
荒漠草原不同生长年限中间锦鸡儿灌丛枝系构型特征
2016-01-15 09:41:24翟德苹,陈林,杨明秀等
浙江大学学报(农业与生命科学版) 2015年3期
关键词:生物量
荒漠草原不同生长年限中间锦鸡儿灌丛枝系构型特征
翟德苹, 陈林, 杨明秀, 宋乃平, 杨新国*
( 宁夏大学西北退化生态系统恢复与重建教育部重点实验室/西北土地退化与生态恢复省部共建国家重点实验室培育基地/
西部生态与生物资源开发联合研究中心,银川750021)
摘要基于分维数理论,对荒漠草原5个不同生长年限(未平茬16年、25年、37年生及平茬后1年、5年生)中间锦鸡儿(Caragana intermedia)灌丛的枝系构型变化进行分析,并探讨其生态适应策略。中间锦鸡儿灌丛的冠幅、株高、基径等指标基本随生长年限的增加呈先增大后降低趋势,未平茬中间锦鸡儿灌丛的地上生物量随林龄的增加呈下降趋势,老林龄(37年生)灌丛以自我更新的方式来抵制衰老的态势;平茬后1年与5年生中间锦鸡儿灌丛的枝径比(ratio of branch diameter,RBD)较未平茬16、25、37年林龄灌丛大,平茬后灌丛枝条间的承载力增大;中间锦鸡儿灌丛的总体分枝率(overall bifurcation ratio,OBR)以及低级枝的逐步分枝率(stepwise bifurcation ratio, SBR1∶2)随林龄增大而减小,平茬后1年生灌丛的OBR及SBR1∶2较未平茬3种年限林龄的灌丛大,但平茬后5年生灌丛的OBR和SBR1∶2急剧降低;未平茬灌丛的树冠分维数(crown fractal dimensions,CFD)较低,均小于1,且随林龄增大,树冠分维数降低。平茬后的灌丛CFD值大于1,尤以平茬后5年生灌丛为最大。未平茬灌丛的分枝分维数(bifurcation fractal dimensions, BFD)随林龄增大而减小,平茬后1年生灌丛BFD值大于未平茬灌丛,平茬后5年生灌丛BFD值最小。在构件水平上,16、25、37年生中间锦鸡儿灌丛出现衰退型发育的趋势。16年生中间锦鸡儿灌丛的枝系空间格局最为复杂,其占据空间的能力以及利用空间资源的潜能较高。但随着灌丛林龄的增加,中间锦鸡儿的枝系构型趋于简化,对空间资源的利用潜能和生态适应能力下降。平茬有利于刺激灌丛空间格局的发展,使灌丛枝系构型复杂化,但平茬后生长5年的灌丛,其构型发展主要以各级分枝枝长为主,分枝数对其贡献不大。
关键词中间锦鸡儿; 生物量; 枝序; 分维数; 枝系构型
中图分类号S 718.42文献标志码A
基金项目:国家科技支撑项目“发达地区省级农村信息服务平台构建与应用”(2014BAD10B02)。
收稿日期(Received):2015-01-28;接受日期(Accepted):2015-04-18;网络出版日期(Published online):2015-05-19

Branching architectures ofCaraganaintermediaat different ages in desert steppe areas. Journal of ZhejiangUniversity(Agric. & LifeSci.), 2015,41(3):340-348
Zhai Deping, Chen Lin, Yang Mingxiu, Song Naiping, Yang Xinguo*(KeyLaboratoryforRestorationandReconstructionofDegradedEcosysteminNorth-WesternChinaofMinistryofEducation/BreedingBaseforStateKeyLaboratoryofLandDegradationandEcologicalRestorationofNorth-WesternChina/UnionResearchCenterforEcologyandExploitationofBiologicalResourcesinWesternChina,NingxiaUniversity,Yinchuan750021,China)
SummaryCaraganaintermediais a bush-like plant, and its branching architectures are more complex. Fractal theory provides a new tool for the study of trees and their properties. The crown characteristics of trees can be quantified by fractal dimensions, and the architectures can be described accurately.
The relationships between branching architectures and growth ofC.intermediawere studied from the viewpoint of fractal theory, in order to study the branching architectures and adaptation strategies to the environment ofC.intermediain desert steppe areas.
Based on the theory of fractal dimension in forestry application, the changes of branching architectures and strategies of ecological adaptation ofC.intermediaat different ages (growth for 16 a, 25 a, 37 a of intact plants and growth for 1 a, 5 a after stumping) were studied in desert steppe areas.
The indexes of crown size, height and basal diameter ofC.intermediaincreased firstly and then decreased with the increasing ages. Aboveground biomass of intactC.intermediaat different ages decreased gradually with the increasing ages. Older shrubs (37 a) resisted the situation of senescence by self renewing. The ratio of branch diameter (RBD) ofC.intermediashrubs which grew for one year after stumping was higher than those without stumping. The bearing capacity of shrub branches increased after stumping. The overall bifurcation ratio (OBR) and the lower-level stepwise bifurcation ratio (SBR1∶2) decreased with the increasing ages. The OBR and SBR1∶2of shrubs which grew for one year after stumping were higher than the shrubs without stumping, but OBR and SBR1∶2of shrubs which grew for 5 years after stumping decreased dramatically. The crown fractal dimensions (CFD) of shrubs without stumping were lower than 1, and CFD decreased with the increasing ages. The CFD of shrubs after stumping were higher than 1, especially the shrubs which grew for five years after stumping were the highest. The bifurcation fractal dimensions (BFD) of shrubs without stumping decreased with the increasing ages. The BFD of shrubs which grew for one year after stumping were higher than those without stumping, and the BFD of shrubs which grew for five years after stumping were the lowest.
At the module level,C.intermediaat the ages of 16 a, 25 a, 37 a presented a tendency of declining development. The branching pattern of 16 a shrubs was the most complex, and the ability to occupy the space and the potential of using spatial resources were higher. But the branching architecture ofC.intermediabecame simplified and the utilization potential of spatial resources and ecological adaptability declined with the increasing ages. Stumping was beneficial to stimulating the development of shrubs’ spatial patterns, and made branching architectures more complicated. The architectures development of shrubs which grew for 5 years after stumping mainly depended on branch length of different branch orders, and the branch number contributed little to it.
Key wordsCaraganaintermedia; biomass; branch order; fractal dimension; branching architecture
植物作为一种构件生物[1],其构件分为地面枝系和地下根系2个亚系统[2]。地上部分通过分生组织连续不断地产生新的分枝,并逐渐向外扩展,形成复杂多样的枝系结构和形态特征[3]。植物在特定的环境条件下形成其构型特征,这种形态学反应与其内部生理、生化性质存在一定的关联[4]。并且植物对其周围空间、光、热、水分、养分等资源的利用情况可通过植物体不同的枝系构型特征以及各构件单元的配置及其动态变化来表征,植株的枝系结构不仅是植物系统发育和形态学的研究内容,而且与植物的生存策略及环境适应性密切相关[5-6]。通过对植物枝系构型的研究,可揭示不同构件之间的相互作用、生物量的空间分配以及植物在长期进化过程中形成的生长模式,为进一步探讨植物的生长规律及种间竞争、群落演替等问题提供理论依据[7-8]。
然而,灌木枝系构型较为复杂,很难以常规的构型结构方法来研究并准确描述其构型特征[9-10]。分形几何理论经过几代人的发展和丰富,由美国著名数学家Mandelbrot将其完善。分形几何理论是一种把自然形态看作是具有无限嵌套层,并在各嵌套层保持某种相似属性的理论。用来描述传统欧氏几何理论无法研究的非规则几何形态,试图透过混乱现象和不规则构型揭示隐藏于现象背后的局部与整体的本质联系和运动规律[11-12]。分形几何所揭示的分形集的各组成部分在一定条件下或过程中,可能在某些方面表现出与整体的相似性,即具有自相似性(确定性的或统计意义上的),并能够用连续取值的分数维数来描述[13]。分维数是定量描述分形的一个重要参数,是分形几何中可以用分数值来度量分形体复杂程度的工具[12,14]。随着分形理论的不断完善与发展,为研究树木及其有关性质提供了一种新的工具,利用分维数可将林木的冠型特征进量化,且操作简单易行成为研究林木构型特征的理想指标[4,9,15-16]。
中间锦鸡儿(Caraganaintermedia)为豆科锦鸡儿属灌木类植物,主要分布在黄土高原和西北荒漠地区,具有抗旱、抗寒、耐热、耐沙埋等特性,防风固沙及保持水土的能力强[17];同时又是优质的灌木饲料植物资源[18-19],具有较高的生态经济价值。在宁夏退耕还林(草)工程中,人工柠条(锦鸡儿属的通称)林得到大面积推广种植[20-21]。人工柠条的进入,使得荒漠草原脱离沙化退化的境地,开始了新的演替发展。同时,柠条逐渐成为该区的优势植被类型。本文从分形角度研究了不同林龄中间锦鸡儿枝系构型特征与生长的关系,旨在探讨和研究荒漠草原中间锦鸡儿灌木枝系构型及其对环境的适应对策。
1研究地区概况与方法
1.1研究地区概况
研究地点选在宁夏盐池县杨寨子自然村(37°47′—37°57′N,107°22′—107°33′E),位于毛乌素沙地南缘,该区为典型的中温带大陆性气候。年均气温7.7 ℃,1月平均气温-8.9 ℃,7月平均气温22.5 ℃,绝对最高和最低气温分别为38.0 ℃和-29.5 ℃;≥0 ℃年积温3 430.3 ℃,≥10 ℃年积温2 949 ℃。年日照时数2 867.9 h,日照率65%。年均降水量250~350 mm,大部分集中在6—8月,且多暴雨,降水年变率较大;年蒸发量2 136 mm,是降雨量的6~8倍;年无霜期162 d。大地貌为缓坡丘陵,地带性土壤为灰钙土,还有大面积的风沙土、黑垆土等非地带性土壤,土壤质地以砂壤、粉砂壤和砂土为主。水质呈碱性,pH值为7.5~8.5。植被主要为自然或人工柠条灌木林及草、灌次生植被。
1.2研究方法
1.2.1灌丛植株选择及主要构型指标选择不同生长年限(1年,5年,16年,25年,37年)的中间锦鸡儿人工林,其中1年生和5年生的中间锦鸡儿林分别为平茬后生长1年和5年。在各样地挑选没有病虫危害、生长良好、具有典型代表性的灌丛3~5株,对灌丛植株基本形态指标和主要枝系构型指标分别进行调查。中间锦鸡儿属于落叶灌木,当年生叶构件主要集中于1~2年生枝上,因此当年生枝构件和叶构件的空间分布能够表现其分形特点。为了比较不同生长年限中间锦鸡儿灌丛的冠形分布,调查的基本指标包括:冠幅(长与宽)、株高、基径、老枝数、更新枝数、生物量、枝长、一级枝分枝数、二级枝分枝数等。在冠层内,由灌丛外向灌丛内以Strahler法确定枝序[22-25],最外层的第一小枝为第一级枝,2个一级枝相遇为第二级枝,2个二级相遇后则为第三级枝,依此类推,如有不同级别枝相遇,则取较高的作为枝级。
1.2.2构型指标的观测方法用钢卷尺(精度0.1 cm)测量每株待测灌丛的高度,作为株高。分别选择东-西和南-北2个方向采用钢卷尺测定每一等份处的长度,作为冠幅的长和宽。用钢卷尺测定不同枝序各级枝的长度,作为各级枝长。用游标卡尺(精度0.01 mm)测量每一分枝靠近根基部的直径,以及不同枝序各级分枝基部的直径作为基径。并计数灌丛中心老枝数与外围更新枝数,以及不同枝序的各级分枝数。
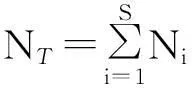
1.2.4分形维数计算方法针对荒漠植物的树冠及不同级别分枝在空间分布中的分维数进行计算[26-27]。1)树冠分维数(crown fractal dimensions,CFD):CFD=-lnr/lnk,式中:r为每一级分枝数,k为连续分枝级别枝长之比。2)分枝分维数(bifurcation fractal dimensions,BFD):SB=C×LBD,式中:SB为各级分枝的分枝数,LB为枝长,D为分形维数,C为比例系数(常量)。将上式变换为lnSB=lnC+DlnLB,即在双对数坐标下对SB和LB的一系列值进行线性回归,所得拟合直线的斜率(或其转换),即为分形维数值(BFD)。
2结果与分析
2.1不同生长年限中间锦鸡儿灌丛基本形态指标与地上生物量
由表1可以看出,不同生长年限的中间锦鸡儿灌丛,其冠幅、株高、基径等指标基本随生长年限的增加呈先增大后降低趋势。且各项指标均以16年生中间锦鸡儿为最大。未平茬灌丛的分枝数随林龄的增大而增多,其中更新枝数贡献最大,尤以37年生中间锦鸡儿地面更新枝数最多。表明作为较老林龄的37年生中间锦鸡儿林自我更新能力较强。平茬后的中间锦鸡儿灌丛分枝数较未平茬16年生灌丛高,且平茬后1年生灌丛分枝数高于平茬后生长5年的灌丛。
由图1可知,未经平茬的3种不同生长年限的中间锦鸡儿灌丛的地上生物量随林龄的增加呈现下降趋势,不同茎粗枝生物量的分布,细枝和中枝在总生物量中所占比例排序为16年>25年>37年生灌丛。由灌丛枝构件种群年龄结构分布(图1)可知,当年生与2年生枝在总生物量中所占比例排序为16年>25年>37年生灌丛,与上述不同茎粗枝条生物量分布相一致。且16年生灌丛生物量以当年生和2年生枝为主,而25年和37年生灌丛生物量以2年生以上枝为主。

表1 不同生长年限中间锦鸡儿人工灌丛基本形态指标分析
a)老枝为灌丛中心的主枝;b)更新枝为灌丛外围的地面更新枝。
a) Old branch is the main branch of brush center; b) Renewal branch is the ground renewal branch in the outside of brush.
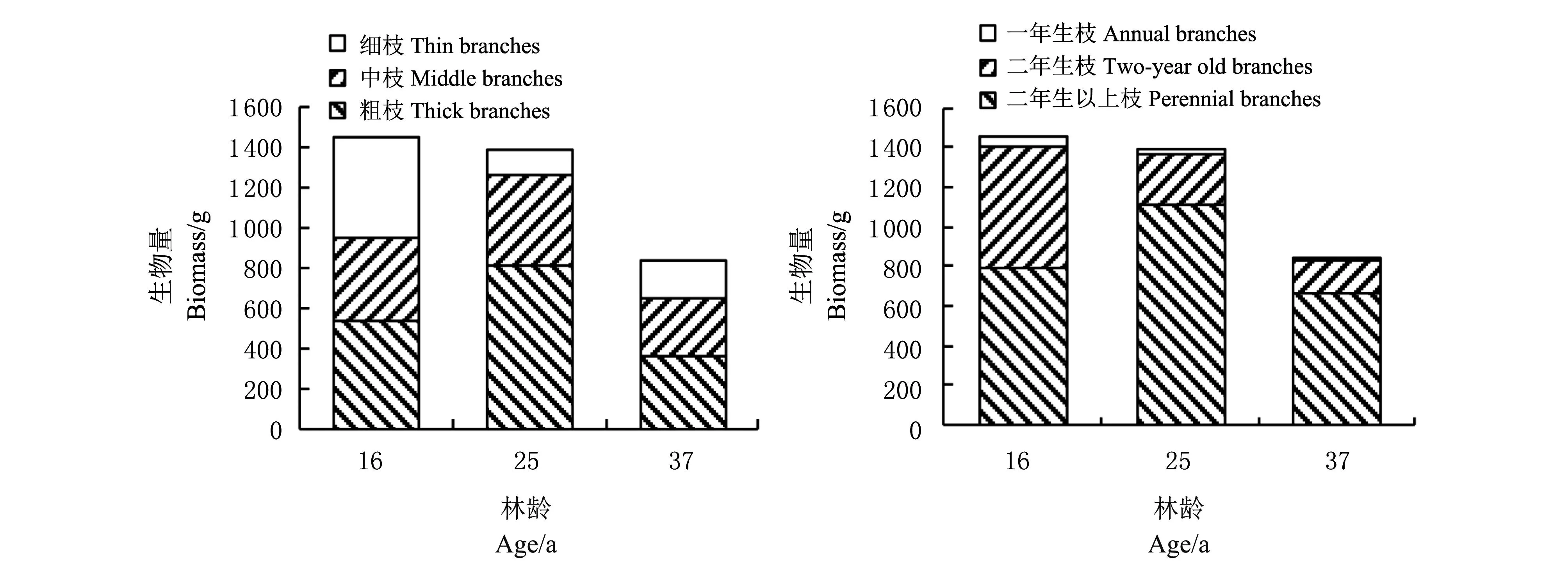
图1 未平茬不同生长年限中间锦鸡儿灌丛枝生物量 Fig.1 Shoot biomass of C. intermedia at different ages without stumping
2.2不同生长年限中间锦鸡儿灌丛不同枝序各级枝的基本指标统计结果
中间锦鸡儿灌丛由外向内枝序分出7个级别。但是并非所有灌丛的枝序都分到第七级,枝序级别数随生长年限的推移先增大后降低,仅有25年生中间锦鸡儿灌丛的枝序分到第七级。不同生长年限的中间锦鸡儿枝生物量随生长年限的增加先增大,在16年林龄达最大,而后随生长年限的增加枝生物量降低(图2)。平茬后1年生与平茬后5年生灌丛各级枝生物量分布无明显规律。16年林龄的灌丛以较高级别的枝生物量相对较小,而低、中级别的枝生物量相对较大。25年与37年林龄的灌丛枝生物量则相反,以低级别的枝生物量相对较小,高级别的枝生物量相对较大。表明老林龄中间锦鸡儿冠层外围的最新分枝发育较差,老林龄灌丛其分枝向外围空间扩展延伸的能力降低。未平茬的16、25和37年生3个林龄中间锦鸡儿各级分枝长度随枝序的增大呈倒“V”型分布(图3)。由图4可知,各生长年限的中间锦鸡儿灌丛其分枝基径随枝序的增大呈严格的单一递增趋势,表明随时间的推移,中间锦鸡儿在生长发育过程中茎不断加粗。

图2 不同生长年限中间锦鸡儿灌丛各级枝生物量统计 Fig.2 Biomass statistics about different order branches of C. intermedia at different ages
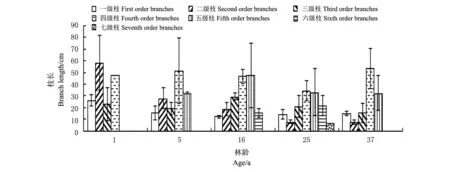
图3 不同生长年限中间锦鸡儿灌丛各级枝枝长统计 Fig.3 Branch length statistics about different order branches of C. intermedia at different ages
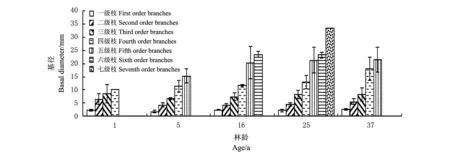
图4 不同生长年限中间锦鸡儿灌丛各级枝基径统计 Fig.4 Basal diameter statistics about different order branches of C. intermedia at different ages
2.3不同生长年限中间锦鸡儿灌丛枝径比
枝径比反映高一级枝条对低一级枝条的支撑能力,对枝条的延伸、分布起着控制性作用,用以衡量植物占据空间的能力[28]。由表2可知,1、5、16、25、37年生灌丛各级枝条平均RBD分别为2.22、1.81、1.65、1.69和1.80,平茬后的枝径比较未平茬灌丛的大,表明未平茬的3种不同林龄的中间锦鸡儿灌丛枝条间承载力均较弱[29]。不同生长年限的中间锦鸡儿各级枝径比基本随枝序的升高而减小,表明枝序级别越高枝条间的承载能力越强,即新枝的承载力相对高于老枝。RBD2∶1为16年<25年<37年,即新枝的承载能力随生长年限的增加而增强;平茬后1年生灌丛的RBD2∶1较平茬后5年生灌丛大,即平茬后5年生灌丛新枝的承载能力相对平茬后1年生灌丛减弱。
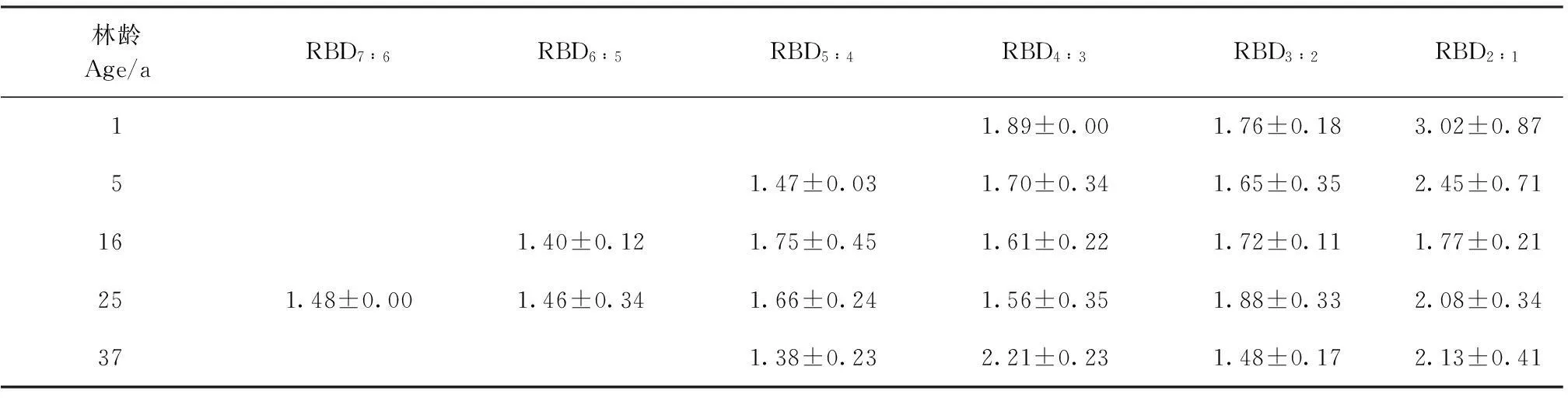
表2 不同生长年限中间锦鸡儿灌丛枝径比统计表
RBD:枝茎比;RBD(i+1)∶i为第i+1级枝与第i级枝的枝茎比。
RBD: Ratio of branch diameter; RBD(i+1)∶iis the ratio of branch diameter between thei+1 order and theiorder branches.
2.4不同生长年限中间锦鸡儿灌丛总体分枝率
分枝率表征枝条产生分枝的能力以及各枝级间的数量配置状况[3]。由表3可以看出,中间锦鸡儿灌丛各枝级枝数之和随生长年限的增加先增大后减小,以16年林龄为最大。16、25、37年生中间锦鸡儿的总体分枝率大小排序为16年>25年>37年,未平茬灌丛的总体分枝率随生长年限的增加而减小。平茬后1年生灌丛总体分枝率较未平茬灌丛大,平茬后5年生灌丛总体分枝率又急剧降低。表明16年生中间锦鸡儿灌丛分枝能力较25、37年林龄强,其占据生态空间的程度和利用生态空间的能力相对较高,平茬有助于刺激灌丛的分枝能力,但平茬后5年生灌丛分枝能力迅速减弱。
表3不同生长年限中间锦鸡儿灌丛总体分枝率统计表
Table 3Overall bifurcation ratio statistics ofC.intermediaat different ages
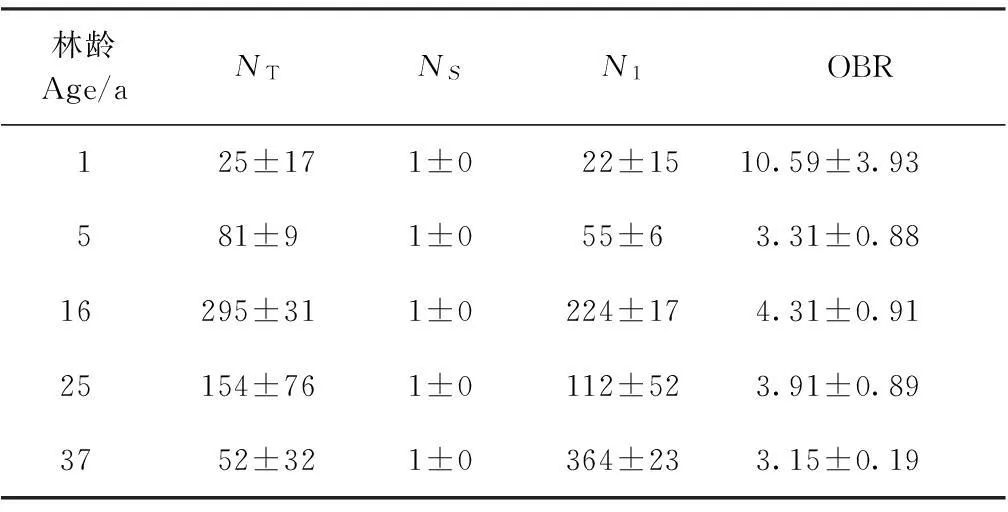
林龄Age/aNTNSN1OBR125±171±022±1510.59±3.93581±91±055±63.31±0.8816295±311±0224±174.31±0.9125154±761±0112±523.91±0.893752±321±0364±233.15±0.19
NT:所有枝序中各级枝条数的总和;NS:最高级枝条数;N1:第一级枝条数;OBR:总体分枝率。
NT: Total branch number of different branch orders;NS: Branch number of the highest order;N1: Branch number of the first order; OBR: Overall bifurcation ratio.
2.5不同生长年限中间锦鸡儿灌丛逐步分枝率
总体分枝率从整体上描述了植物的分枝能力,但未能表现不同枝序各级枝条间的数量比例关系。逐步分枝率表征不同级别枝条的分枝能力[29]。由表4可以看出,不同生长年限中间锦鸡儿灌丛的逐步分枝率随枝序级别的降低,整体呈上升趋势,说明新枝的分枝能力较强,新枝不断向外扩展延伸,对周围空间资源的利用程度较高。在逐级分枝中,以SBR1∶2的生物生态学意义最为重要.尤其是温带落叶植物树种,因其涉及的一、二级枝系是叶片着生的主要部位[3]。未平茬灌丛的SBR1∶2随生长年限的增加而降低,平茬后1年生灌丛的SBR1∶2高于未经平茬的16、25、37年这3种林龄的灌丛,但平茬后5年生灌丛的SBR1∶2又急剧降低,其值介于未平茬25年与37年生中间锦鸡儿灌丛之间,表明老林龄的中间锦鸡儿冠层外围新枝向空间伸展的能力逐渐减弱,平茬虽提高了外围新枝的分枝能力,但平茬5年后冠层外部枝条的分枝能力不及生长25年的未平茬灌丛。

表4 不同生长年限中间锦鸡儿灌丛逐步分枝率统计表
SBR:逐步分枝率;SBRi∶(i+1)为第i+1级枝向第i级枝的分枝率。
SBR: Stepwise bifurcation ratio; SBRi∶(i+1)is the stepwise bifurcation ratio from thei+1 order branch to theiorder branch.
从表4还可发现,不同生长年限的中间锦鸡儿其逐步分枝率由低级到高级最终都归于同一数值2.00。即不论何种生长年限的中间锦鸡儿灌丛其最高级别的逐步分枝率相同,均为2.00。说明中间锦鸡儿灌丛的基部老枝分枝能力始终保持相对稳定的状态。
2.6不同生长年限中间锦鸡儿灌丛枝系分形维数特征
树冠分维数(CFD)将不同枝序的各级枝长与分枝数综合起来,定量描述树木的分枝格局。树冠分维数值一般介于1~2之间,分维数值越大,表明灌丛的空间格局越复杂,其占据空间与利用空间资源的能力越强。未平茬的3种不同林龄的中间锦鸡儿灌丛CFD值均较小,低于1,且随林龄的增大其值降低,而平茬后的灌丛CFD值大于1,尤以平茬后5年生的灌丛为最大。平茬后5年生中间锦鸡儿的CFD值较大,说明其灌丛枝系的空间格局较为复杂,但其分枝率相对较小,则对灌丛空间格局起主要贡献的为枝长,而非分枝数。
表5不同生长年限中间锦鸡儿灌丛分维数比较
Table 5Fractal dimension comparison ofC.intermediaat different ages

林龄Age/aCFDBFD11.25±0.611.48±0.5354.06±2.231.03±0.11160.30±0.031.20±0.20250.26±0.061.15±0.29370.22±0.071.07±0.24
CFD:树冠分维数;BFD:分枝分维数。
CFD: Crown fractal dimensions; BFD: Bifurcation fractal dimensions.
分枝分维数(BFD)是通过对分枝数和枝长在双对数坐标下的一系列值进行线性回归后所得拟合直线的斜率[29]。分枝分维数基本随生长年限的增大而降低,以平茬后1年生灌丛为最大,说明中间锦鸡儿灌丛随着林龄的增长,分枝格局复杂程度降低,平茬后1年生灌丛分枝格局较为复杂,而平茬后5年生灌丛分枝分维数最小,与上述分枝率较小相一致。
3讨论
由于植物的固着生长,植物体在完成整个生活史的过程中,为适应连续变化的外部环境,其生理及形态等方面也不断发生变化。在生长发育过程中,植物体的构件表现出一定的可塑性。构件单元的配置及其动态变化特征反映了植物对空间、光等资源的利用特性及其在特定环境条件下的适应策略[30]。植物的枝系构型越复杂,对空间资源的利用率越高。
不同生长年限的中间锦鸡儿灌丛,从冠幅、株高、基径等基本形态指标来看,16年生中间锦鸡儿均高于其他生长年限的灌丛。且16年生灌丛的地上生物量较25、37年林龄大。但地面更新枝数随着林龄的增大而增多,表明老林龄的中间锦鸡儿灌丛通过萌生更多的更新枝来维持自身的生存,以抵制其衰老的态势。此外,从不同林龄中间锦鸡儿灌丛枝构件种群年龄结构来看,在构件水平上,16、25、37年林龄的中间锦鸡儿灌丛出现衰退型发育的趋势。
枝条长度是对植物枝系向空间伸展能力的衡量,影响植物对资源的利用及不同构件生物量的空间配置情况[29]。由于荒漠草原生境中资源的有限性,为了减少植物种内和种间竞争,荒漠植物需对其周围资源进行充分利用,故其分枝始终呈现不断向外扩展延伸的态势。在本研究中不同生长年限中间锦鸡儿各级分枝长度随枝序的增大基本呈倒“V”型分布,基部较高级枝基本丧失拓展能力,枝长较短;冠层外围新枝枝长也较短,但随着生长发育的推进,新枝不断增长,且一级枝不断发展成二级枝,又有新的一级枝出现。如此循环重复,使灌丛不断扩充,空间格局复杂化。且在各级分枝发育变化的同时,生物量的分配表现出与枝长类似的规律。
分枝格局决定了植物对资源的利用情况,同时也是对外界环境的一种响应[30]。枝径比表征枝条间的承载力。在本研究中,较老枝的枝条间承载力较低,分枝率也较低;新枝的枝条间承载力较大,分枝率也较高。这与何明珠[29]的研究结论,即枝径比高的其分枝率也较高相一致。中间锦鸡儿灌丛分枝率随生长年限的增大而降低,表明16年生中间锦鸡儿灌丛分枝能力较强,随林龄逐渐变大,灌丛分枝能力减弱,对空间资源的利用率减小;平茬有利于激发灌丛的分枝潜能,但平茬后5年生的灌丛分枝能力又急剧减弱,尤以冠层外围新枝最为明显。16年生中间锦鸡儿新枝的逐步分枝率在未平茬的3种林龄中最高,冠层外围新枝的分枝能力较强,对空间资源的利用程度较高。此外,中间锦鸡儿基部老枝的分枝能力最弱,但较为稳定,基本不随生长年限发生变化。
分形维数揭示树冠分枝格局占据生态空间的程度和利用生态空间的能力,其生态学意义在于刻画分枝格局对环境资源的利用能力[31]。树木冠型结构(如枝条数量、分枝特性等)对林木的生长和生产力具有极为重要的影响,是树木生长的主要决定因子之一[4]。未平茬的3种林龄的灌丛树冠分维数和分枝分维数随林龄的增加而减小,表明生长年限较短的16年生中间锦鸡儿灌丛的枝系空间格局最为复杂,其占据空间的能力较强,对空间资源的利用潜能相对较高。而37年生中间锦鸡儿灌丛枝系空间格局最为简单。平茬后的灌丛2种分维数均大于未平茬灌丛,平茬增加了灌丛分枝格局的复杂程度,平茬5年生灌丛主要以各级枝长增大了空间格局的复杂程度,分枝数对其影响不大。
以上所述,16年生中间锦鸡儿灌丛枝系构型较为复杂,但是随林龄的增长,灌丛空间结构趋于简化,平茬有利于丰富灌丛的枝系构型,增强其在空间的填充程度。
参考文献(References):
[1]Harper J L. Plant demography and ecological theory.Oikos, 1980,35:244-253.
[2]Barlow P W. Meristems, metamers and modules and the development of shoot and root systems.BotanicalJournaloftheLinneanSociety, 1989,100(3):255-279.
[3]陈波,宋永昌,达良俊.木本植物的构型及其在植物生态学研究的进展.生态学杂志,2002,21(3):52-56.
Chen B, Song Y C, Da L J. Woody plant architecture and its research in plant ecology.ChineseJournalofEcology, 2002,21(3):52-56. (in Chinese with English abstract)
[4]李火根,黄敏仁.分形及其植物研究中的应用.植物学通报,2001,18(6):684-690.
Li H G, Huang M R. Fractal theory and its application to plants researches。ChineseBulletinofBotany, 2001,18(6):684-690. (in Chinese with English abstract)
[5]何明珠,王辉,张景光.民勤荒漠植物枝系构型的分类研究.西北植物学报,2005,25(9):1827-1832.
He M Z, Wang H, Zhang J G. Classification of the branching architectures of the desert plants in Minqin County.ActaBotanicaBoreali-OccidentaliaSinica, 2005,25(9):1827-1832. (in Chinese with English abstract)
[6]Borchert R, Slade N A. Bifurcation ratios and the adaptive geometry of trees.BotanicalGazette, 1981,142(3):394-40.
[7]Halle F, Oldeman R A A, Tomlinson P B.TropicalTreesandForests:AnArchitecturalAnalysis. New York: Springer-Verlag, 1978:233-236.
[8]Kuppers M. Ecological significance of aboveground architectural patterns in woody plants: A question of cost-benefit relationships.TrendsinEcology&Evolution, 1989,4(12):375-379.
[9]马克明,祖元刚.兴安落叶松分枝格局的分形特征.植物研究,2000,20(2):235-241.
Ma K M, Zu Y G, Fractal characteristics of the branching pattern ofLarixgmelini.BulletinofBotanicalResearch, 2000,20(2):235-241. (in Chinese with English abstract)
[10]冯长根,李后强,祖元刚.非线性科学的理论、方法和应用.北京:科学出版社,1997:115-120
Feng C G, Li H Q, Zu Y G.TheTheory,MethodandApplicationofNonlinearScience. Beijing: Science Press, 1997:115-120. (in Chinese)
[11]田超,刘阳,杨新兵,等.冀北山地阴坡优势树种的树体分维结构.生态学报,2011,31(22):6753-6765.
Tian C, Liu Y, Yang X B,etal. Fractal structure of dominant tree species in north-facing slope of mountain of northern Hebei.ActaEcologicaSinica, 2011,31(22):6753-6765. (in Chinese with English abstract)
[12]邹修国.分形理论的发展及其在农林业上的应用.江西农业学报,2011,23(4):118-120.
Zou X G. Development of fractal theory and its application in agriculture and forestry.ActaAgriculturaeJiangxi, 2011,23(4):118-120. (in Chinese with English abstract)
[13]张莉.基于分形理论的计算机辅助园林设计的研究与应用.济南:山东师范大学,2007.
Zhang L. Computer-aided landscape architecture design based on fractal theory and application, Jinan: Shandong Normal University, 2007. (in Chinese with English abstract)
[14]李学玲,林慧龙.分形理论在草地科学中的应用概述.草地学报,2011,19(4):705-711.
Li X L, Lin H L. Fractal theory application on grassland science (review).ActaAgrestiaSinica, 2011,19(4):705-711. (in Chinese with English abstract)
[15]李火根,阮锡根,王友箐,等.林木复杂性状分维数计算软件(FDC 1.0)的研制与应用.南京林业大学学报:自然科学版,2004,28(3):5-8.
Li H G, Ruan X G, Wang Y Q,etal. Development and application of software for fractal dimension calculation (FDC 1.0) of complex traits in forest trees.JournalofNanjingForestryUniversity:NaturalSciencesEdition, 2004,28(3):5-8. (in Chinese with English abstract)
[16]常杰,陈刚,葛滢.植物形态结构定量研究的新方法:分形模拟.植物学通报,1996,13(2):57-62.
Chang J, Chen G, Ge Y. A new method of quantitative study on plant structure and morphology: Fractal simulation.ChineseBulletinofBotany, 1996,13(2):57-62. (in Chinese with English abstract)
[17]周海燕,张景光,李新荣.生态脆弱带不同区域近缘优势灌木的生理生态学特性.生态学报,2005,25(1):168-175.
Zhou H Y, Zhang J G, Li X R. Ecophysiological characteristics of kindred dominant shrubs in different areas of ecologically vulnerable zone.ActaEcologicaSinica, 2005,25(1):168-175. (in Chinese with English abstract)
[18]赵艳云,程积民,王延平,等.半干旱区环境因子对柠条灌木林结构的影响.水土保持通报,2005,25(3):10-14.
Zhao Y Y, Cheng J M, Wang Y P,etal. Influence of environmental factors on formation of morphological structure ofCaraganakorshinskiiin semi-arid region.BulletinofSoilandWaterConservation, 2005,25(3):10-14. (in Chinese with English abstract)
[19]张帆,陈建文,王孟本.幼龄柠条细根的空间分布和季节动态.生态学报,2012,32(17):5484-5493.
Zhang F, Chen J W, Wang M B。 The spatial distribution and seasonal dynamics of fine roots in a youngCaraganakorshinskiiplantation.ActaEcologicaSinica, 2012,32(17):5484-5493. (in Chinese with English abstract)
[20]宋乃平,王磊,张庆霞,等.农牧交错区典型村域的土地利用变化过程研究:以盐池县马儿庄村为例.资源科学,2010,32(6):1148-1153.
Song N P, Wang L, Zhang Q X,etal. A study on dynamic changes of land use in typical villages in farming-grazing transitional zones: A case study on the Ma’erzhuang Village Yanchi County.ResourcesScience, 2010,32(6):1148-1153. (in Chinese with English abstract)
[21]闫志坚,扬持,高天明,等.6种常用固沙植物的生态经济价值比较.干旱区资源与环境,2006,20(3):163-168.
Yan Z J, Yang C, Gao T M,etal. The comparison study of eco-economy value on six fixing sandy plants.JournalofAridLandResourcesandEnvironment, 2006,20(3):163-168. (in Chinese with English abstract)
[22]Oohata S, Shidei T. Studies on the branching structure of trees Ⅰ. Bifurcation ratio of trees in Horton’s law.JapaneseJournalofEcology, 1971,21:7-14.
[23]何明珠,张景光,王辉.荒漠植物枝系构型影响因素分析.中国沙漠,2004,26(7):626-627.
He M Z, Zhang J G, Wang H. Analysis of branching architecture factors of desert plants.JournalofDesertResearch, 2004,26(7):626-627. (in Chinese with English abstract)
[24]林勇明,洪滔,吴承祯,等.桂花植冠的枝系构型分析.热带亚热带植物学报,2007,15(4):301-306.
Lin Y M, Hong T, Wu C Z,etal. Architectural analysis ofOsmanthusfragranscrown geometry.JournalofTropicalandSubtropicalBotany, 2007,15(4):301-306. (in Chinese with English abstract)
[25]孙书存,陈灵芝.辽东栎植冠的构型分析.植物生态学报,1999,23(5):433-440.
Sun S C, Chen L Z. Architectural analysis of crown geometry inQuercusliaotungensis.ActaPhytoecologicaSinica, 1999,23(5):433-440. (in Chinese with English abstract)
Li J, Zhu J Z, Zhu Q K. Review on methods of calculating fractal dimension.JournalofBeijingForestryUniversity, 2002,24(2):71-78. (in Chinese with English abstract)
[27]Mandelbrot B B.TheFractalGeometryofNature. San Francisco: W. H. Freeman and Company, 1982:286-287.
[28]林勇明,俞伟,刘奕,等.不同距海处木麻黄分枝格局特征及冠形分析.应用与环境生物学报,2013,19(4):587-592.
Lin Y M, Yu W, Liu Y,etal. Characteristics of branching pattern and crown morphology ofCasuarinaequisetifoliaL. in different coastal distances.ChineseJournalofApplied&EnvironmentalBiology, 2013,19(4):587-592. (in Chinese with English abstract)
[29]何明珠.荒漠植物枝系构件及其持水力研究.兰州:甘肃农业大学,2004.
He M Z. Branching module and water-retaining capability of desert plants. Lanzhou: Gansu Agricultural University, 2004. (in Chinese with English abstract)
[30]张广帅,邓浩俊,俞伟,等.不同密度下木麻黄幼苗枝系特征变化的研究.热带作物学报,2014,35(6):1062-1066.
Zhang G S, Deng H J, Yu W,etal. Architectural responses ofCasuarinaequisetifoliaL seedlings to different population density.ChineseJournalofTropicalCrops, 2014,35(6):1062-1066. (in Chinese with English abstract)
[31]周元满,王平,刘素青,等.无瓣海桑人工林树冠结构的分形分析.福建林学院学报,2012,2(3):252-256.
Zhou Y M, Wang P, Liu S Q,etal. Fractal analyses on crown structure ofSonneratiaapetalaplantation.JournalofFujianCollegeofForestry, 2012,2(3):252-256. (in Chinese with English abstract)
*通信作者(Corresponding author):吕晓男,Tel:+86-571-86404270;E-mail:luxngis@163.com
第一作者联系方式:邓勋飞,E-mail:dengxf@mail.zaas.ac.cn
URL:http://www.cnki.net/kcms/detail/33.1247.s.20150519.1346.014.html
猜你喜欢
中国交通信息化(2022年7期)2022-10-27 06:36:28
今日农业(2020年19期)2020-12-14 14:16:52
农家科技中旬版(2020年7期)2020-07-24 08:39:04
安徽农学通报(2020年7期)2020-05-26 01:58:28
甘肃科技(2020年21期)2020-04-13 00:33:46
中国野生植物资源(2019年2期)2019-06-11 08:01:44
吉林农业(2017年18期)2017-09-15 08:16:54
湖南林业科技(2017年1期)2017-02-06 05:29:12
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:47
