
南方根结线虫侵染对不同抗性番茄砧木渗调物质及羟脯氨酸含量的比较
2016-01-27 07:11贾双双徐坤
草业学报 2015年7期
关键词:番茄
贾双双,徐坤
(1.安徽科技学院生命科学学院,安徽 凤阳 233100;2.山东农业大学园艺科学与工程学院,山东 泰安 271018)
南方根结线虫侵染对不同抗性番茄砧木渗调物质及羟脯氨酸含量的比较
贾双双1,2,徐坤2*
(1.安徽科技学院生命科学学院,安徽 凤阳 233100;2.山东农业大学园艺科学与工程学院,山东 泰安 271018)
摘要:为探讨番茄砧木对南方根结线虫的抗性生理机制,以高感砧木Ls-89与高抗砧木坂砧2号为试材,采用盆栽人工接种线虫二龄幼虫法,研究了南方根结线虫侵染对不同抗性番茄砧木幼苗渗透调节物质及细胞壁羟脯氨酸含量的影响。研究表明,未接种南方根结线虫的番茄砧木幼苗,其根系与叶片可溶性糖、可溶性蛋白、脯氨酸等渗透调节物质与细胞壁羟脯氨酸含量变化并未因品种抗性不同而有显著差异(P>0.05);接种南方根结线虫后,渗透调节物质含量与细胞壁羟脯氨酸含量变化规律基本一致,均呈周期性升高,且坂砧2号升幅显著高于Ls-89(P<0.05)。据此认为,可溶性糖、可溶性蛋白、脯氨酸、细胞壁羟脯氨酸等物质的含量变化均与番茄砧木幼苗对南方根结线虫的抗性密切相关,可作为评判植株抗性的综合指标。
关键词:番茄;砧木幼苗;南方根结线虫;渗透调节物质;细胞壁羟脯氨酸
DOI:10.11686/cyxb2014333
Jia S S, Xu K. The effects ofMeloidogyneincognitainfection on osmolyte and hydroxyproline levels in tomato rootstock seedlings with different resistance. Acta Prataculturae Sinica, 2015, 24(7): 123-130.
贾双双, 徐坤. 南方根结线虫侵染对不同抗性番茄砧木渗调物质及羟脯氨酸含量的比较. 草业学报, 2015, 24(7): 123-130.
http://cyxb.lzu.edu.cn
收稿日期:2014-08-20;改回日期:2014-12-10
基金项目:安徽省教育厅项目(KJ2013B078),安徽科技学院引进人才项目(ZRC2013351),山东省现代农业产业技术创新体系(SDAIT-02-022-05)和山东省农业重大应用技术创新项目(31648)资助。
作者简介:贾双双(1984-),女,河北赵县人,讲师,博士。E-mail:jiashuang84@163.com
通讯作者*Corresponding author. E-mail:xukun@sdau.edu.cn
Abstract:In order to clarify the physiological resistance mechanism of tomato rootstock to Meloidogyne incognita, both a highly susceptible rootstock variety Ls-89 and a highly resistant rootstock variety Banzhen No.2 were planted in pots and inoculated with second instar larvae to study the effects of M. incognita infection on osmolyte levels and cell wall hydroxyproline contents of these differentially resistant tomato rootstock varieties. Without M. incognita inoculation, there was no significant difference (P>0.05) between seedlings of the two varieties in concentrations of osmoregulation substances, including soluble sugar, soluble protein, proline and cell wall hydroxyproline. After inoculation, the concentrations of osmoregulation substances and cell wall hydroxyproline contents were increased in roots and leaves in seedlings of the two varieties, with the responses in Banzhen No.2 being significantly higher (P<0.05) than those of Ls-89. These results indicate that changes in soluble sugar, soluble protein, proline and cell wall hydroxyproline concentrations are closely associated with the resistance of tomato rootstock seedlings to M. incognita, and could be used as general indices of plant resistance elevation.
The effects ofMeloidogyneincognitainfection on osmolyte and hydroxyproline levels in tomato rootstock seedlings with different resistance
JIA Shuang-Shuang1,2, XU Kun2*
1.CollegeofLifeScience,AnhuiScienceandTechnologyUniversity,Fengyang233100,China; 2.CollegeofHorticultureScienceandEngineering,ShandongAgriculturalUniversity,Tai’an271018,China
Key words: tomato (Lycopersiconesculentum); rootstock seedlings;Meloidogyneincognita; osmoregulation substance; cell wall hydroxyproline
植物寄生线虫病是农业生产中的重要病害之一,已在世界范围内引发巨大经济损失[1-2],因此,前人对植物线虫病的防治方法[3-4]、抗性机制[5]等进行了较多的研究。前人研究表明,植物寄生线虫病不仅显著降低根系吸收能力[6],影响根系活力[7-8],还显著降低叶片叶绿素含量[9-10]。谈家金等[11]研究表明,松材线虫侵染使黑松(Pinusthunbergii)幼苗茎部可溶性糖与可溶性蛋白含量降低,并认为可溶性蛋白含量可用于病害的早期诊断;而郝玉金等[12]研究发现,山定子(Malusbaccata)接种南京毛刺线虫后,根尖游离脯氨酸与还原糖含量呈现不同程度的增加,而总糖含量却降低。
富含羟脯氨酸糖蛋白(hydroxyproline-rich glycoprotein,HRGP)是高等植物细胞壁中特有的一种结构蛋白,广泛存在于植物中。近年来研究发现,它与植物的抗病性存在密切关系[13]。其羟脯氨酸(Hyp)含量高是这类糖蛋白的特点之一,且细胞壁中HRGP含量与Hyp含量呈正相关[14],因此,对羟脯氨酸(Hyp)含量的测定,在某种程度上代表了HRGP的含量。翟衡等[15]研究发现,不同抗性苹果砧木接种南京毛刺线虫后,根尖组织细胞壁中Hyp含量均有不同程度的增加,且抗性砧木的增加幅度均大于感病砧木,并认为Hyp含量的增加可能是苹果砧木抵御南京毛刺线虫的一种防御机制。赵小钒和弭忠祥[16]也发现,不同抗性大豆(Glycinemax)品种感染灰斑病后,抗病品种叶片细胞壁中Hyp无论是积累含量还是维持时间均显著高于感病品种,并认为Hyp含量的积累与大豆对灰斑病的抗性密切相关。
番茄(Lycopersiconesculentum)是我国设施栽培面积最大的蔬菜之一,因其复种指数高、连作重茬栽培较为普遍,致使番茄根结线虫病频发[17-18]。Lopez-Perez等[19]、Rivard和Louws[20]指出,采用抗性品种和利用抗性砧木嫁接栽培将成为最具潜力的防控作物根结线虫病的有效方法,但不同抗性番茄砧木对南方根结线虫的反应不同,而且关于番茄砧木抗南方根结线虫的生理机制研究甚少。泰勒和萨塞[21]指出,在适宜的环境条件下,根结线虫的生活周期为30 d左右。由于植物苗期是对逆境胁迫最为敏感的时期,在植物生长早期进行抗性筛选鉴定较为容易[22],因此,本试验在前文[23]研究基础上,选用2个抗南方根结线虫水平显著不同的番茄砧木为试材,研究南方根结线虫一个侵染周期内,幼苗根系与叶片中可溶性糖、可溶性蛋白、脯氨酸等渗透调节物质及细胞壁Hyp的变化动态,旨在探讨渗透调节物质及细胞壁Hyp积累与番茄砧木幼苗抗南方根结线虫的关系,为进一步研究番茄砧木抗线虫生理机制及抗性砧木的筛选鉴定提供理论参考。
1材料与方法
1.1 试验材料
试验于2011-2012年在山东农业大学园艺实验站进行,供试番茄砧木为抗南方根结线虫水平显著不同的2个品种Ls-89(高感)、坂砧2号(高抗)[23]。种子浸种催芽后,播于50孔穴盘内,播后置日光温室培养。幼苗长至3~4片真叶时,移植于直径25 cm、高18 cm的塑料盆中,每盆2株,每品种栽植180盆,每30盆为一个处理小区,其中3个小区接种南方根结线虫二龄幼虫,另3个小区为对照,随机区组排列,重复3次。盆内栽培基质(砂∶土=1∶2)经160℃高温杀菌2 h。
1.2 接种取样
移栽番茄砧木幼苗缓苗后,在幼苗根围打孔(5孔/盆),将南方根结线虫二龄幼虫按5000条/盆注入,对照注入等量清水,然后用土掩埋。
分别于接种南方根结线虫后第0,1,3,7,14,21,28,35 天取样,每次每小区随机取3株,将根系及叶片用流水洗净,擦去表面水分后,投入液氮中,带回实验室进行相应生理指标的测定,同时调查计算根结指数和病情指数。
1.3 测定指标和方法
1.3.1抗病指标的测定参照毛爱军等[24]制定的标准调查番茄幼苗发病情况,并计算病情指数(disease index,DI)。0级:植株根部无根瘤;1级:植株根部形成1~2个根瘤;2级:植株根部形成3~10个根瘤;3级:植株根部形成11~30个根瘤;4级:植株根部形成31~100个根瘤;5级:植株根部形成根瘤数超过100个。
DI=∑(各病级植株数×该级数)/(调查总株数×最重病级数值)×100
采用Boiteux和Charchar[25]的方法计算根结指数(gall index,GI),GI=单株根结数/单株根鲜重(g)。
1.3.2可溶性糖含量的测定按照赵世杰等[26]的方法进行,采用蒽酮比色法进行提取和测定,于630 nm下比色。
1.3.3可溶性蛋白含量的测定按照赵世杰等[26]的方法进行,采用考马斯亮蓝法进行提取和测定,于595 nm下比色。
1.3.4脯氨酸(Pro)含量的测定按照赵世杰等[26]的方法进行,采用磺基水杨酸—酸性茚三酮法进行提取和测定,于520 nm下比色。
1.3.5细胞壁羟脯氨酸(Hyp)含量的测定按照冯洁等[27]的方法制备细胞壁干样,按照Shailasree等[28]的方法测定细胞壁Hyp含量。取细胞壁干样20 mg,悬浮于5 mL 6 mol/L HCl中,110℃酸解18 h。取酸解液2 mL,用1 mol/L KOH溶液调pH值至7.0,加1 mL pH 8.7 0.8 mol/L硼酸缓冲液和2 mL 0.2 mol/L氯胺T,室温下反应25 min,加2 mL 3.6 mol/L Na2S2O3溶液终止反应,加2 g KCl饱和后用3 mL甲苯萃取,弃甲苯层,于沸水浴中加热30 min,冷却后加4 mL甲苯再次萃取,取甲苯层2 mL与1 mL对甲氨基苯甲醛(DAMB)试剂室温显色20 min,560 nm处测定OD值,计算Hyp含量。
1.4 数据处理
采用Microsoft Excel 2003与SPSS 17.0软件对数据进行处理和统计分析。
2结果与分析
2.1 接种南方根结线虫对不同抗性番茄砧木幼苗发病情况的影响
从表1可以看出,接种南方根结线虫后7 d,高感品种Ls-89幼苗根系已形成少量根结,说明线虫已侵入根系,其后,随侵染时间的延长,Ls-89的根结指数(GI)与病情指数(DI)均迅速升高,至接种后35 d时,其GI与DI分别高达30.45和96.67;而高抗品种坂砧2号直至接种28 d后,植株根系才出现根结,且其GI与DI分别仅为0.15和9.67,接种后35 d,分别为0.44和26.67。
2.2 南方根结线虫侵染对番茄砧木幼苗渗透调节物质含量的影响
2.2.1幼苗根系与叶片可溶性糖含量的变化Ls-89与坂砧2号未接种根结线虫的对照幼苗根系与叶片可溶性糖含量逐渐升高(图1),且坂砧2号显著高于Ls-89(P<0.05)。接种后,Ls-89与坂砧2号根系与叶片可溶性糖含量均随着侵染发生显著性变化,至接种后7 d,根系可溶性糖含量均达峰值,两砧木分别较其对照增加65.60%和106.01%(P<0.01),之后迅速降低,至28 d时降至最低,随后于35 d因二次侵染又有所升高,在整个侵染进程中,坂砧2号始终显著高于Ls-89(P<0.05)。接种后,Ls-89与坂砧2号叶片可溶性糖含量均在21 d达到峰值,分别较其对照增加46.80%和83.44%(P<0.01)。

表1 接种南方根结线虫后供试番茄砧木幼苗发病情况比较
注:同列不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)。
Note:Different small letters in the same column mean significant difference at 0.05 level, different capital letters mean extremely significant difference at 0.01 level.
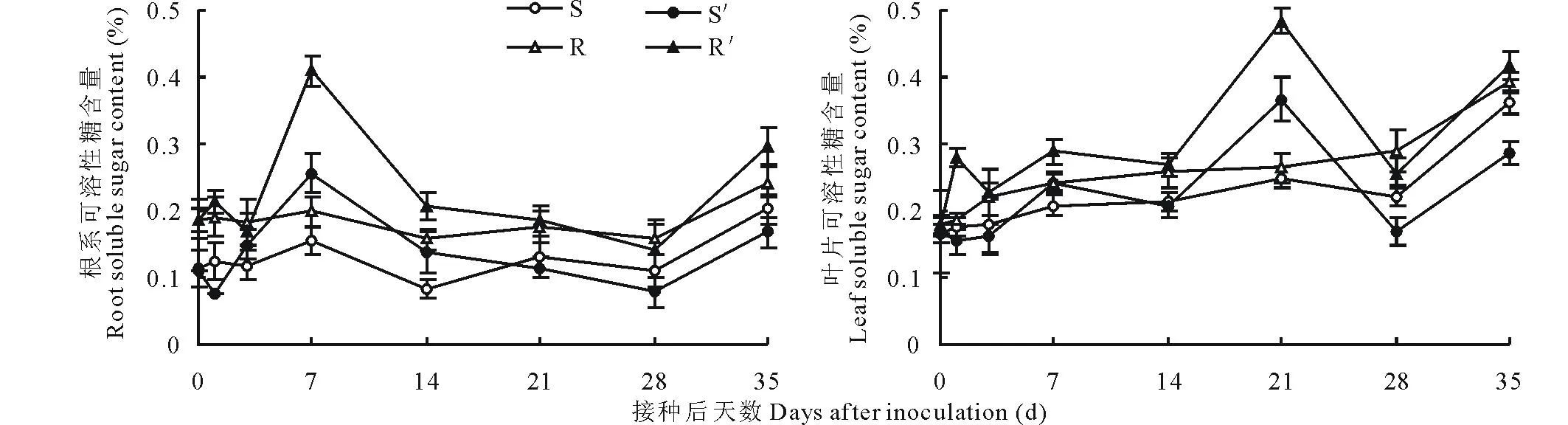
图1 南方根结线虫对番茄砧木幼苗根系和叶片可溶性糖含量的影响Fig.1 Effect of M. incognita on soluble sugar content in roots and leaves of tomato rootstock seedlings S′与R′分别表示接种南方根结线虫的Ls-89与坂砧2号,S与R分别表示其对照,下同。S′ and R′ represent Ls-89 and Banzhen No.2 inoculated with M. incognita, respectively, S and R represent that of the control, respectively. The same below.
2.2.2幼苗根系与叶片可溶性蛋白含量的变化图2可以看出,Ls-89与坂砧2号未接种根结线虫的对照幼苗根系与叶片可溶性蛋白含量差异不显著(P>0.05)。接种后7 d,Ls-89与坂砧2号根系可溶性蛋白含量均达峰值,分别较其对照增加172.88%和198.75%(P<0.01);而叶片则在接种后14 d达到峰值,分别较其对照增加48.67%和74.68%(P<0.05)。
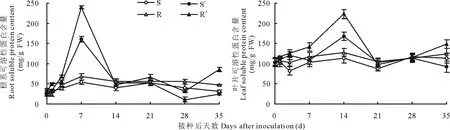
图2 南方根结线虫对番茄砧木幼苗根系和叶片可溶性蛋白含量的影响Fig.2 Effect of M. incognita on soluble protein content in roots and leaves of tomato rootstock seedlings
2.2.3幼苗根系与叶片脯氨酸含量的变化图3所示,未接种根结线虫的Ls-89与坂砧2号对照幼苗根系与叶片Pro含量变化不大,且差异不显著(P>0.05)。接种3 d,Ls-89与坂砧2号根系Pro含量均达峰值,分别较其对照增加166.30%和242.86%(P<0.01),而叶片均在接种后14 d达到峰值,分别较其对照增加73.90%和108.68%(P<0.01)。
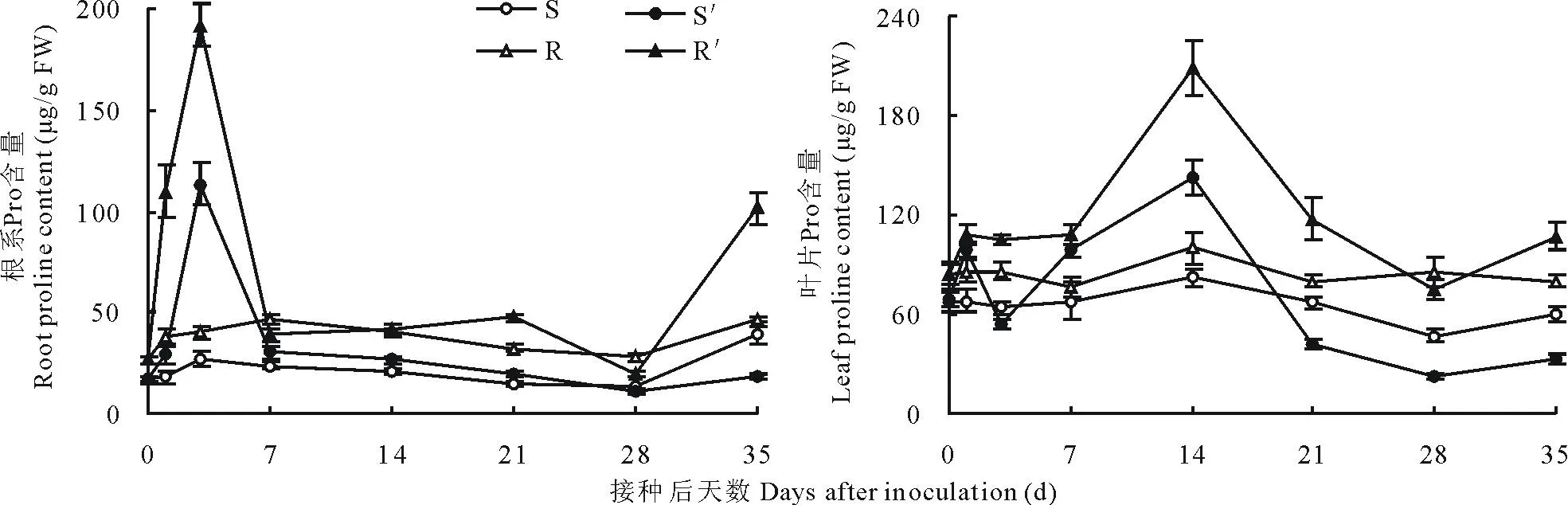
图3 南方根结线虫对番茄砧木幼苗根系和叶片脯氨酸含量的影响Fig.3 Effect of M. incognita on proline content in roots and leaves of tomato rootstock seedlings
2.2.4幼苗根系与叶片细胞壁Hyp含量的变化图4所示,未接种根结线虫的Ls-89与坂砧2号幼苗根系与叶片细胞壁Hyp含量变化基本稳定,且差异不显著(P>0.05),但根系显著高于叶片(P<0.05)。接种后,Ls-89和坂砧2号根系与叶片细胞壁Hyp含量发生剧烈变化,至7 d时,两砧木根系细胞壁Hyp含量达峰值,分别较其对照增加91.79%和133.33%(P<0.01),之后迅速降低,至接种后28 d降至低谷,随后因二次侵染又有所升高。接种后至7 d,Ls-89叶片细胞壁Hyp含量逐渐升高,7~21 d间又稍有降低,至28 d达峰值,较其对照增加96.44%(P<0.01);而坂砧2号于侵染后14 d达峰值,较其对照增加109.62%(P<0.01)。整个侵染过程中,坂砧2号根系与叶片细胞壁Hyp含量均高于Ls-89。
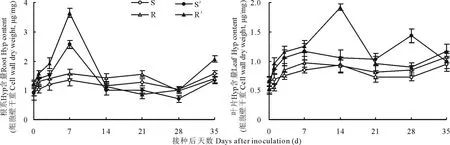
图4 南方根结线虫对番茄砧木幼苗根系和叶片羟脯氨酸含量的影响Fig.4 Effect of M. incognita on Hyp content in roots and leaves of tomato rootstock seedlings
3讨论
3.1 南方根结线虫对不同抗性番茄砧木幼苗渗透调节物质的影响
可溶性糖、可溶性蛋白与脯氨酸等均是植物体内适应逆境胁迫的重要渗透调节物质,与植物抗性密切相关[10,29-32]。刘大伟等[33]研究表明,不同抗性大豆品种接种大豆胞囊线虫后,抗病品种可溶性糖含量低于感病品种,而可溶性蛋白含量则相反。Lewis和McClure[34]研究发现,不同抗性棉花(Gossypiumhirsutum)品种遭受南方根结线虫侵染后,根系脯氨酸含量增加,但感病品种增幅显著高于抗病品种。而本研究结果表明,接种南方根结线虫后,高感砧木Ls-89与高抗砧木坂砧2号根系与叶片可溶性糖、可溶性蛋白与脯氨酸等渗透调节物质含量升高,这与刘业霞等[35]的研究结果一致,与谈家金等[11]相反,且坂砧2号升幅显著高于Ls-89(P<0.05),这与Lewis和McClure[34]的研究结果相反。这可能是因为抗病砧木可以通过调节体内渗透调节物质含量来调节代谢平衡,延缓根结线虫侵害,而感病砧木随着线虫侵染,根结指数与病情指数持续增长,植株生长受抑制,可用渗透调节物质减少,无法正常调节代谢平衡。
3.2 南方根结线虫对不同抗性番茄砧木幼苗细胞壁Hyp含量的影响
细胞壁富含羟脯氨酸糖蛋白,与植物的抗病性存在密切关系,目前已在多种植物病害中证实[13]。Esquerre-Tugaye等[36]研究认为,甜瓜(Cucumismelo)幼苗感染炭疽病后,茎部细胞壁Hyp含量增加;Sujeeth等[37]研究发现,不同抗性珍珠粟(Pennisetumglaucum)幼苗感染霜霉病后,抗病品种胚芽鞘细胞壁Hyp含量显著高于感病品种。本研究结果也证实,不同抗性番茄砧木幼苗接种南方根结线虫后,根系与叶片细胞壁Hyp含量升高,且高抗砧木坂砧2号显著高于高感砧木Ls-89(P<0.05)。
4结论
不同抗性番茄砧木幼苗渗透调节物质差异不大,而遭受南方根结线虫侵染后,可溶性糖、可溶性蛋白与脯氨酸等渗透调节物质含量迅速升高,细胞壁也随之积累较多的羟脯氨酸,且均以高抗砧木坂砧2号显著高于高感砧木Ls-89(P<0.05)。说明高抗砧木可通过较高含量的可溶性糖、可溶性蛋白及脯氨酸等渗透调节物质,细胞壁也通过积累较多的羟脯氨酸来调节代谢平衡,延缓根结线虫侵害。渗透调节物质与细胞壁羟脯氨酸含量的增加是番茄砧木幼苗抗根结线虫的机制之一。
References:
[1]Williamson V M, Hussey R S. Nematode pathogenesis and resistance in plants. The Plant Cell, 1996, 8(10): 1735-1745.
[2]Sasser J N. Plant Pasasitic Nematodes: the Farmer’s Hidden Enemy[M]. Beijing: Popular Science Press, 1992.
[3]Giannakou I O, Anastasiadis I. Evaluation of chemical strategies as alternatives to methyl bromide for the control of root-knot nematodes in greenhouse cultivated crops. Crop Protection, 2005, 24: 499-506.
[4]Cao Z P, Yu Y L, Chen G K,etal. Impact of soil fumigation practices on soil nematodes and microbial biomass. Pedosphere, 2004, 14(3): 387-393.
[5]Tan J J, Ye J R. Research advance in pathogenic mechanism of pine wood nematode disease. Journal of Huazhong Agricultural University, 2003, 22(6): 613-617.
[6]Zhai H, Guan X Q, Zhao C Z,etal. Screening of Chinese grape species resistant toM.incognita. Acta Horticulturae Sinica, 2000, 27(1): 27-31.
[7]Li W C, Dong H, Wang X F. Effects of root-knot nematode on growth, quality and yield of cucumber in greenhouse. Journal of Shandong Agricultural University, 2006, 37(1): 35-38.
[8]Bin S Y, Yao S M, Lin J T,etal. Some physiological changes of peanut plants infected withMeloidogynearenaria. Journal of Huazhong Agricultural University, 1999, 18(2): 121-124.
[9]Guo Y Y, Xu K, Wang X F. Effects ofMeloidogyneincognitaonon the growth and development of ginger. Acta Agriculturae Boreali-occidentalis Sinica, 2005, 14(5): 155-158.
[10]Chen Y H, Ye J R, Wei C J,etal. Effects of pine wood nematode (PWN) infection on water regime and metabolism of related to hosts. Acta Phytopathologica Sinica, 2005, 35(3): 201-207.
[11]Tan J J, Ye J R, Hao D J,etal. Effects of pine wood nematode,Bursaphelenchusxylophilus, on some physio-biochemical indexes of Japanese black pine,Pinusthunbergii. Acta Phytopathologica Sinica, 2011, 41(1):44-48.
[12]Hao Y J, Zhai H, Wang S H. Change of physiochemical substances inMalusbaccatainfected withTrichodorusnanjingensis. Acta Phytopathologica Sinica, 1999, 29(1): 82-85.
[13]Song F M, Zheng Z, Ge Q X. Hydroxyproline-rich glycoproteins in plant-pathogen interactions in the accumulation, the role and regulation. Plant Physiology Communications, 1992, 28(2): 141-145.
[14]Lamport D T, Miller D H. Hydroxyproline arabinosides in the plant kingdom. Plant Physiology, 1971, 48: 454-456.
[15]Zhai H, Hao Y J, Wang S H. Preliminary study on the resistance mechanism of apple rootstock to a stubby root nematode. Acta Phytophylacica Sinica, 1999, 26(3): 208-212.
[16]Zhao X F, Mi Z X. Study on content of hydroxyproline in cell wall in relation to resistance toCrecosporasojinahare. Soybean Science, 2000, 19(2): 146-149.
[17]Peng D L, Tang W H. Advance of resistance gene Mi to root-knot nematodes in tomato. Journal of Shenyang Agricultural University, 2001, 32(3): 220-223.
[18]Sasser J N, Eisenback J D, Carter C C,etal. The internationalMeloidogyneproject-its goals and accomplishments. Annual Review Phytopathology, 1983, 21: 271-288.
[19]Lopez-Perez J A, Strange M L, Kaloshian I,etal. Differential response ofMigene-resistant tomato rootstocks to root-knot nematodes (Meloidogyneincognita). Crop Protection, 2006, 25: 382-388.
[20]Rivard C L, Louws F J. Tomato grafting for disease resistance and increased productivity. Sustainable Agriculture Research &Education, 2011, 12: 1-8.
[21]Taylor A L, Sasser J N. Plant Root-knot Nematode[M]. Yang B J, translation. Beijing: Science Press, 1983.
[22]Zhang L Q, Zhang F Y, Hasi A. Research progress on alfalfa salt tolerance. Acta Prataculturae Sinica, 2012, 21(6): 296-305.
[23]Jia S S, Gao R G, Xu K. Screening and evaluation of tomato rootstocks for resistance toMeloidogyneincognita. Scientia Agricultura Sinica, 2009, 42(12): 4301-4307.
[24]Mao A J, Chai M, Yu S C,etal. Technique of identification for resistance to root-knot nematode in tomato and its application. Acta Agriculture Boreali-occidentalis Sinica, 2005, 14(4):140-144.
[25]Boiteux L S, Charchar J M. Genetic resistance to root-knot nematode (Meloidogynejavanica) in eggplant (Solanummelongena). Plant Breeding, 1996, (3): 198-200.
[26]Zhao S J, Liu H S, Dong X C. Techniques of Plant Physiological Experiment[M]. Beijing: China Agricultural Science and Technology Press, 1998.
[27]Feng J, Chen Q Y, Shi L Y. Accumulation of HRGP in cell wall of cotton treated withFusariumoxysporumF. SP.Vasinfectumrelatedto the resistance toFusariumwilt. Acta Phytopathologica Sinica, 1995, 25(2): 133-138.
[28]Shailasree S, Ramachandra Kini K, Deepak S,etal. Accumulation of hydroxyproline-rich glycoproteins in pearl millet seedlings in response toSclerosporagraminicolainfection. Plant Science, 2004, 167: 1227-1234.
[29]Cao F X, Wang M, Teng T,etal. Influence of pine wood nematode on plasma-membrane permeability and soluble sugar content ofPinuselliottiileaves. Journal of Central South Forestry University, 2006, 26(6): 169-171.
[30]Zhang H S, Zhao G Q, Li M F,etal. Physiological responses ofPennisetumlongissimumvar.intermediumseedlings to PEG, low temperature and salt stress treatments. Acta Prataculturae Sinica, 2014, 23(2): 180-188.
[31]Liu A R, Zhang Y B, Zhong Z H,etal. Effects of salt stress on the growth and osmotica accumulation ofColeusblumei. Acta Prataculturae Sinica, 2013, 22(2): 211-218.
[32]Shi L R, Niu Y L, Li M Z. Physiological response ofSonchusbrachyotusto salt stress. Acta Prataculturae Sinica, 2010, 19(6): 272-275.
[33]Liu D W, Duan Y X, Chen L J,etal. Physiological mechanism of huipizhiheidou resistant to Race 3 of soybean cyst nematode. Soybean Science, 2010, 29(3): 471-473.
[34]Lewis S A, McClure M A. Free amino acids in roots of infected cotton seedlings resistant and susceptible toMeloidogyneincognita. Journal of Nematology, 1975, 7(1): 10-15.
[35]Liu Y X, Jiang F, Zhang N,etal. Relationship between osmoregulation and Bacterial wilt resistance of grafted pepper. Acta Horticulturae Sinica, 2011, 38(5): 903-910.
[36]Esquerre-Tugaye M T, Lafitte C, Mazau D,etal. Cell surfaces in plant-microorganism interactions. Plant Physiology, 1979, 64: 320-326.
[37]Sujeeth N, Deepak S, Shailasree S,etal. Hydroxyproline-rich glycoproteins accumulate in pearl millet after seed treatment with elicitors of defense responses againstSclerosporagraminicola. Physiological and Molecular Plant Pathology, 2010, 74: 230-237.
参考文献:
[2]Sasser J N. 植物寄生线虫——农业的隐蔽敌害[M]. 北京: 科学普及出版社, 1992.
[5]谈家金, 叶建仁. 松材线虫病致病机理的研究进展. 华中农业大学学报, 2003, 22(6): 613-617.
[6]翟衡, 管雪强, 赵春芝, 等. 中国葡萄抗南方根结线虫野生资源的筛选. 园艺学报, 2000, 27(1): 27-31.
[7]李文超, 董会, 王秀峰. 根结线虫对日光温室黄瓜生长、果实品质及产量的影响. 山东农业大学学报(自然科学版), 2006, 37(1): 35-38.
[8]宾淑英, 姚圣梅, 林进添, 等. 花生根结线虫对花生植株主要生理指标的影响. 华中农业大学学报, 1999, 18(2): 121-124.
[9]郭衍银, 徐坤, 王秀峰. 南方根结线虫对生姜生长发育的影响. 西北农业学报, 2005, 14(5): 155-158.
[10]陈玉惠, 叶建仁, 魏初奖, 等. 松材线虫侵染对马尾松、黑松水分及其相关代谢的影响. 植物病理学报, 2005, 35(3): 201-207.
[11]谈家金, 叶建仁, 郝德君, 等. 松材线虫对黑松一些生理生化指标的影响. 植物病理学报, 2011, 41(1): 44-48.
[12]郝玉金, 翟衡, 王寿华. 山定子感染南京毛刺线虫后几种生理生化物质的变化. 植物病理学报, 1999, 29(1): 82-85.
[13]宋凤鸣, 郑重, 葛起新. 富含羟脯氨酸糖蛋白在植物-病原物相互作用中的积累、作用及调控. 植物生理学通讯, 1992, 28(2): 141-145.
[15]翟衡, 郝玉金, 王寿华. 苹果砧木对南京毛刺线虫的抗性机制初探. 植物保护学报, 1999, 26(3): 208-212.
[16]赵小钒, 弭忠祥. 细胞壁羟脯氨酸的含量与大豆灰斑病抗性关系的研究. 大豆科学, 2000, 19(2): 146-149.
[17]彭德良, 唐文华. 番茄抗根结线虫Mi基因研究进展. 沈阳农业大学学报, 2001, 32(3): 220-223.
[21]泰勒 A L, 萨塞 J N. 植物根结线虫[M]. 杨宝君, 译. 北京: 科学出版社, 1983: 5-43.
[22]张立全, 张凤英, 哈斯阿古拉. 紫花苜蓿耐盐性研究进展. 草业学报, 2012, 21(6): 296-305.
[23]贾双双, 高荣广, 徐坤. 番茄砧木对南方根结线虫抗性鉴定. 中国农业科学, 2009, 42(12): 4301-4307.
[24]毛爱军, 柴敏, 于拴仓, 等. 番茄抗根结线虫接种鉴定技术及其应用. 西北农业学报, 2005, 14(4): 140-144.
[26]赵世杰, 刘华山, 董新纯. 植物生理学实验指导[M]. 北京: 中国农业科技出版社, 1998.
[27]冯洁, 陈其煐, 石磊岩. 枯萎菌诱导棉花细胞壁富含羟脯氨酸糖蛋白积累与枯萎病抗性间的关系. 植物病理学报, 1995, 25(2): 133-138.
[29]曹福祥, 王猛, 滕涛, 等. 松材线虫对湿地松叶片质膜和可溶性糖的影响. 中南林学院学报, 2006, 26(6): 169-171.
[30]张怀山, 赵桂琴, 栗孟飞, 等. 中型狼尾草幼苗对PEG、低温和盐胁迫的生理应答. 草业学报, 2014, 23(2): 180-188.
[31]刘爱荣, 张远兵, 钟泽华, 等. 盐胁迫对彩叶草生长和渗透调节物质积累的影响. 草业学报, 2013, 22(2): 211-218.
[32]时丽冉, 牛玉璐, 李明哲. 苣荬菜对盐胁迫的生理响应. 草业学报, 2010, 19(6): 272-275.
[33]刘大伟, 段玉玺, 陈立杰, 等. 灰皮支黑豆抗大豆胞囊线虫3号生理小种的生理机制. 大豆科学, 2010, 29(3): 471-473.
[35]刘业霞, 姜飞, 张宁, 等. 嫁接辣椒对青枯病的抗性及其与渗透调节物质的关系. 园艺学报, 2011, 38(5): 903-910.
猜你喜欢
小猕猴学习画刊(2022年12期)2022-02-06
今日农业(2021年21期)2022-01-12
今日农业(2021年21期)2022-01-12
今日农业(2020年23期)2020-12-15
今日农业(2020年16期)2020-09-25
红领巾·探索(2020年2期)2020-05-19
启蒙(3-7岁)(2018年2期)2018-03-15
海峡姐妹(2017年6期)2017-06-24
小学生作文(低年级适用)(2017年12期)2017-02-06
创新作文(5-6年级)(2015年9期)2015-10-21
