
红鳍笛鲷(Lutjanus erythopterus)卵巢发育的组织学研究*
2016-01-15 03:52张玉红刘楚吾
海洋与湖沼 2016年1期
刘 皓 张玉红 罗 杰 刘楚吾 刘 丽
(广东海洋大学水产学院 南海水产经济动物增养殖广东普通高校重点实验室 湛江 524025)
红鳍笛鲷(Lutjanus erythopterus Bloch 1790)隶属硬骨鱼纲、辐鳍亚纲、鲈形目、笛鲷科、笛鲷属, 为暖水性近底层鱼类, 生存温度范围为 2—34°C, 最适生长温度范围为 25—30°C, 适应盐度为 5—40, 最适盐度为20—30, 分布范围广, 在我国主要产于南海和东海南部, 为南海重要经济鱼类之一。红鳍笛鲷个体较大, 生长速度快, 肉质丰厚, 含丰富的蛋白质和脂肪, 经济效益显著。近年来, 对红鳍笛鲷的研究主要集中在繁殖技术、养殖技术、生化研究和遗传学方面的研究(陈武各, 2002; 郑乐云等, 2004; 彭银辉等,2008), 有关红鳍笛鲷性腺发育的研究很少, 仅见杨春玲等(2010)用组织学方法对 1.5—2.5龄红笛鲷(Lutjanus sanguineus)的性腺发育进行了初步研究。本文对红鳍笛鲷性腺发育进行了组织学研究, 以期掌握红鳍笛鲷性腺发育的过程及周期变化、各发育时期生殖细胞的细胞学特点, 了解其繁殖性能, 进一步开发和保护好这一优良种质资源, 并且进一步丰富鱼类繁殖生物学的内容。
1 材料与方法
养殖红鳍笛鲷性腺发育材料由湛江海康流沙湾鱼排提供。1龄以内, 每1—2个月取材一次; 1龄以上, 每3—6个月取材一次, 每次取材5—10尾。测量体长、体重后, 解剖取出性腺, 用Bouin’s液固定, 参照芮菊生(1980)《组织切片技术》的方法, 在室温下作连续石蜡切片, 切片厚度 6—8µm, 苏木精-伊红染色, Leica DM 6000 B摄影生物显微镜观察、摄影。
2 结果
2.1 卵细胞生长期时相的划分及特点
根据红鳍笛鲷雌性生殖细胞在各个发育时期的体积大小、内含物累积变化等细胞生物学特征, 以及其形态特征, 参照刘筠(1993)的分期标准, 将其生长发育过程(包括退化)分为6个时相。
Ⅰ时相(卵原细胞) 由原始生殖细胞分化而来的卵原细胞, 细胞直径13µm左右, 圆形或椭圆形,以分散(Ⅰ期早期卵巢)或者细胞团(Ⅰ期晚期卵巢)的方式分布在蓄卵板的边缘, 呈微嗜酸性反应; 细胞核位于细胞中央, 体积较大, 直径8µm左右, 核质比为61%。核切面中央部位一般可见大的核仁1—2个(图1, 图 10)。
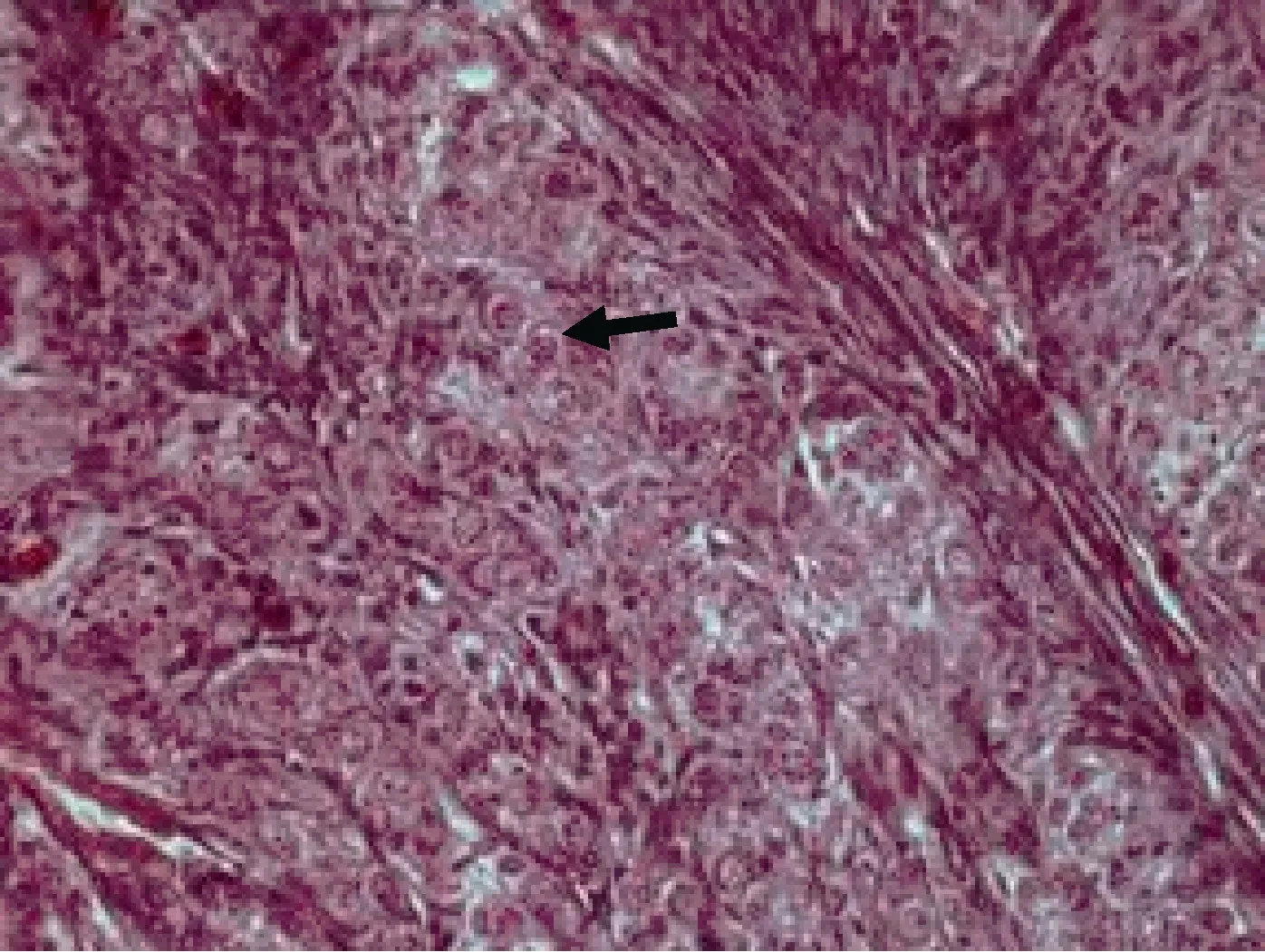
图 1 Ⅰ时相(卵原细胞期)(×400)Fig.1 The oogonium in phase Ⅰ(black arrow) (×400)
Ⅱ时相卵母细胞 由卵原细胞分化而来, 属于初级卵母细胞的小生长期。细胞呈圆形或椭圆形,多角, 细胞质呈嗜碱性反应。细胞膜外包围一层滤泡细胞构成的滤泡膜。卵母细胞平均直径 36µm 左右,胞核直径平均20µm左右, 核质比为55%。细胞核中的核仁数增多, 多靠近核膜内侧分布。也有的Ⅱ时相中期卵母细胞中似乎具有旁核, 但不是普遍现象。
根据Ⅱ时相卵母细胞和核的直径, 核仁的数量、大小及核仁位置的变化、染色的深浅等特征, 可将其发育分为早、中、晚3个阶段。(1) 早期: 刚从卵原细胞分化而来, 体积较小, 染色深, 核仁数目少(1到多个)而体积大, 多位于核的中央, 一般都有一个体积特别大的核仁(图2); (2) 中期: 体积较大, 染色深,核仁数目较多, 一般位于核的边缘(图 3); (3) 晚期:体积进一步增大, 细胞质嗜碱性反应减弱, 染色变浅,核仁多个(图3)。
Ⅲ时相卵母细胞 初级卵母细胞进入大生长期, 细胞质为弱嗜碱性。体积明显增大, 细胞直径203µm左右, 核径直径100µm左右, 核质比49%。其主要特点是细胞质膜外包围由二层滤泡细胞构成的滤泡膜, 外层细胞呈梭形, 有一大的核, 细胞之间的界限不清楚; 滤泡膜和细胞质膜之间出现薄层的放射膜(图 4); 细胞质中出现皮质液泡并由边缘逐渐向中央增多, 液泡的数量和大小随着卵母细胞的进一步生长发育而增加; 卵黄颗粒从细胞的边沿开始累积、不断增加并向中央扩展。此时, 核仁大部分都分布在核膜的边缘, 只有少数分散在核的中央。
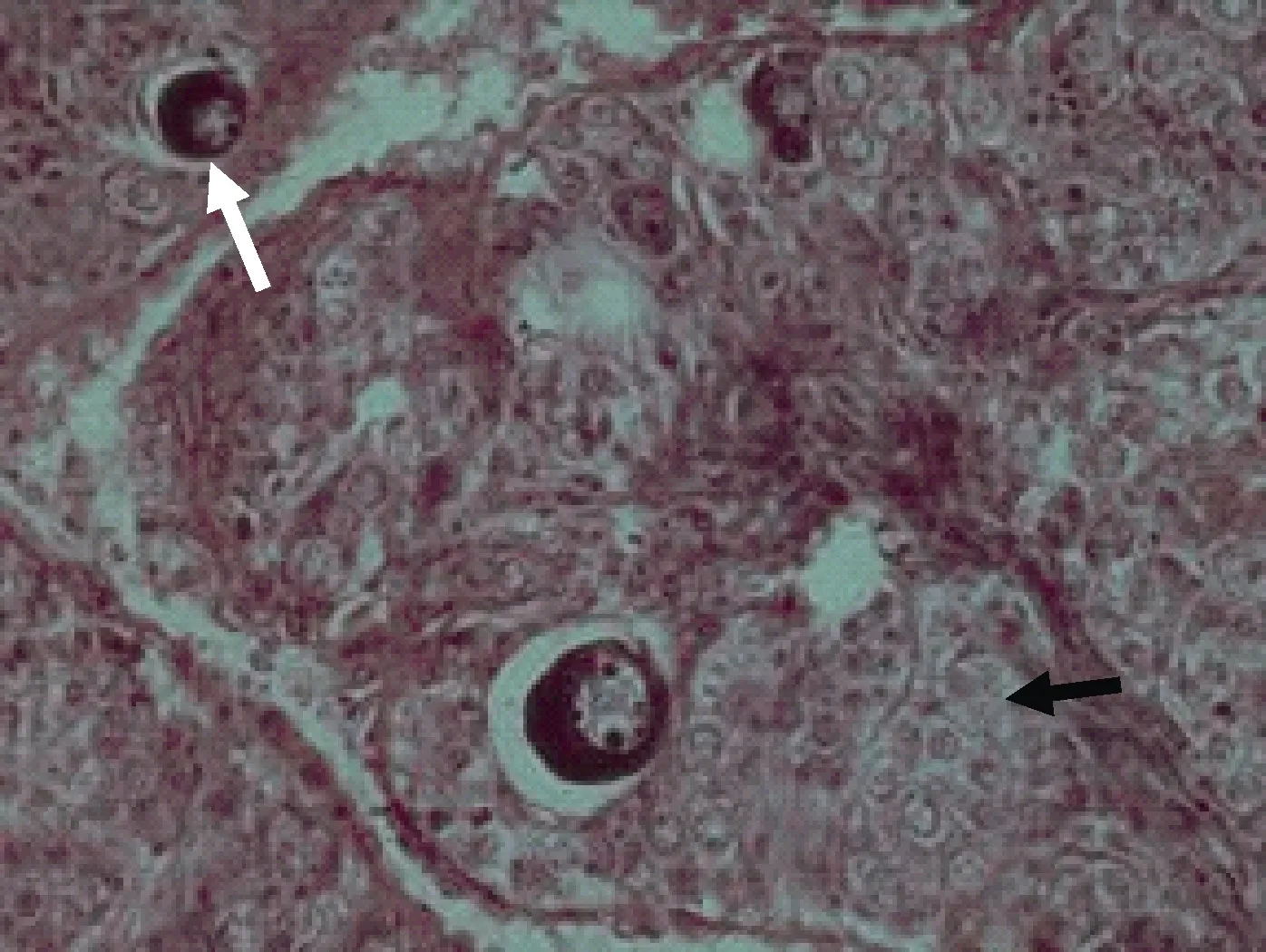
图2 Ⅱ时相早期卵母细胞(白色箭头)、卵原细胞(黑色箭头)(×400)Fig.2 The oocyte of the early stage in phaseⅡ (white arrow)and the oogonium in phase Ⅰ(black arrow) (×400)

图3 Ⅱ时相中期(虚线箭头)和Ⅱ时相晚期(实线箭头)卵母细胞(×400)Fig.3 The oocytes of the mid-stage(dotted arrow) and the late stage in phaseⅡ (solid arrow) (×400)

图4 Ⅲ时相卵母细胞(箭头)(×400)Fig.4 The oocytes in phase Ⅲ (arrow) (×400)
Ⅳ时相卵母细胞 为初级卵母细胞大生长期的后阶段, 即生长成熟阶段。细胞体积明显增大, 细胞直径平均362µm左右, 核径平均82µm左右, 核质比22%。细胞质中卵黄继续积累、增加并由细胞的边缘逐渐向中央扩展, 最终充满整个细胞, 而且卵黄颗粒不仅数量增加, 体积也不断增大而达到最终大小;细胞核从卵中央逐渐移向边缘, 最终到达动物极一端; 随着卵母细胞的成熟, 构成滤泡膜的两层滤泡细胞更加明显, 并且内层滤泡细胞变得比外层滤泡细胞圆而大; 放射膜增厚(图5)。
Ⅵ时相卵母细胞(闭锁滤泡) 本研究没有获得第5期卵巢和产后卵巢, 因此没有观察到卵母细胞排出后残留的滤泡细胞等形成的产后黄体及残留的退化中的未产出的Ⅳ时相和Ⅴ时相卵母细胞, 但在第Ⅱ期、第Ⅲ期和第Ⅳ期卵巢中都发现有退化的卵母细胞形成的闭锁滤泡, 可见卵母细胞的退化是存在于各个发育时期的普遍现象。闭锁滤泡的主要特征是:滤泡细胞增多、肥大, 而且排列不规则; 放射膜不规则, 局部增厚; 卵黄溃散、液化; 核膜崩解, 细胞核消失或处于消失过程中(图6)。

图6 Ⅵ时相(退化中)的卵母细胞(×400)Fig.6 The oocyte in phase Ⅵ (atresic follicle) (×400)
2.2 生长期卵巢的组织学结构
通过显微镜检查, 根据卵巢中卵细胞不同发育时相, 以切片视野中数量占优势或者占面积比例最大的卵细胞类型为依据将发育到生长成熟期的卵巢划分为4期。
Ⅰ期卵巢 Ⅰ期卵巢的形状早期为透明的细线状, 晚期为扁平带状, 肉眼无法区分出雌、雄性别。雌鱼 4—6月龄时卵巢腔形成, 卵巢已分化完全。卵巢壁主要由上皮细胞和结缔组织构成, 在早期(刚分化的Ⅰ期卵巢)蓄卵板尚未形成。卵巢中的生殖细胞主要是第Ⅰ时相卵原细胞, 分散排列在卵巢基质中(图 7)。到比较晚的时期形成一个个的褶皱状的蓄卵板(图 8), 蓄卵板的内层(靠近卵巢腔一侧)主要由较有规则的、一团团的卵原细胞构成, 蓄卵板的中央为卵巢壁外层的结缔组织延伸来的间质(图 9), 卵巢切片的一侧有较大的血管。
Ⅱ期卵巢 8月龄至 25月龄的雌性红鳍笛鲷的卵巢处于第Ⅱ期, 卵巢长度增加, 体积增大, 外观呈圆柱状, 表面可见微血管分布, 呈肉红色。肉眼大体上可分辨出雌、雄性别, 但肉眼仍难看清卵粒, 固定后卵巢呈现花瓣状的分叶, 这就是切片上看到的蓄卵板。随着发育的进展, 切片中蓄卵板和卵巢腔都比较发达, 蓄卵板中央间质细胞所占空间比例减少(图11)。Ⅱ期早期的卵巢中主要是Ⅱ时相早期及部分Ⅱ时相中期的卵母细胞。Ⅱ期晚期卵巢中主要是Ⅱ时相晚期和中期的卵母细胞,也有少数Ⅲ时相的卵母细胞。Ⅱ期卵巢蓄卵板的边缘存在不少染成浅红色的卵原细胞。
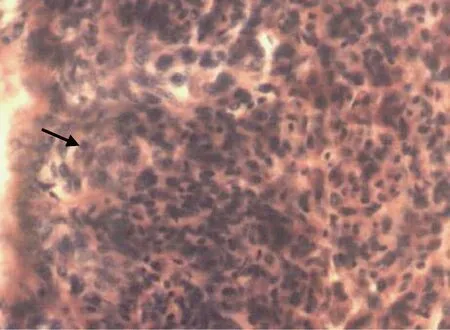
图7 第Ⅰ期卵巢早期(×400)Fig.7 The ovary at earlier stage Ⅰ (×400)卵原细胞较少, 分散排列

图8 第Ⅰ期卵巢晚期(×200)Fig.8 The ovary at later stage Ⅰ (×200)卵原细胞比早期增多, 排列变得有规律
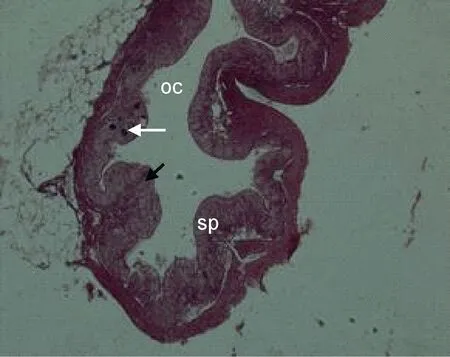
图9 Ⅰ期卵巢向Ⅱ期卵巢过渡Fig.9 The ovary transited from stage Ⅰ t o stageⅡ
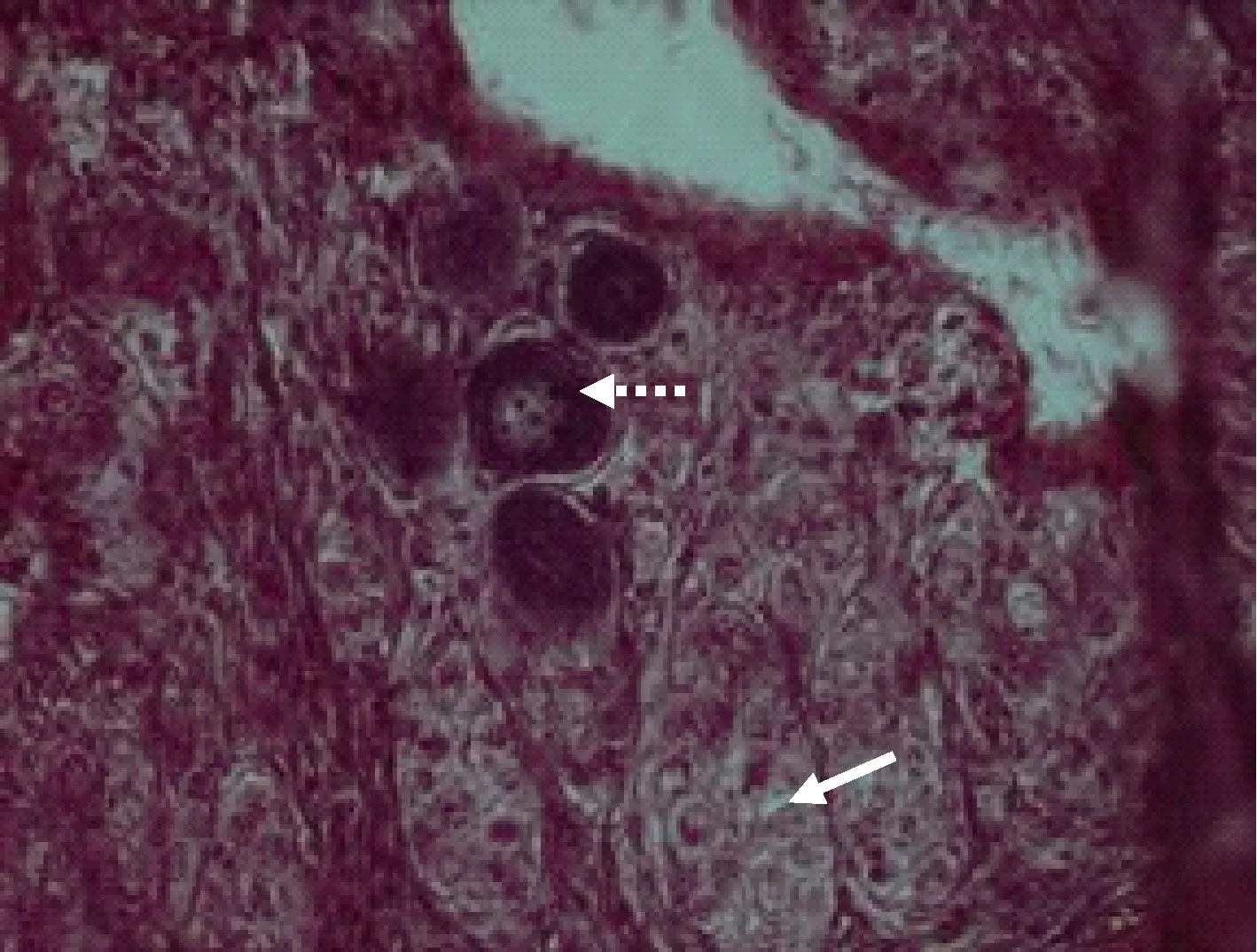
图10 图9的局部放大Fig.10 The close-up of figure 9
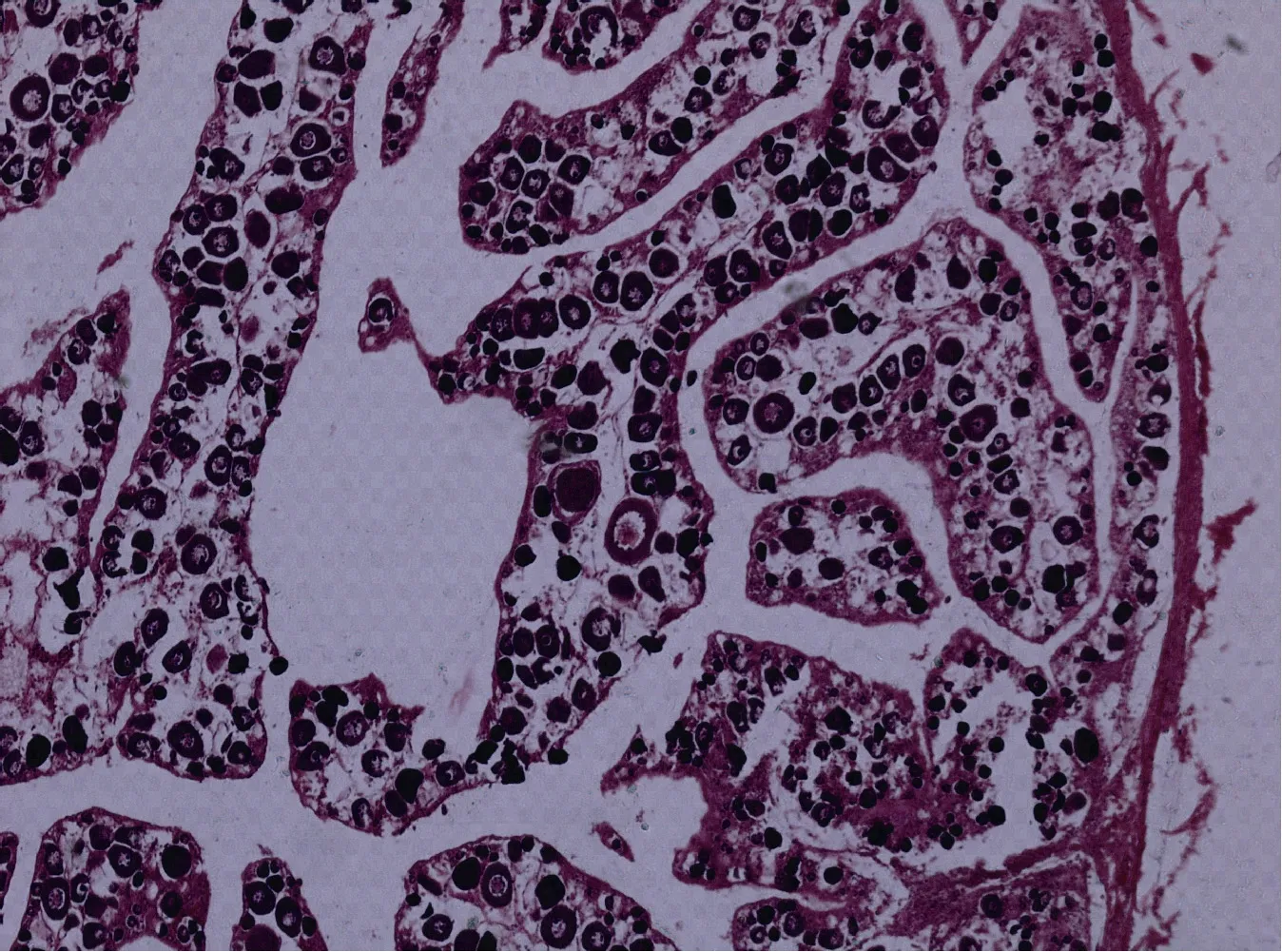
图11 第Ⅱ期卵巢(×50)Fig.11 The ovary at stage (×50Ⅱ)
Ⅲ期卵巢 25月龄以上雌性红鳍笛鲷的卵巢处于第Ⅲ期, 其体积较Ⅱ期卵巢明显增大、变圆, 表面有丰富的血管。Ⅲ期卵巢中卵母细胞主要有Ⅲ时相和Ⅱ时相, 也有少量的Ⅳ时相的卵母细胞, 作为卵母细胞来源提供者的卵原细胞总是存在的(图12)。
Ⅳ期卵巢 2.5—3龄红鳍笛鲷可达生长成熟,卵巢为Ⅳ期。卵巢为长椭圆形, 淡黄色, 表面血管发达。卵巢内卵细胞明显, 可见少量透明卵粒。卵巢内卵母细胞排列比较松散, 多数为Ⅳ时相卵母细胞和较多的Ⅲ时相卵母细胞, 也有数量不少的Ⅱ时相卵母细胞, 但只在蓄卵板的边缘可见少数分散分布的卵原细胞(图5, 图13)。

图12 第Ⅲ期卵巢(×50)Fig.12 The ovary at stage Ⅲ (×50)
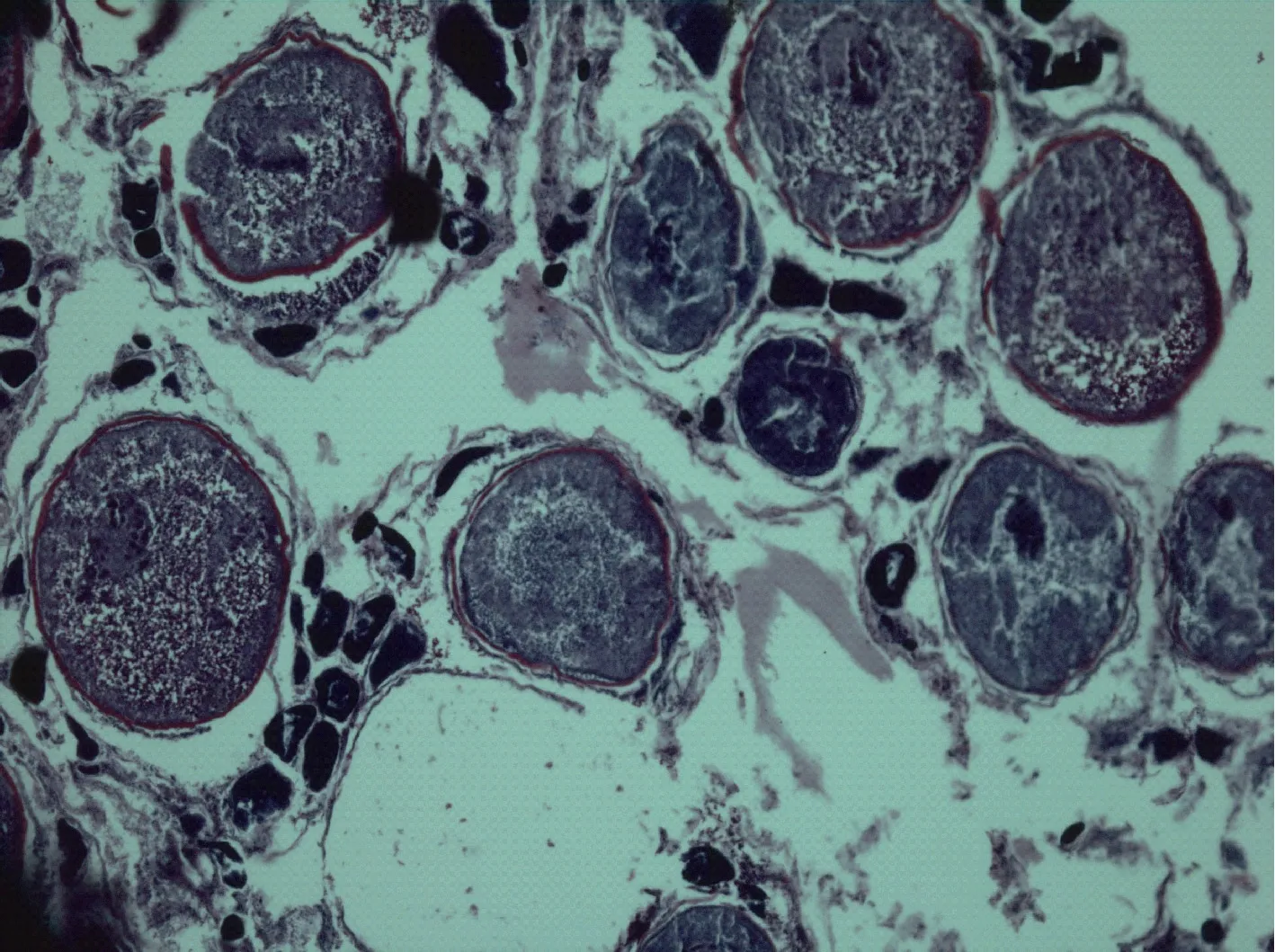
图13 第Ⅳ期卵巢(×50)Fig.13 The ovary at stage Ⅳ (×50)
2.3 第一个性周期内红鳍笛鲷卵巢的发育程序
本研究表明, 4月龄左右红鳍笛鲷卵巢已分化成形, 达到Ⅰ期卵巢, 卵巢内的生殖细胞主要是卵原细胞(图 7, 图 8, 图 9, 图 10)。8月龄时卵巢中部分卵原细胞分化为第Ⅱ时相早期的卵母细胞, 因此卵巢开始发育为Ⅱ期(图11), Ⅱ期卵巢一直维持到24月龄;25月龄红鳍笛鲷卵巢有部分可发育到Ⅲ期(图12), 其中有较多的 Ⅲ时相卵母细胞, 也有少量的Ⅳ时相的卵母细胞(图 5, 图 13), 但总体来说, 还是第Ⅲ期卵巢。此后, 随着生长发育, 卵巢在内、外环境条件的调控下进入成熟时期, 2.5—3龄鱼的卵巢可达到第Ⅳ期、Ⅴ期的成熟期。
3 讨论
3.1 关于卵巢腔的形成
关于脊椎动物生殖腺的形成(Gilbert, 2010), 如两栖类及哺乳类, 先是中肾两内侧腹膜增厚向体腔突出而形成生殖嵴, 原始生殖细胞迁入生殖嵴形成生殖腺, 然后生殖腺背部形成初级生殖腔。它充满了由间叶细胞形成的网索(原始性索)而成为髓部, 而周围的上皮和间质细胞构成皮层。当分化为雌性时, 髓部的网索消失产生许多空腔(次级生殖腔), 彼此连通而成为卵巢腔, 原始生殖细胞从网索迁入皮层而成为卵原细胞。许多研究者通过研究, 结果认为在不同的鱼类, 卵巢腔的形态、形成的时间和方式各不相同。Nakamura(1978)将卵巢腔形成分成3种方式: 一种是生殖腺分化时, 其外侧面上下各形成一个组织突, 两个突起相对延伸、融合而形成卵巢腔, 如银汉鱼(Odontesthes bonariensis)等(Strüssmann et al, 1996);第二种是腹膜壁在连接性腺的地方有一小团细胞逐渐延伸, 与已形成的生殖腺的边缘融合而形成卵巢腔,如 C. auratus及其它一些鲤科鱼类(Nakamura, 1978);另一种是性腺向体腔后面延伸, 与体腔后壁融合而形成卵巢腔, 鳉如青(Oryzias latipes) (Nakamura,1978)。但游秀容等(2012)对大黄鱼卵巢腔形成方式的研究结果与这 3 种方式都有区别, 是由卵巢腹部微血管旁的性腺组织向背方生长, 而卵巢柄状背部的体细胞增殖并向腹面生长, 二者融合而形成卵巢腔。本文从红鳍笛鲷较早期卵巢的组织学结构来看(图9),其卵巢腔的形成似乎类似于大黄鱼卵巢腔的形成方式, 由卵巢两端延伸、融合而形成。
3.2 关于雌性生殖细胞各时相名称及其与内容的统一
关于鱼类性腺发育分期, 国际上没有一个统一的标准(瑔施芳, 1988; 刘筠, 1993; 张贤芳等, 2006;代明山等, 2009)。最早对硬骨鱼雌性生殖细胞进行分期的应是前苏联学者B.A.Meйeн, 他将鱼类卵母细胞的生长期分为胞核改变期、小生长期和大生长期三个时期。然而, 这个分期方法既不系统, 也不全面, 例如, 他对胞核改变期的描述包括细线期、偶线期、粗线期、双线期等, 实际上这是减数分裂前期染色体演变的四个阶段。但这种染色体演变程序不一定就只局限于他所指的稚龄时相之前的胞核改变期。另外, 该分期方法中也没有包括卵原细胞期。在科研和生产应用中, 我国科技工作者, 在Meйeн的分期方法的基础上不断加以改进和完善, 根据硬骨鱼卵巢的组织学特征, 现已基本上统一将其分为6期, 根据卵细胞的细胞学特征, 将其生长发育过程分为6个时相, 其方法、内容和名称日趋一致(徐亚飞等, 2015), 但是仍不是很统一, 有的把时相的划分与卵巢的分期基本能对应起来, 有的不能对应, 例如, 大多将第Ⅰ时相的定义为卵原细胞期(有的包括由卵原细胞向初级卵母细胞过渡的阶段), 有的将第Ⅰ时相定义为由卵原细胞向初级卵母细胞过渡的细胞构成(孙鹏等, 2013);有的虽然将卵原细胞划分为第Ⅰ时相, 而在对卵巢进行分期时, 又没有包括卵原细胞; 还有, 对第Ⅰ时相的名字和内容也有不一致的地方或者文字与图不一致, 例如有的称为第Ⅰ时相卵母细胞, 但其描述又是卵原细胞期(曹伏君等, 2010; 马世磊等, 2014; 曲焕韬等, 2015; 李培伦等, 2015)。本文作者认为, 经过如此长时间的研究和应用, 现在应该可以形成一致的概念了, 根据大多数人的提法, 可将第Ⅰ时相的定义为卵原细胞期, 第Ⅰ期卵巢主要由卵原细胞和少数刚向初级卵母细胞分化的细胞组成; 卵原细胞一旦不再进行有丝分裂而进行分化, 便进入初级卵母细胞的早期阶段, 因此可以定义为第Ⅱ时相卵母细胞。
3.3 红鳍笛鲷的性成熟与产卵类型
在对性腺发育研究的基础上, 学者们将硬骨鱼类的产卵类型分为一次性产卵和多次性产卵。一次产卵类型, 是指一年一度产卵一次, 如四大家鱼, 其卵母细胞生长发育的特点主要是: 从Ⅲ时相至Ⅳ时相是基本上是同步的, 全产后的卵巢处于第Ⅱ期, 没有或很少有次发性的Ⅲ时相和Ⅳ时相卵母细胞, 在一年之内, 卵巢成熟系数只在繁殖季节出现一次高峰(刘筠, 1993)。多次产卵类型则是在一年之内可以产卵多次, 如尼罗罗非鱼, 其卵母细胞和卵巢在生长发育过程中的主要特点是:由Ⅲ时相发育到Ⅳ时相是非同步性的, 在繁殖季节, 第Ⅳ期卵巢中Ⅰ、Ⅱ、Ⅲ、Ⅳ时相卵母细胞同时存在, 并且Ⅳ+、Ⅳ++、Ⅳ+++同时存在, 产卵后的卵巢组织学结构仍然是Ⅳ期, 在一年内卵巢成熟系数明显地形成多次波峰和波谷(刘筠等,1983)。但不同研究者对同一种鱼产卵类型的研究曾获得了不同的结果, 如有些研究者认为草鱼是一次性产卵类型(刘筠等, 1978), 而有的研究者则认为是多次产卵类型(林光华等, 1985)。这可能与环境因素的影响及饲养条件有关(中山大学生物系动物学教研室等, 1978; 瑔施芳, 1992; 林浩然, 1999)。在本研究的取材过程中发现, 鱼的性成熟年龄和产卵类型可能确与环境及饲养条件相关, 例如, 在海南省南部红鳍笛鲷从3月到12月都可以产卵, 作者在6月和12月分别在海南省三亚市红沙龙胜鱼苗场和海南三亚蜈支洲岛鱼苗场现场进行了早期胚胎发育的活体观察, 而与海南只琼州海峡之隔的湛江, 红鳍笛鲷在秋冬季不能进行繁殖, 说明环境因素对其性成熟和产卵的影响很大; 同时, 从本研究的性腺发育取材来看,10月龄的鱼的个体(平均体重678.3g)比8月龄的个体(平均体重 778.3g), 前者的卵巢刚开始从Ⅰ期卵巢向Ⅱ期卵巢分化, 只有极少数Ⅱ时相的卵母细胞, 而后者的卵巢内已有较多的Ⅱ时相的卵母细胞, 其发育比 10月龄的要快, 也说明不同养殖条件下会对性成熟产生影响。
也有很多学者将卵巢发育分为完全同步型、部分同步型(分批同步型)和不同步型(分批非同步型)(施瑔芳, 1988; 林浩然, 1999; 张贤芳等, 2006)。作者认为这种划分实际上是对一次产卵和多次产卵类型提法的补充和完善, 特别是分批同步型(部分同步型)能够解释同种鱼在不同环境条件下在一年内一次产卵或多次产卵。完全同步型是指卵巢内卵母细胞处于相同的发育阶段或者卵子发育基本同步, 成熟系数每年只出现一次高峰期, 比如南方鲇(张耀光等, 1996)、大弹涂鱼(蔡泽平, 1994)以及前述的四大家鱼等, 实际应是属一次性产卵, 鳠而斑在人工驯养条件下卵巢发育为非同步型, 但其产卵类型仍为为一次产卵类型, 其产卵结束后, 卵巢中虽然仍存在大量的Ⅲ期、Ⅳ期卵母细胞, 但发育到Ⅴ期需要较长的时间和大量的营养积累, 鳠斑一次产卵结束后, 需要摄取大量的营养物质, 为下一年产卵做准备(谢少林等,2014)。部分同步型的卵巢一年只成熟一次, 但断续进行几次产卵, 产卵时间有一定的间隔, 如纳木错裸鲤(何德奎等, 2001)等; 人工养殖半滑舌鳎(陈彩芳等,2010)、圆斑星鲽(徐永江等, 2011)等, 也是 1年中只有1个繁殖期(1次成熟), GSI在一年的产卵期内仅出现一个峰值区, 也应属于分批同步型。分批非同步型可连续多次产卵, 比如革胡子鲇(林光华等, 1995)、旗鱼(Arocha, 2002)、罗非鱼(刘筠等, 1983)等, 属于典型的多次产卵类型。
本文从红鳍笛鲷第Ⅳ期卵巢中存在各个不同发育等级的生殖细胞来看, 符合分批非同步型性腺的特征, 在生产实践中, 一年也能进行多次繁殖, 因此,红鳍笛鲷应是属于多次产卵类型, 当然红鳍笛鲷的产卵类型究竟是属于分批同步型(部分同步型)还是分批非同步型(不同步型)的准确结论还是要通过性成熟后性腺的周年变化情况的研究才能得到。
马世磊, 耿 智, 徐冬冬等, 2014. 黄姑鱼性腺发育的组织学观察. 浙江海洋学院学报(自然科学版), 34(2): 129—133
中山大学生物系动物学教研室, 广东省南海县水产养殖场,1978. 草鱼人工繁殖中一年多次产卵的生物学基础. 水生生物学集刊, 6(3): 261—272
代明山, 刘庆武, 2009. 浅谈硬骨鱼类卵巢的形态结构和分期标准. 黑龙江水产, (2): 17—18
曲焕韬, 刘 勇, 杨元金等, 2015. 长 鳍吻 鮈 卵巢发育组织学及周年变化的初步研究. 水产科学, 34(1): 32—37
刘 筠, 1993. 中国养殖鱼类繁殖生理学. 北京: 农业出版社,22—41
刘 筠, 刘国安, 陈淑群等, 1983. 尼罗罗非鱼性腺发育的研究. 水生生物学集刊, 8(1): 17—30
刘 筠, 陈淑群, 王义铣等, 1978. 草鱼产卵类型的研究. 水生生物学集刊, 6(3): 247—257
孙 鹏, 尹 飞, 施兆鸿等, 2013. 养殖银鲳卵巢发育的组织学观察. 中国水产科学, 20(2): 293—298
芮菊生, 1980. 组织切片技术. 北京: 人民教育出版社
李培伦, 刘 伟, 王继隆, 2015. 乌苏里白鲑洄游群体性腺发育组织学观察. 生物学杂志, 32(1): 34—38
杨春玲, 刘楚吾, 2010. 红笛鲷(Lutjanus sanguineus)性腺发育初步研究. 广西农业科学, 41(4): 375—379
何德奎, 陈毅峰, 蔡 斌, 2001. 纳木错裸鲤性腺发育的组织学研究. 水生生物学报, 25(1): 1—13
张贤芳, 张耀光, 王志坚, 2006. 硬骨鱼类的卵巢发育和卵子发生研究综述. 海南师范学院学报(自然科学版), 19(1):70—78
张耀光, 谢小军, 1996. 南方鲇的繁殖生物学研究: 性腺发育及周年变化. 水生生物学报, 20(l): 8—16
陈武各, 2002. 红鳍笛鲷人工育苗技术. 渔业现代化, 29(4):20—21
陈彩芳, 温海深, 陈晓燕等, 2010. 人工养殖半滑舌鳎卵巢发育及其产卵类型研究. 海洋科学, 34(8): 29—34
林光华, 翁世聪, 张丰旺, 1985. 性成熟草鱼卵巢发育的年周期变化. 水生生物学报, 9(2): 186—193
林光华, 熊敬维, 1995. 革胡子鲇卵巢在第一次性周期内分化与发育的研究. 动物学研究, 16(4): 365—372
林浩然, 1999. 鱼类生理学. 广州: 广东高等教育出版社,185—188
郑乐云, 方琼珊, 王涵生, 2004. 红鳍笛鲷亲鱼培育及产卵技术研究. 海洋科学, 28(8): 1—4
施 瑔 芳, 1988. 鱼类性腺发育研究新进展. 水生生物学报,12(3): 248—258
施 瑔 芳, 1992. 我国鱼类生殖生理学研究概况. 海洋与湖沼,23(3): 325—333
徐永江, 柳学周, 刘君刚等, 2011. 圆斑星鲽卵巢发育的组织学和数量形态特征研究. 渔业科学进展, 32(3): 7—15
徐亚飞, 陈再忠, 高建忠等, 2015. 人工养殖七彩神仙鱼性腺发育的研究. 安徽农业大学学报, 42(1): 115—123
曹伏君, 罗 杰, 刘楚吾, 2010. 大弹涂鱼性腺发育的组织学观察. 水生生物学报, 34(2): 418—425
彭银辉, 刘楚吾, 郭昱嵩等, 2008. 三种笛鲷的野生群体和养殖群体遗传多样性的微卫星分析. 农业生物技术学报,16(5): 810—814
游秀容, 蔡明夷, 姜永华等, 2012. 大黄鱼性腺性别分化的组织学观察. 水产学报, 36(7): 1057—1064
谢少林, 房伟平, 陈金涛等, 2014. 养 殖斑 鳠 的性腺发育及其组织学观察. 大连海洋大学学报, 29(2): 147—150
蔡泽平, 1994. 大弹涂鱼卵巢发育的组织学研究. 热带海洋,13(1): 31—38
Arocha F, 2002. Oocyte development and maturity classification of swordfish from the north-western Atlantic Ocean. J Fish Biol, 60(1): 13—27
Gilbert S F, 2010. Developmental Biology. 9thed. Sunderland:Sinauer Associate, Inc, Publishers, 473—478
Nakamura M, 1978. Morphological and experimental studies on sex differentiation of the gonad in several teleost fishes.Hokkaido, Japan: Ph.D. Thesis of Hokkaido University
Strüssmann C A, Takashima F, Toda K, 1996. Sex differentiation and hormonal feminization in pejerrey Odontesthes bonariensis. Aquaculture, 139(1—2): 31—45
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21
中国临床医学影像杂志(2018年2期)2018-08-02
中国医学影像技术(2018年5期)2018-05-18
中国医药指南(2017年3期)2017-11-13
现代电生理学杂志(2016年4期)2016-07-10
中国卫生标准管理(2015年5期)2016-01-14
天津医科大学学报(2015年2期)2015-12-22
现代检验医学杂志(2015年1期)2015-02-06
中国医学装备(2015年2期)2015-01-30
食品工业科技(2014年15期)2014-03-11
