性
注:数字为均值±标准误,同列中的不同字母表示在P<0.05水平上差异显著。
Note: Values are mean±SE. Different letters in the same column denoted significant differences atP<0.05.2.3DCA分析
通过对高寒草甸样地78个样方进行除趋势对应分析(DCA)得到二维排序图(图1)。从图中可以看出,DCA第1轴解释了植物群落13.6%的变异,第2轴解释了植物群落6.6%的变异。在排序图中,各群落类型都有着自己的分布区域,虽然各自分成小团块分布,但相邻样地的群落在图中的位置仍有少量的交错。从群落类型来看,SJY-4790与SJY-4480海拔较高,湿度相对较高,而土壤温度较低,二者均以藏嵩草为建群种,群落种类组成相似,二者在排序图上所处位置相近,均处于排序图的左边;SJY-4140与SJY-3880处于中间海拔,均为高山嵩草草甸,群落组成差异相对较小,二者在排序图上处于中间偏下的位置;SJY-3490与SJY-3220处于相对较低海拔梯度, 土壤温度较高, 含水量较低,禾本科植物种类增多,而莎草科植物种类减少,群落组成处于草甸群落向草原群落过渡地带,在排序图上处于右边位置。从群落SJY-4790、SJY-4480、SJY-4140、SJY-3880、SJY-3490、SJY-3220过渡的连续性来看,它们之间的相互关系是较为紧密的,随着海拔的逐渐降低,呈现出从高寒草甸向高寒草原化草甸的过渡,样方中莎草科植物种所占比重逐渐减小,禾本科植物与杂类草所占比重逐渐增加,草甸逐渐草原化。以上结果表明,各不同群落类型在排序图上的位置基本反映出群落过渡的连续性,表明植物多样性沿着海拔梯度进行更替,海拔梯度明显地影响了植物群落结构和多样性。
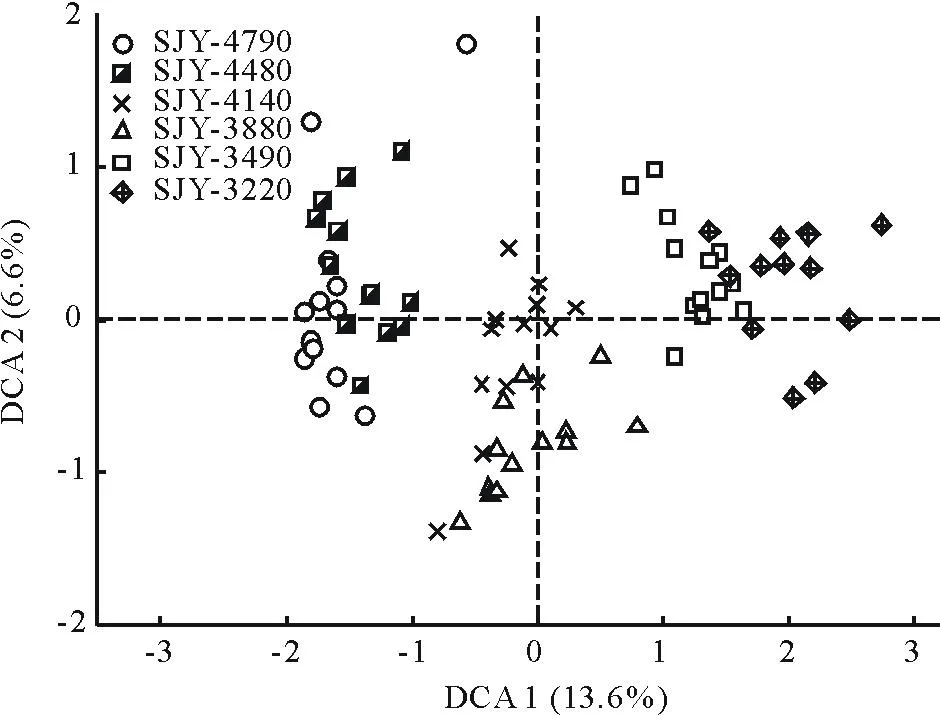
图1 6个样地植物群落DCA二维排序图Fig.1 DCA biplot of plant communities in 6 sample sites
2.4 植物群落与环境因子的相关性分析
CCA分析能在一定程度上解释植物群落的分布格局与环境因子的关系。本研究选取10个与植物群落结构显著相关的环境因子,构成以78×10的环境因子矩阵,与78×74的样地物种矩阵进行CCA排序分析,得到图2。图2是样方的二维排序图,图中箭头表示环境因子,箭头长短表示植物群落的分布与环境因子的相关性。CCA双序图中,箭头所处象限表示环境因子与排序轴之间相关性的正负。第1排序轴解释了样本中39.7%的变异,第2排序轴解释了16.3%的变异。第1轴和第2轴与环境因子显著相关,其特征值分别为0.616和0.253。
从图2可以看出,与第1排序轴相关性最高的是土壤pH值,呈极显著正相关关系,相关系数为0.957(P<0.01),其次为土壤温度,相关系数为0.676(P<0.01);与第1排序轴负相关最高的是土壤含水量,相关系数为0.927(P<0.01),其次为土壤有机碳和碱解氮,相关系数为0.872和0.871(P<0.01),说明第1排序轴主要反映了植物群落分布格局在土壤pH值与土壤含水量梯度上的变化;与第2排序轴相关性最高的为土壤温度,其相关系数为0.675(P<0.01),说明第2排序轴主要反映了植物群落分布格局在土壤温度梯度上的变化。而从箭头长短来看,箭头较长的是pH值,说明pH值可能是影响植物群落分布最重要的因素,同时,土壤温度、土壤含水量、土壤有机碳、碱解氮和全钾也具有较长的箭头,说明这些因素对高寒草甸群落结构分布格局也具有重要影响。而土壤全磷、全硫、速效磷和速效钾的箭头长度较短,与CCA两排序轴的相关性相对较小,说明其与群落分布的关系也较小。
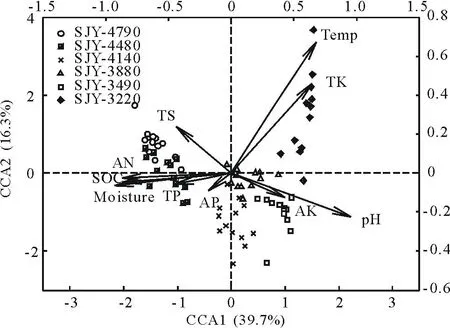
图2 植物群落与环境因子典范对应分析Fig.2 Canonical correspondence analysis between plant community and environmental factors pH:土壤pH值 Soil pH;Temp:土壤温度 Soil temperature; Moisture:土壤水分 Soil moisture; SOC:土壤有机碳Soil organic carbon;TP:全磷 Total phosphorus;TK:全钾 Total potassium;TS:全硫 Total sulfur;AN:碱解氮 Available nitrogen;AP:速效磷 Rapid available phosphorus;AK:速效钾 Rapid available potassium.
3讨论
近年来,由于受到全球气候变暖,尤其是超载过牧等因素的影响,三江源地区草地生态系统出现恶化,退化草场面积达到可利用草场面积的26%~46%[26],水土流失严重,土壤碳大量损失,已有研究表明,青藏高原因草地退化而每年损失的土壤碳储量约为全国的土壤碳储量的6.5%[27]。高寒草甸是该地区最主要的植被类型,因此,研究高寒草甸植物群落特征与多样性分布格局及其与环境的关系,对理解高寒草甸生态系统的变化过程和机理以及保护和改善生态环境具有重要意义。本研究通过对6个海拔梯度共78个样方的高寒草甸植物群落进行调查,在较大尺度上对植物物种多样性的海拔分布格局及其影响因子进行了研究,6块样地取样数目一致且同一样地样品数量较多,极大地减少了空间异质性造成的误差,得到的研究结果更具可靠性和代表性。
6个样地的高寒草甸均由耐寒的多年生草本植物为主,由于影响植物分布的因素较多,草本植物多样性沿海拔变化的关系比较复杂。本研究结果表明,除受到人为干扰明显的SJY-3880以外的5个样地,沿着海拔的降低植物物种多样性和物种丰富度呈现先增加后减少的变化趋势,中海拔梯度植物多样性较高,即呈单峰分布格局,这与王长庭等[28]在青海省果洛州玛沁县海拔3840~4435 m的高寒草甸群落植物多样性的研究结果一致。王长庭等[28]认为高寒草甸群落物种多样性、丰富度和均匀度在中间海拔梯度最高;三者与地上生物量之间的趋势模拟均呈负二次函数关系,说明在群落生产力水平与物种丰富度中等时,物种多样性最高。段敏杰等[18]以藏北地区那曲县日囤布山的高寒草地为试验样地,对海拔4523~4673 m共4个海拔梯度的植物多样性分布格局进行了研究,认为该区域植物群落物种丰富度、多样性指数和均匀度指数均沿海拔梯度呈现“单峰”分布格局。而在本研究中,均匀度指数在样地SJY-3880明显低于其他5个样地,而其他5个样地均匀度指数变化趋势沿海拔降低而逐渐升高。
土壤 pH值是土壤重要的基本性质之一,与土壤的肥力状况、微生物活动及作物生长有密切关系。pH可以通过影响土壤营养元素的有效性而影响植物生长[29]。赵振勇等[30]认为土壤含盐量与物种多样性指标无显著关系,土壤盐分含量变化影响着植物群落物种组成,随土壤盐分含量增加,群落中盐生植物种类逐渐占据优势。本研究呈现出类似的趋势,随着海拔的降低,土壤pH值逐渐升高,禾本科牧草和杂类草的数量增加,这些植物比高海拔的嵩草属牧草更加耐旱、抗盐碱,对环境适应性更强。CCA分析结果表明,土壤pH值可能是影响高寒草甸植物群落结构最重要的因素。
本研究区域处于海拔3220~4790 m之间,研究尺度相对较大,DCA研究结果表明海拔对植物群落分布的影响较为明显。随着海拔的变化,土壤温度与土壤含水量随之发生改变,这在一定程度上代表了气候的变化,这种水热条件的变化所引起的物种选择、资源竞争、生境的变化是影响物种多样性的重要因素[31]。徐满厚和薛娴[32]对青藏高原高寒草甸进行模拟增温实验得知,增温对植被产生正效应,植被高度和地上生物量随着温度升高有增加趋势。张庆等[33]对内蒙古草原短花针茅(Stipabreviflora)群落的DCCA分析结果反映出群落结构格局主要受到热量、水分梯度变化的影响。因此,水热条件也是影响植物群落变化的主要因素,田海芬等[34]在研究大青山山地植物分布格局的影响因素时也得出了类似结果。此外,土壤有机碳、氮素和磷素、钾素等是土壤主要的养分指标,在本研究中,土壤含水量、土壤有机碳、碱解氮和全钾含量对植物群落分布影响较大,是影响高寒草甸植物群落分布格局的关键因子,这与丛静等[35]对神农架保护区森林植被影响因素的研究结果基本一致。同时我们可以看出,由海拔变化引起的自然温度梯度作为气候变化的替代实验系统,通过海拔梯度对植物多样性及其分布格局的影响来预测全球气候变化对植物多样性影响和响应,这种方法是合理可行的。气候变化极有可能对三江源地区高寒草甸植物群落结构与分布格局造成影响。
References:
[1]Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227.
[2]He J S, Chen W L. A review of gradient changes in species diversity of land plant communities. Acta Ecologica Sinica, 1997, 17(1): 91-99.
[3]Tang Z Y, Fang J Y. A review on the elevational patterns of plant species diversity. Biodiversity Science, 2004, 12(1): 20-28.
[4]He Y H, Yan M, Zhang Q D. Altitudinal pattern of plant species diversity in the Wulu Mountain Nature Reserve, Shanxi, China. Acta Ecologica Sinica, 2013, 33(8): 2452-2462.
[5]Kong X H, Li Z J. Species diversity and altitudinal gradient patterns of evergreen broad-leaved forest in Meihuashan National Natural Reserve, Fujian Province. Plant Diversity and Resources, 2012, 34(2): 179-186.
[6]Marini L, Bona E, Kunin W E. Exploring anthropogenic and natural processes shaping fern species richness along elevational gradients. Journal of Biogeography, 2010, 38(1): 78-88.
[7]Ohlemuller R, Wilson J B. Vascular plant species richness along latitudinal and altitudinal gradients: a contribution from New Zealand temperate rainforests. Ecology Letters, 2000, 3(4): 262-266.
[8]Li G L, Cong J, Lu H,etal. Numerical classification and ordination of forest communities in habitat of Sichuan Snub-nosed Monkey in Hubei Shennongjia National Nature Reserve. Acta Ecologica Sinica, 2012, 32(23): 7501-7511.
[9]Liu H W, Cheng X Q, Kang F F. Changes of understory plant community inPinustabuliformisplantation and associated environmental explanations. Chinese Journal of Ecology, 2014, 33(2): 290-295.
[10]Chen B R, Li H S, Zhu Y X,etal. The spatial pattern and environmental interpretation of the plant community of Hulunber grassland. Acta Ecologica Sinica, 2010, 30(5): 1265-1271.
[11]Ren B. Analysis on the Typical Phytocoenoses and its Principle Factors of Distribution Patterns in East Dongting Lake Wetland[D]. Changsha: Hunan Agricultural University, 2012.
[12]Hu L, Wang C T, Wang G X,etal.Changes in the activities of soil enzymes and microbial community structure at different degradation successional stages of alpine meadows in the headwater region of Three Rivers, China. Acta Prataculturae Sinica, 2014, 23(3): 8-19.
[13]Liu Y S, Fan J W, Li Y Z,etal.Plant community productivity and diversity on alpine meadow steppe in the Three River Headwater Region, Qinghai Province under different denudation levels. Acta Prataculturae Sinica, 2014, 23(3): 1-7.
[14]Liu Y J, Shi G X, Mao L,etal. Direct and indirect influences of 8 yr of nitrogen and phosphorus fertilization on Glomeromycota in an alpine meadow ecosystem. New Phytologist, 2012, 194(2): 523-535.
[15]Rui Y C, Wang S P, Xu Z H,etal. Warming and grazing affect soil labile carbon and nitrogen pools differently in an alpine meadow of the Qinghai-Tibet Plateau in China. Journal of Soils and Sediments, 2011, 11(6): 903-914.
[16]Liu M C, Li D Q, Wen Y M,etal. The ecological function analysis and evaluation of ecosystem in Sanjiangyuan region. Acta Scientiae Circumstantiae, 2005, 25(9): 1280-1286.
[17]Li D Q, Li J W. Biodiversity: Scientific Investigation Report on Sanjiangyuan Natural Reserve[M]. Beijing: China Science and Technology Press, 2002.
[18]Duan M J, Gao Q Z, Guo Y Q,etal.Species diversity distribution pattern of alpine grassland communities along an altitudinal gradient in the Northern Tibet. Pratacultural Science, 2011, 28(10): 1845-1850.
[19]Bryant J A, Lamanna C, Morlon H,etal. Microbes on mountainsides: contrasting elevational patterns of bacterial and plant diversity. Proceedings of The National Academy of Sciences, 2008, 105(Supplement 1): 11505-11511.
[20]Wang K, Hong F Z, Zong J Y. Grassland resources and their sustainable utility in the “Three-River Headwaters” Region. Acta Agrestia Sinica, 2005,13(z1): 28-47.
[21]Fang J Y, Wang X P, Shen Z H,etal. Methods and protocols for plant community inventory. Biodiversity Science, 2009, 17(6): 533-548.
[22]Zhang Y G, Cong J, Lu H,etal. Community structure and elevational diversity patterns of soilAcidobacteria. Journal of Environmental Sciences, 2014, 26(8): 1717-1724.
[23]Ma K P, Liu Y M. Measurement of biotic community diversity I α diversity (Part 2). Biodiversity Science, 1994, 2(4): 231-239.
[24]Institute of Soil Science, Chinese Academy of Sciences. Soil Physical and Chemical Property Analysis[M]. Shanghai: Shanghai Science and Technology Press, 1983.
[25]Bao S D. Soil Agricultural Chemistry Analysis[M]. Beijing: China Agriculture Press, 2003.
[26]Xiao T, Shao Q Q, Sun W Y,etal. Grassland degradation characteristics of typical alpine meadow slopes in the Three-River Source Region of Qinghai province. Acta Agrestia Sinica, 2013, 21(3): 452-459.
[27]Wang G X, Li Y S, Wang Y B,etal. Effects of permafrost thawing on vegetation and soil carbon pool losses on the Qinghai-Tibet Plateau, China. Geoderma, 2008, 143(1): 143-152.
[28]Wang C T, Wang Q J, Long R J. Changes in plant species diversity and productivity along an elevation gradient in an alpine meadow. Acta Phytoecologica Sinica, 2004, 28(2): 240-245.
[29]Zhao Y K, Zhang W S, Wang Y N,etal. Research progress in physiology and molecular biology of plant responses to high pH. Chinese Journal of Eco-Agriculture, 2008, 16(3): 783-787.
[30]Zhao Z Y, Wang R H, Yin C H,etal. Species diversity and spatial heterogeneity of plant communities in piedmont plain of south slope of Tianshan Mountains. Acta Botanica Boreali-Occidentalia Sinica, 2007, 27(4): 784-790.
[31]Wang J B, Zhang D G, Cao G M,etal. Regional characteristics of the alpine meadow degradation succession on the Qinghai-Tibetan Plateau. Acta Prataculturae Sinica, 2013, 22(2): 1-10.
[32]Xu M H, Xue X. A research on summer vegetation characteristics & short-time responses to experimental warming of alpine meadow in the Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 2013, 33(7): 2071-2083.
[33]Zhang Q, Niu J M, Buyantuyev A,etal. Ecological analysis and classification ofStipabrevifloracommunities in the Inner Mongolia region: the role of environmental factors. Acta Prataculturae Sinica, 2012, 21(1): 83-92.
[34]Tian H F, Liu H M, Wang W,etal. The distribution patterns of biodiversity and environmental interpretation in Daqingshan Mountain. Journal of Arid Land Resources and Environment, 2014, 28(8): 172-177.
[35]Cong J, Ying H Q, Lu H,etal. Species diversity and environmental interpretation of typical vegetation types in the Shennongjia Natural Reserve. Scientia Silvae Sinicae, 2013, 49(5): 30-35.
参考文献:
[2]贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征. 生态学报,1997,17(1): 91-99.
[3]唐志尧, 方精云. 植物物种多样性的垂直分布格局. 生物多样性, 2004,12(1): 20-28.
[4]何艳华, 闫明, 张钦弟. 五鹿山国家级自然保护区物种多样性海拔格局. 生态学报, 2013, 33(8): 2452-2462.
[5]孔祥海, 李振基. 福建梅花山常绿阔叶林植物物种多样性及其海拔梯度格局. 植物分类与资源学报, 2012, 34(2): 179-186.
[8]李广良, 丛静, 卢慧, 等. 神农架川金丝猴栖息地森林群落的数量分类与排序. 生态学报, 2012, 32(23): 7501-7511.
[9]刘宏文, 程小琴, 康峰峰. 油松人工林林下植物群落变化及其环境解释. 生态学杂志, 2014, 33(2): 290-295.
[10]陈宝瑞, 李海山, 朱玉霞, 等. 呼伦贝尔草原植物群落空间格局及其环境解释. 生态学报, 2010, 30(5): 1265-1271.
[11]任勃. 东洞庭湖湿地典型植物群落及其格局主因子分析[M]. 长沙: 湖南农业大学, 2012.
[12]胡雷, 王长庭, 王根绪, 等. 三江源区不同退化演替阶段高寒草甸土壤酶活性和微生物群落结构的变化. 草业学报, 2014, 23(3): 8-19.
[13]刘艳书, 樊江文, 李愈哲, 等. 三江源地区不同剥蚀退化高寒草甸群落生物量与多样性特征. 草业学报, 2014, 23(3): 1-7.
[16]刘敏超, 李迪强, 温琰茂, 等.三江源地区生态系统生态功能分析及其价值评估. 环境科学学报, 2005, 25(9): 1280-1286.
[17]李迪强, 李建文. 三江源生物多样性: 三江源自然保护区科学考察报告[M]. 北京: 中国科学技术出版社, 2002.
[18]段敏杰, 高清竹, 郭亚奇, 等.藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局. 草业科学, 2011, 28(10): 1845-1850.
[20]王堃, 洪绂曾, 宗锦耀.“三江源”地区草地资源现状及持续利用途径. 草地学报, 2005, 13(增刊): 28-47.
[21]方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容, 方法和技术规范. 生物多样性, 2009, 17(6): 533-548.
[23]马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ.α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239.
[24]中国科学院南京土壤研究所. 土壤理化分析[M].上海: 上海科学技术出版社, 1983.
[25]鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2003.
[26]肖桐, 邵全琴, 孙文义, 等. 三江源高寒草甸典型坡面草地退化特征综合分析. 草地学报, 2013, 21(3): 452-459.
[28]王长庭, 王启基, 龙瑞军. 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究. 植物生态学报, 2004, 28(2): 240-245.
[29]赵彦坤, 张文胜, 王幼宁, 等. 高pH对植物生长发育的影响及其分子生物学研究进展. 中国生态农业学报, 2008, 16(3): 783-787.
[30]赵振勇, 王让会, 尹传华, 等. 天山南麓山前平原植物群落物种多样性及空间分异研究. 西北植物学报, 2007, 27(4): 784-790.
[31]王建兵, 张德罡, 曹广民, 等.青藏高原高寒草甸退化演替的分区特征. 草业学报, 2013, 22(2): 1-10.
[32]徐满厚, 薛娴. 青藏高原高寒草甸夏季植被特征及对模拟增温的短期响应. 生态学报, 2013, 33(7): 2071-2083.
[33]张庆, 牛建明, Buyantuyev A, 等. 内蒙古短花针茅群落数量分类及环境解释. 草业学报, 2012, 21(1): 83-92.
[34]田海芬, 刘华民, 王炜, 等. 大青山山地植物区系及生物多样性研究. 干旱区资源与环境, 2014, 28(8): 172-177.
[35]丛静, 尹华群, 卢慧, 等. 神农架保护区典型植被的物种多样性和环境解释. 林业科学, 2013, 49(5): 30-35.
猜你喜欢
英语世界(2023年10期)2023-11-17 09:19:12
资源节约与环保(2022年12期)2022-02-02 02:39:16
中华环境(2021年9期)2021-10-14 07:51:16
生态文明新时代(2018年4期)2018-04-25 11:57:12
生态文明新时代(2018年1期)2018-03-21 05:16:48
农业与技术(2016年22期)2017-03-07 02:01:16
中国高新技术企业(2016年32期)2016-12-27 11:42:43
湖北农业科学(2016年18期)2016-12-08 17:07:13
太阳能(2015年7期)2015-04-12 06:49:50
