
中国沿海多鳞四指马鲅(Eleutheronema rhadinum)与四指马鲅(E. tridactylum)形态与遗传位点差异分析*
2016-01-15 03:51赵优庄平张涛赵峰
海洋与湖沼 2016年1期
赵 优 庄 平 张 涛 赵 峰
(1. 上海海洋大学水产与生命学院 上海 201306; 2. 中国水产科学研究院东海水产研究所 农业部东海与远洋渔业资源开发利用重点实验室 上海 200090; 3. 唐山出入境检验检疫局 唐山 063000)
多鳞四指马鲅(Eleutheronema rhadinum)与四指马鲅(E. tridactylum)均隶属于辐鳍鱼纲(Actinopterygii)、鲈形目(Perciformes)、马鲅科(Polynemoidea)、四指马鲅属(Eleutheronema)(庄平等, 2006)。由于多鳞四指马鲅与四指马鲅在形态上相似度高, 曾一度将多鳞四指马鲅与四指马鲅混淆, 直到 Motomura等(2002)对四指马鲅属鱼类进行重新划分, 才将多鳞四指马鲅从四指马鲅种中划分出来, 并确定四指马鲅属包括多鳞四指马鲅、四指马鲅、三趾四指马鲅(Eleutheronema tridactylum)三个种。目前, 在我国常见为多鳞四指马鲅与四指马鲅, 主要分布在我国东海及南海海域。多鳞四指马鲅及四指马鲅均为大型经济鱼类, 具有生长速度快、盐度适应范围广、经济价值高等特点, 是海水鱼类中的名贵种, 具有较好养殖开发前景, 已被 FAO作为重点推广的养殖鱼类(Rainboth, 1996)。但由于对四指马鲅属鱼类研究起步较晚, 仅在资源调查(蒋日进等, 2008; 陈渊戈等,2011; 龚小玲等, 2011)、遗传结构(林少珍等, 2012;Horne et al, 2013; Sun et al, 2013; Wang et al, 2014)、生物学(黄桂云等, 2012; 常有民等, 2013)及人工繁育(毛连环, 2009)等方面有相关报道, 特别是利用遗传结构开展资源鉴定方面的研究较为罕见, 目前仅靠个别形态特征进行多鳞四指马鲅和四指马鲅鱼类的物种鉴定, 给两物种资源评估与人工养殖开发带来一定困难。
形态学具有易操作、受样品状态影响小、结果直观等优势, 已被许多研究者应用于鱼类群体及物种间差异分析的研究中。多变量形态度量学方法是基于框架位点而引入多元统计的一种分析方法, 是对鱼类外部形态的连续性特征差异的分析, 可更加全面反应种内及种间差异(Booksteinet al, 1985)。简化基因组测序技术是目前最高效的群体分析的手段之一,近年发展起来的方法主要有GBS、RAD-seq、SLAF-seq等。该技术通过选取合适的限制性内切酶结合高通量群体测序构建 SNP分子标记, 性价比高、稳定性好,可广泛用于群体进化分析、高密度遗传图谱构建、QTL定位以及辅助基因组组装连接染色体等领域(陈士强等, 2013)。基于SLAF-seq技术的SNP多态分子标记以及 InDEL分子标记的开发, 通过测序扫描整个基因组中存在的单核苷酸多态性(SNP)、结构变异(SV)等遗传变异, 开展遗传图谱研究, 完善鱼类育种和选择技术。
针对多鳞四指马鲅与四指马鲅种质鉴定研究较为缺乏的现状, 本研究首次应用框架形态学方法对多鳞四指马鲅及四指马鲅两物种形态差异进行比较,并利用简化基因组测序技术(SLAF-Seq), 对我国两种易混淆的四指马鲅鱼类进行多态性标记分析, 获得大量的变异信息(SNPs、InDels), 为进一步开展多鳞四指马鲅及四指马鲅物种资源评估与管理、人工选育和功能基因开发等提供基础参考。
1 材料与方法
1.1 材料
本实验于2013年11—12月, 在湛江海域采集多鳞四指马鲅与四指马鲅样本。所有样本采用Motomura等(2002)形态鉴定方法进行初步鉴定后,用于形态数据测量, 并取背部肌肉放置于 95%的酒精中, 低温保存。观察样本数量和规格见表1。

表1 多鳞四指马鲅与四指马鲅群体观测样本数量和规格Tab.1 The number and size of samples of E. rhadinum and E. tridactylum populations
1.2 形态学分析
1.2.1 形态学数据测量与校正 测量数据分为传统形态学测量数据和框架形态数据两部分。传统形态数据包括全长(TL)、头长(HL)、肛前体长(LBA)、叉长(FL)、体高(BH)、眼后头长(PL); 并参照杨阳等(2013)的框架测量方法建立多鳞四指马 鮁 框架定标点, 形态学测量参数用钢尺和游标卡尺测量, 精确至0.1mm, 框架数据测量图见图1。
1.2.2 形态学数据分析 为消除样品个体规格差异对形态分析的影响, 采用Reist(1985)的方法对原始测量数据进行校正, 校正公式为:e= exp[lnY–b(lnX–lnXL)]。其中e为校正值,Y为测量值,b为lnY与lnX的斜率,X为样本体长,XL为所有样本的平均体长。校正后可量性状及框架性状的标准数据运用 SPSS(version 19.0)统计软件进行分析。
采用利用多变量方差分析方法对多鳞四指马鲅群体及四指马鲅群体的26个形态特征值进行主成分分析(principal component analysis, PCA), 提取主成分,并对主成分中相关变量贡献率进行分析。

图1 多鳞四指马鲅与四指马鲅的框架测量图(杨阳等,2013)Fig.1 The framework measurement diagram of E. rhadinum andE. tridactylum
应用逐步判别法对多鳞四指马鲅及四指马鲅群体进行判别分析(discriminant analysis)。
判别准确率的计算公式为:
判别准确率P1=(O/M)×100%;
式中:O=该群体判别正确的尾数,M=该群体实际尾数,Ai=第i个群体中判别正确的尾数,Bi=第i个群体中的实际尾数,k=群体数。将校正数据在 SPSS 19.0软件运用逐步判别法进行判别分析, 构建2个群体的判别函数。
1.3 简化基因组测序
采用蛋白酶 K和苯酚/氯仿法提取基因组 DNA(马洪雨等, 2006), 以半滑舌鳎(Cynoglossus semilaevis)(GC含量40.81%)作为参考基因组设计酶切方案。分别取多鳞四指马鲅和四指马鲅基因组DNA各500 ng,加 0.6 UMseI (NEB, Hitchin, Herts, UK)、T4 DNA 连接酶(NEB)、ATP (NEB)和MseI接头(NEB)在 37℃下反应15 h, 65℃退火1 h, 然后加限制性内切酶HaeIII和BfaI在37℃下反应3 h。反应结束后用Quick Spin column (Qiagen)纯化酶切产物。
以回收的酶切产物为模板进行 PCR反应, 反应体系 25μL, 包括模板 DNA 100ng、10×buffer 2.5 μL、2.5 mmol/L dNTP 2 μL、5U的Taq DNA合成酶(NEB)0.3 μL、10 μmol/LMseI引物 2 μL、ddH2O 加至 25 μL。PCR产物经E.Z.N.A. Cycle Pure Kit (Omega)纯化后,加入MseI、T4 DNA连接酶、ATP及Solexa接头, 37℃下反应5 h。利用Quick Spin Column (Qiagen)纯化产物, 2%琼脂糖胶电泳, 并以其为模板, 在含Phusion Master Mix (NEB)和Solexa引物混合物中进行 PCR扩增, 扩增产物经纯化后用 Illumina GAIIx(Illumina, San Diego, CA, USA)测序。
2 结果与分析
2.1 两群体形态差异分析
2.1.1 可数性状比较 从外部形态特征比较, 多鳞四指马鲅无梨骨齿板, 胸鳍基部有黑色斑块, 侧线鳞83—97, 侧线上鳞12—14, 侧线下鳞15—17; 四指马鲅有梨骨齿板, 胸鳍基部有黄色斑块, 侧线鳞71—79, 侧线上鳞 9—12, 侧线下鳞 13—15。两群体可数性状特征比较如表2所示。

表2 多鳞四指马鲅与四指马鲅群体形态特征比较Tab.2 The morphological comparison between E. rhadinum and E. tridactylum populations
2.1.2 主成分分析 对多鳞四指马鲅群体与四指马鲅群体的形态测量数据进行主成分分析, 共提取了 7个主成分, 累积贡献率为 96.112%, 其中前三个主成分贡献率分别为28.759%、21.467%、15.469%, 累计贡献率 65.695%。在第一主成分中 D1-2、D1-4、D5-8、D6-8、D6-9、D7-9、D7-10、D7-11 的负荷绝对值较大, 主要反应的是鱼体头部、后半部分体侧及尾柄部特征; 第二主成分中头长、叉长、肛前体长、眼后头长、D9-11的负荷绝度值较大, 主要反应的是鱼体头尾轴特征; 第三主成分中D3-4 、D3-5、D3-8、D4-8的负荷绝对值较大, 反应的是鱼体前半部体侧的形态特征(详见表3)。
图 2为多鳞四指马鲅与四指马鲅群体形态分析前两个主成分的散点图, 从散点图分析, 利用前两个主成分可将多鳞四指马鲅群体与四指马鲅群体进行划分, 并且2个群体在主成分1坐标轴上的差异比主成分2坐标轴的差异显著。
2.1.3 判别分析 本研究利用逐步判别方法, 对2个群体野形态比例性状进行判别分析。根据逐步判别法, 筛选变量, 从特征性状中帅选出 PL、D9-11、D5-8、D6-8四个特征值进行判别分析, 综合判定准确率即可达到 100%, 该判别函数对于两物种的形态鉴定提供了重要参考。利用个形态特征值构建了多鳞四指马鲅群体与四指马鲅群体的判别函数, 其判别公式如下:

其中,X1—X4分别代表 PL、D9-11、D5-8、D6-8。将判别函数带入多鳞四指马鲅与四指马鲅群体进行检验, 综合判定准确率为100%(表4)。
2.2 两群体遗传位点差异分析
2.2.1 SLAF标签信息统计 利用多鳞四指马鲅及四指马鲅样本总共开发出 SLAF标签数为 140973个, 多鳞四指马鲅样品得到SLAF标签109005个, 四指马鲅样本得到SLAF标签 93036个。其中, 多鳞四指马鲅与四指马鲅共有SLAF标签33428个。图3显示对两份样本得到的140973个SLAF标签进行分型,最终得到 3种类型的 SLAF标签, 包括重复序列(Repeat) 9619个(6.82%), 多态性标签(Marker) 32216个(22.85%), 非多态标签(No Poly) 99138个(70.32%)。根据开发得到的32216个Marker, 利用GATK进行基于2个样品的群体内部SNP以及INDEL检测。多鳞四指马鲅及四指马鲅样品之间共开发出 SNP 68992个, InDel 12884个。

表3 2个群体形态变量主成分分析中的因子负荷矩阵的特征向量及累计贡献率Tab.3 Eigenvectors and cumulative contribution rates of factor loadings in principal component analysis on morphological variation in two populations
2.2.2 SNP及InDel信息统计 表5所示利用简化基因组测序技术, 开发出多鳞四指马鲅与四指马鲅SNP 69207个, 其中转换类型中: C/T 22458个(32.45%), A/G类型 22173个(32.04%); 颠换类型中A/T 7900 (11.42%); A/C 6171 (8.92%); T/G 6040(8.73%); C/G 4465 (6.45%)。
多鳞四指马鲅样本和四指马鲅样本之间总共开发出 12884个 InDel, 包含转化类型 773个(6.00%),颠换类型8031个(62.33%), 样品中杂合类型4080个(31.67%)。
3 讨论
3.1 多鳞四指马鲅与四指马鲅形态学分析

图 2 2个群体形态特征前两个主成分分布图Fig.2 The distribution of first 2 principal components in morphological indices of the two populations

表4 多鳞四指马鲅与四指马鲅群体判别分析结果Tab.4 The discrimination results between E. rhadinum and E.tridactylum populations

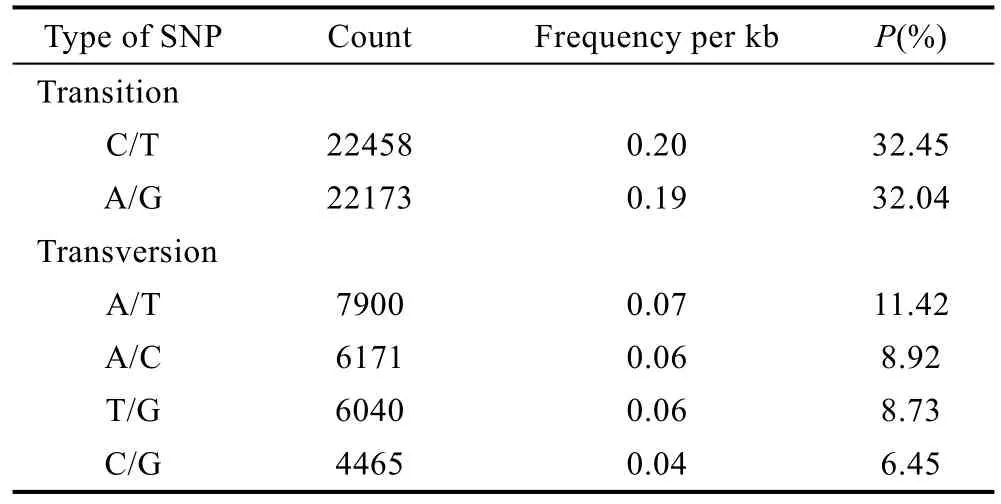
表5 2个群体间SNP类型统计Tab.5 The statistical results of SNP type between E. rhadinum and E. tridactylum populations
早在 19世纪, 研究学者 Bleeker(1862)开始对多鳞四指马鲅进行分类鉴定, 将其鳞片大小、有无犁骨齿板等定义为区分四指马鲅属鱼类的主要形态特征。Motomura等(2002)利用测定鱼体可量性状、可数性状以及外部典型特征区分四指马鲅属多鳞四指马鲅、四指马鲅、三趾四指马鲅三个物种。但传统的鱼类形态学研究参数主要遵循Hubbs等(1947)提出的在鱼体头尾轴及背腹轴水平及垂直方向的测定方法, 而框架测量法利用鱼体的解剖学同源坐标点, 可有效反映鱼体纵向、斜向和中间部分的许多信息(Ergudenet al,2005)。目前国内外已广泛开展了利用多变量形态学方法对鱼类形态差异的比较研究, 如狼鲈(Dicentrarchus labrax) (Ergudenet al, 2005)、澳洲鲐(Scomber australasicus) (Tzeng, 2004)、鲻鱼(Mugil cephalus) (刘建勇等, 2009)等, 均得到较好的判定效果。
从主成分分析结果来看, 多鳞四指马鲅与四指马鲅形态差异主要集中体现在鱼体头背部、尾柄部、头尾轴及背腹轴的特征上。两群体形态特征差异与栖息的水域环境及游泳运动相关, 以适应其生活环境及洄游、摄食等生活习性。其中背腹轴及头尾轴差异影响了鱼体的体型特征, 头尾轴长、背腹轴短的鱼类体形为纺锤型, 在水中游动阻力小, 承受水压能力强,更适宜在水流湍急的环境中游动; 尾柄则起到平衡及前进助推的作用; 而鱼体头背部形状对其游泳速度可产生影响。目前已有研究资料报道, 多鳞四指马鲅与四指马鲅主要栖息在河口近岸(Motomuraet al,2002), 多鳞四指马鲅具有生殖洄游习性, 幼鱼在河口索饵育肥(陈渊戈等, 2011; 龚小玲等, 2011)。多鳞四指马鲅与四指马鲅具有纺锤型体形, 对两种群适宜沿岸近海水流湍急的栖息环境及洄游习性具有重要意义。目前关于多鳞四指马鲅及四指马鲅背景资源缺乏, 后续可结合两个物种资源调查数据对两种群的栖息地、生活习性、摄食方式等开展进一步的研究。
本研究利用 4个形态特征值建立了多鳞四指马鲅与四指马鲅判别函数, 判别函数经回代检验综合判别准确率达到 100%, 相比传统形态学鉴定, 该方法更加全面反映多鳞四指马鲅与四指马鲅的形态差异。赵峰等(2011)利用4个形态参数将三种鲳鱼100%予以区分; 王新安等(2008)利用多元分析方法建立七带石斑鱼不同地理群体的判别函数, 综合判定准确率同样达到了100%。本研究结果进一步证实利用传统形态学及框架数据进行多变量分析是一种理想的形态学鉴定方法。
3.2 两群体遗传位点差异分析
由于生物形态是物种多个性状的集合, 是遗传因子和环境因素共同作用的结果。因此分析及鉴定多鳞四指马鲅与四指马鲅种群差异, 应在形态学判定的基础上, 结合两群体遗传结构差异, 进一步进行比较研究。目前开发SNP标记的方法很多, 但大多技术繁琐、成本高。SLAF-seq技术是基于生物信息学、高通量测序仪等的高度自动化技术, 获得的海量序列信息可满足任何密度的全基因组分布, 以实现候选功能区域的精细定位, 数字化信号和高覆盖保证了获得标签的准确性, 高通量分子标记开发成本低于常规分子标记, 可以一次性获得大量的特异 DNA序列, 为开发大量的特异分子标记提供了序列基础。据北京百迈客公司统计, SLAF-seq技术开发分子标记的平均成本不到 AFLP技术的 1/8, 而其效率却高近 27倍, 与转录组技术相比在也可以较低成本获得大量标记。可见, 利用 SLAF-seq技术开发四指马鲅属鱼类特异分子标记具有很高的重复性、特异性及实用性。目前, 国内外尚无多鳞四指马鲅与四指马鲅遗传差异相关报道, 本研究利用 SLAF-seq技术首次对四指马鲅属两个重要经济物种进行遗传结构差异分析, 挖掘出两近似种遗传结构上存在的大量差异位点, 大大降低了试验成本, 由于方法原理差异, 本研究得出四指马鲅属两种鱼类的 SNP位点转换的概率远大于颠换概率, 此结果与利用转录组技术开发鲤鱼(Cyprinus carpio) (Li et al, 2015)SNP结果不同。本研究为后续开发多鳞四指马鲅与四指马鲅共显性分子标记, 在遗传结构上进行物种鉴定提供有力的基础研究参考, 也对开发四指马鲅属鱼类相关功能基因、群体遗传学、基因组作图、物种间比较基因组学等研究具有重要意义。
马洪雨, 郭金峰, 岳永生, 2006. 用改进的酚-氯仿法提取鱼类基因组DNA效果的分析. 家畜生态学报, 27(2): 85—87
王新安, 马爱军, 陈 超等, 2008. 七带石斑鱼(Epinephelus septemfasciatus)两个野生群体形态差异分析. 海洋与湖沼,39(6): 655—660
毛连环, 2009. 四指马鲅人工繁殖技术. 水产科技情报, 36(6):275—278
庄 平, 王幼槐, 李圣法等, 2006. 长江口鱼类. 上海: 上海科学技术出版社, 194—195
刘建勇, 杨廷宝, 2009. 我国沿海鲻鱼(Mugil cephalus)不同地理群体形态差异研究. 海洋与湖沼, 40(5): 572—576
杨 阳, 庄 平, 张 涛等, 2013. 多鳞四指马鲅 4个地理群体的形态差异. 上海海洋大学学报, 22(6): 849—854
陈士强, 秦树文, 黄泽峰等, 2013. 基于SLAF-seq技术开发长
穗偃麦草染色体特异分子标记. 作物学报, 39(4):727—734
陈渊戈, 张 宇, 钟俊生等, 2011. 长江口南支和杭州湾北岸碎波带水域仔稚鱼群聚的比较. 上海海洋大学学报, 20(5):688—696
林少珍, 王丹丽, 王亚军等, 2012. 基于COⅠ序列分析东海区四指马鲅(Eleutheronema tetradactylum)的种群遗传结构.海洋与湖沼, 43(6): 1261—1265
赵 峰, 马春艳, 庄 平等, 2011. 东海常见鲳属鱼类的形态差异及系统进化关系探讨. 海洋渔业, 33(2): 138—143
黄桂云, 张 涛, 赵 峰等, 2012. 多鳞四指马鲅幼鱼消化道形态学和组织学的初步观察. 海洋渔业, 34(2): 154—162
龚小玲, 张晓懿, 朱 敏等, 2011. 长江口九段沙湿地潮沟鱼类组成及其多样性. 上海海洋大学学报, 20(4): 517—524
常有民, 张 涛, 庄 平等, 2013. 多鳞四指马鲅耳石形态特征的观察. 海洋渔业, 35(1): 24—33
蒋日进, 钟俊生, 张冬良等, 2008. 长江口沿岸碎波带仔稚鱼的种类组成及其多样性特征. 动物学研究, 29(3):297—304
Bleeker P, 1862. Noticeichthyogque. (I—X). Verse Akad Amsterdam, 14: 123—141
Bookstein F L, Chernoff B, Elder R et al, 1985. Morphometrics in Evolutionary Biology. Philadelphia: Academy of Natural Sciences of Philadelphia, 182—191
Erguden D, Turan C, 2005. Examination of genetic and morphologic structure of sea-bass (Dicentrarchus labrax L.,1758) populations in Turkish coastal waters. Turk J Vet Ani Sci, 29(3): 727—733
Horne J B, Momigliano P, van Herwerden L et al, 2013. Murky waters: searching for structure in genetically depauperate blue threadfin populations of western Australia. Fish Res,146: 1—6
Hubbs C L, Lagler K F, 1947. Fishes of the Great Lake Region.Michigan: University of Michigan Press, 26
Li G X, Zhao Y L, Liu Z H et al, 2015. De novo assembly and characterization of the spleen transcriptome of common carp(Cyprinus carpio) using Illumina paired-end sequencing.Fish Shellf Imm, 44(2): 420—429
Motomura H, Iwatsuki Y, Kimura S et al, 2002. Revision of the Indo-West Pacific polynemid fish genus Eleutheronema(Teleostei: Perciformes). Ichthyol Res, 49(1): 47—61
Rainboth W J, 1996. Fishes of the Cambodian Mekong. In: FAO Species Identification Field Guide for Fishery Purposes.Rome: FAO, 38—45
Reist J, 1985. An empirical evaluation of several univariate methods that adjust for size variation in morphometric data.Canadian Journal of Zoology, 63(6): 1429—1439
Sun X X, Xu D D, Lou B et al, 2013. Genetic diversity and population structure of Eleutheronema rhadinum in the East and South China Seas revealed in mitochondrial COI sequences. Chin J Oceanol Limnol, 31(6): 1276—1283
Tzeng T D, 2004. Morphological variation between populations of spotted mackerel (Scomber australasicus) off Taiwan.Fish Res, 68(1—3): 45—55
Wang J J, Sun P, Yin F, 2014. Low mtDNA Cytb diversity and shallow population structure of Eleutheronema tetradactylum in the East China Sea and the South China Sea. Biochem System Ecol, 55: 268—274
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
科学大众(2020年10期)2020-07-24
当代陕西(2019年6期)2019-04-17
中国继续医学教育(2015年2期)2016-01-06
振动、测试与诊断(2014年6期)2014-03-01
现代检验医学杂志(2014年1期)2014-02-06
现代检验医学杂志(2014年5期)2014-02-02
中国火炬(2012年8期)2012-07-25
中国火炬(2011年11期)2011-07-25
